Embed Size (px)
Citation preview

Institut de zoologic, Facultd des sciences, Universitd de Kioto, Japo~
Reeherches sur l'expression des faeteurs ldtaux hdrdditaires ehez l'embryon de la drosophile
I I I . La v a r i a t i o n d e l a c h a r g e 6 1 e c t r i q u e ~ l a s u r f a c e de l ' oeuf p e n d a n t le d f v e l o p p e m e n t i n i t i a l
Par
Tadaslli Imaizumi
Avec 3 figures
Introduction
Des observations sur la variation de la charge dlectrique ~ la surface ont faites l'objet de cette dtude. Les mafdriaux employds sont les ceufs des mouches sauvages de Kitakata-24 chez D. oirilis, d'Oregon-R chez D. melano-
~jvsteur rtdrtsst'ur dr" voltage /
/ 81npereln~tp~
T TY\ ~ I i o I \ I ~lectPod#
#'altm ~ tre
Fig'. i. L'appareillage des experiments.
gaster et le mutant g X-attach6 chez D~ melanogaster. Tousles ~eufs out ~t6 d6chorionn~s prSalablement et un champ ~lecirique form~ dans de la paraffine liquide a ~t6 employ& Voici un sch6ma de cet appareillage (Fig. i). En ce qui concerne les stades du d~veloppement normal, le diagramme standard donnd dans notre premigre 6rude a df~ adopt6; pour les embryons l~taux et les ~eufs vierges les stades suivants out 6td adopt&:
(Eats vierges : u C'est le sfade oh le protoplasme s'est contract6 et a un ellipsoYde.
II correspond au stade 2 de l'embryon f~cond& VD: C'est le moment o~le protoplasme s'est cytolys& I1 correspondait
un stade entre 5 et 9 chez l'embryon f~condd, si l 'muf s'&ait d~velopp&

T. Imaizumi: L'expression des facteurs 16taux h6r6ditaires, etc. 23
(Eufs 16taux aux d6pourvus de chromosome X (nullo X): L C : C'est un mdlange des premiers stades des embryons ldtaux et
normaux, lorsque le protoplasme s'est contract6; l ' embryon ldtal de nullo X diminue en effet de volume, lui aussi, et il ne peut 6t6 distingu6 des normaux h ce stade.
L B D : C'est le stade off le protoplasme a atteint son volume maximum. I1 correspond au stade 5 de l '(euf fdcond6.
Les observations ont 6t6 exdcut4es sur des embryons plac4,s dans de la paraff ine liquide. La dur4e pendant ]aquelle l ' embryon peut survivre a 4t4
observ4e; ees observations sont nof4es dans les tableaux suivants.
Tableau l. D4veloppement d'embryons qui ont 4t4 plac4s pendant dix minutes dans de la paraffine liquide au stade 2, puts report4s dans da Ringer.
incub6s . . . . . . . . . . . . . . . . . 11 embryons anormaux, non incubds . . . . 8 organogen~se ineomplbte . . . . . . . . 7 aucun d4veloppement . . . . . . . . . . 0
Total des embryons . . . . . . . . . . . 26
Tableau 2. D6veloppement d'exnbryons qui ont dt6 plac6s pendant trente minutes dans de la paraffine liquide au stade 5, pats report,s dans du Ringer.
incub4s . . . . . . . . . . . . . . . . . 20 organogen6se complete, non incub6s . . . 10 embryons anorlnaux, non incub4s . . . . l0 organogen~se incomplete . . . . . . . . t0 aucun d6veloppement . . . . . . . . . . 0
Total des embryons . . . . . . . . . . . 50
Ces montrent que l ' embryon peut survivre normalement dans de la paraff ine liquide pendant un moment; par cons4quent routes des observa- tions ont 4t4 limit4es h trois ou quatre minutes.
O b s e r v a t i o n s
Les observations ont 4t4 ex4cut4es sur des embryons plac6s dans un petit seau de verre rempli de paraffine liquide pure. Quand le courant 41ectrique direct a 4t4 mis entre deux 41ectrodes de cuivre d ' u n e surface de 5 mm z, le comporte- ment des o~ufs a 4t4 observ4 avee un microscope horizontal. Quat re observations distinctes ont 4t4 faites.
Observat ion I. Les 41ectrodes sont 6t4 plac4s ~t droite et h gauche h une distance de 5ram (Fig, 2). Quand le mat4rial 4tudi4, qui avait 4t4 laiss4 tom- ber de la surface de paraff ine liquide, 4fair arriv4e
Fig 2. L'appareil par l'observation I.
au milieu des 41ectrodes, un courant a 4t4 lanc4 ef la r4action de l'ceuf a 4t6 observ4e. Les ceufs vivants, les ceufs tu4s par de l 'eau bouillante et le protoplasme extrai t de l'ceuf en pressant la membrane vitelline dans la

24 T. Imaizumi
paraff ine liquide se meuvenf tous vers la cathode. On salt que, chez rceuf de la drosophile, on peur distinguer la part ie cdphalique de celle qui se trouve en art iste, parce qu'i] y a une petite pro~minence du c6t~ de la tSte; quand l'o3uf a 6t$ soumis g une diff6rence de potentiel, l 'axe longi- tudinal est parall~le h la force 61ectrique; dans ce cas, ta direction vers laquelle la pointe de la tSte de l'o3uf est orient~e n 'a aucune relation avec la port ion des ~lectrodes.
l~tant donn6 qu 'une couche double 61ectrique se forme h la surface par taquelle deux di~lectriques diff6rents sont en contact, une charge n~- gative est donn6e ~ la paraff ine tiquide et une charge positive ~ la surface de l'ceuf.
Observat ion If. Quand un voltage est lanc6 ~t quelques ceufs qui avaient 6t6 mis en une lois dans le seau rempli de paraffine liquide,
i~ _~ ces ceufs se p lacent en une rang~e formant un pont entre les deux 6lectrodes; ~ ce moment, le protoplasme de chaque o~uf est inclin6 vers te cSte de t 'anode. Quoique le protoplasme
~ puisse ~tre tu~ par le courant ~lectrique, il semble bien quql
J y air, dans le protoplasme, beaucoup de particules charg~es n6gativement.
Une observation d'~lectrophor~se a dtd alors ex6cutde : des ~lectrodes de platine et te protoplasme d 'un o3uf broy~ dans de t 'eau distill~e aux stades 2 et 5, ont ~t~ emp loySs :dans ce routes les particules se meuven~ vers F a n o d e : i l en c a s ,
- - r~sulte quc routes les particules par lesquelles ]e protoplasme Fig. 3. L~ap - est constiLu~ sont chargdes n~gativemeni ~t l 'eau. parei[ par Observat ion I IL Les ~lectrodes ont 4t~ dispos~es en haut
l'obser- et bas (Fig. 3) et ]a valeur du voltage pour laquelle un ceuf ration III. qui a ~td mis sur l'61ectrode inf~rieure commence h se mou-
voir a ~t~ mesur~e. La distance entre les deux ~lectrodes est de I mm eL ta valenr du voltage a ~t~ mesur~e pour chaque (euf. Les r~sultats sont indiqu~s dans les tableaux 3 eL 4.
Tableau 3. Yaleurs du voltage pour lesquelles un eeuf ~ chaque stade dlez D. Dirilis commence ~t se mouvoir. Le signe entre parenthbses indique le signe de
la charge dans l'61ectrode sup6rieure. La temp6rature est de 18~176
Yaleur moyenne (volt) Limite Total Stade et erreur probable des valeurs des ceufs
2 (+) 174 - 0,7 150 N 200 ~5i '3 (+) t87 -4- 0,7 t60 ~, 200 71 5 (+) t97 + 0,5 170 ,~ 220 152 6 (+) 185 + 0,7 t60 ~ 200 84 9 (+) t80 + 0,7 t60 ,~ 210 89
2 (--) ~z~ + 05 16o ~ 18o 1o5 3 (--) 172 +0,4 160,-, t80 t13 5 (--) 176 + 0,3 160 ,-, 190 177 6 (--) 174 + 0,4 160 ~ 190 t51 9 (--) 174 -_4_-_ 0,4 160 ,-~ 190 112
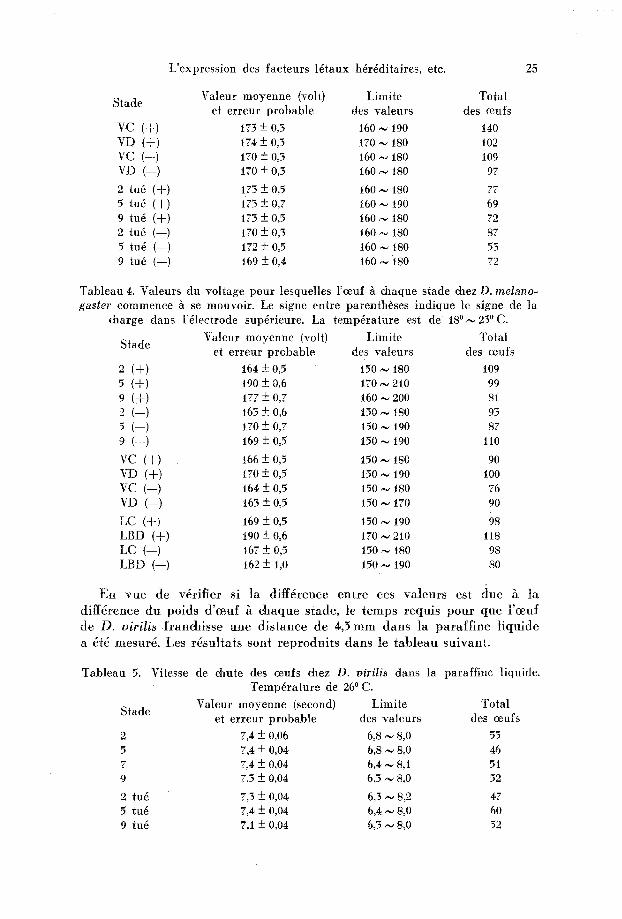
L'express ion des fac teurs 16taux h6r6ditaires, etc. 25
Valeur moyenne (vol t ) Limi te Tota l Stade
et e r reur p robab le des valeurs des ceufs
VC (+) 173 + 0,3 160 ,,~ 190 140 VD (+) 174 + 0,3 t70 ~ 180 t02 VC (--) 170 + o,3 16o ~ 180 1o9 VD (--) t7o + o,5 16o ,-~ 18o 97
2 tu6 (+) 173 --_ 0,5 t6o N t8o 77 5 tu6 (+) 173 + o,7 16o ~ t9o 69 9 tu6 (+) 173 +-- 0,5 t60 ~ 180 72 2 tu6 (--) 170 + 0,3 160 ~ iS0 87 5 fu@ (--) 172 --- 0,5 t60 ~ iS0 53 9 tu6 (--) t69 + 0,4 t60 ~,, 180 72
Tab leau 4. Valeurs du vol tage pour lesquelles l 'ceuf ~ chaque stade chez D. melano- gaster commence ~ se mouvoir . Le signe entre parentheses ind ique le signe de la
daarge dans l '61ectrode sup6rieure. La temp6ra ture est de 1S~ 23~
Valeur moyenne (volt) L imi te Tota l Stade
et e r reur p robab le des valeurs des oeufs
2 (+) t64 -+ 0,5 150 ,,~ 180 109 5 (+) 190 + 0,6 170 ,~ 210 99 9 (+) 177 + 0,7 t60 ,~ 200 S1 2 (--) t63 + 0,6 150 ~., 180 93 5 (--) t70 + 0,7 t50 ,~ 190 87 9 (--) 169 + 0,5 150 ~ 190 110
VC (+) t66 + 0,5 150 ,~ 180 90 u (+) 170 + 0,5 150 ,~ 190 100 VC (--) 164 + 0,5 150 ~ 180 76 VD (--) i63 + 0,5 150 ,~ 170 90
LC (+) 169 + 0,5 t50,-~ 190 98 LBD (+) t90 + 0,6 170 ,~ 210 118 LC (--) 167 + 0,5 150 ~ t80 98 LBD (--) 162 + 1,0 150,~ 190 so
E n r u e d e v d r i f i e r si ta d i f f d r e n c e e n t r e ces v a l e u r s es t d u e h l a
d i f f d r e n c e d u p o i d s d ' o~uf ~t c h a q u e s t ade , le t e m p s r e q u i s p o u r q u e l ' c eu f
d e D. o i r i l i s f r a n c h i s s e u n e d i s t a n c e d e 4 , S m m d a n s l a p a r a f f i n e t i q u i d e
a dr6 m e s u r d . Les r ~ s u l t a t s son t r e p r o d u i t s darts le t a b l e a u s u i v a n t .
Tab leau 5. Vitesse de daute des ceufs chez D. oirilis darts la paraff ine l iquide. Temp6ra tu re de 260 C.
Valeur moyenne (second) Limi te Tota l Stade
et e r reur p robab le des valeurs des oeufs
2 7,4 + 0,06 6,8 ,-~ 8,0 55 5 7,4 + 0,04 6,8 ,~ 8,0 46 7 7,4 + 0,04 6,4 ~ 8,1 51 9 7,3 - 0,04 6,3 N 8,0 52
2 tu6 7,3 +-- 0,04 6,3 ~ 8,2 47 5 tu~ 7,4 --- 0,04 6,4 ,~ 8,0 60 9 tu~ 7,1 --+ 0,04 6,3 ,~ 8,0 52
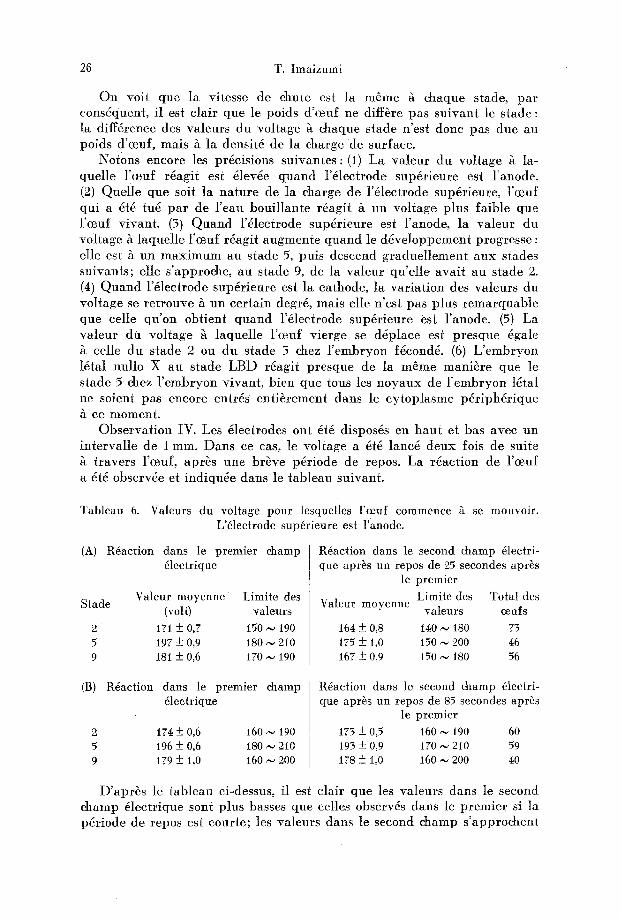
26 T. [maizumi
On voit que la vitesse de chute est la m6me h chaque stade, pa r consdquent, il est clair que le poids d 'ceuf ne diff~re pas su ivant le s tade : la diff6rence des valeurs du vol tage h chaque stade n 'est donc pas due au poids d'ceuf, raais ~t la den sit4 de la charge d e surface.
Not0ns encore les prdcisions su ivantes : (1) La va leur du vol tage h la- quelle l 'ceuf r6agit est 4lev6e quand l '6lectrode supdrieure est l 'anode. (2) Quel le que soit la na tu re de la charge de l '61ectrode sup~rieure, l 'ceuf qui a dtd tug pa r de Feau boui l lante r6agit & un vol tage plus faible que l'ceuf vivant . (3) Q u a n d l '6lectrode supdrieure est l 'anode, la va leur du voltage & laquel le l 'ceuf r~agit augmente quand le d6ve loppement progresse : elle est h un m a x i m u m au s tade 5, puts descend graduel lement aux stades suivants; elle s 'approche, au s tade 9, de la va leur qu 'el le ava i t au s tade 2. (4) Q u a n d l'61ectrode sup~rieure est la cathode, la var ia t ion des valeurs du vol tage se re t rouve & un cer ta in degr6, raais elle n 'est pas plus r emarquab l e que celle qu 'on obtient q u a n d l '6lectrode sup6rieure est l 'anode. (5) La va leur du vol tage ~t laquelle l'~euf vierge se d6place est presque @ale
celle d u s tade 2 ou du stade 5 chez l ' e rabryon f6cond& (6) L ' e m b r y o n 16tal hullo X au s tade LBD r6agit p resque de la m~me raani~re que le s tade 5 chez l ' e rabryon vivant , bien que t ou s l e s noyaux de l ' embryon ldtal ne soient pas encore entr6s ent i~rement dans le cy top lasme pdr iph6r ique ~t ce moment .
Observa t ion IV. Les 61ectrodes ont did dispos6s en hau t et bas avec un interval le de 1 rata. Dans ce cas, le vol tage a 6t4 lanc4 deux fois de suite
t ravers lkeuf, apr~s une br~ve p~riode de repos. La r6action de l 'ceuf a 6td observde et indiqu~e dans le t ab leau suivant .
Tableau 6. Valeurs du voltage pour lesquelles l'ceuf commence ~t se mouvoir. L'6lectrode sup6rieure est l'anode.
(A) R6action dans le premier champ 61ectrique
Valeur moyenne Limite des Stade
(volt) valeurs 2 171 + 0,7 150 ,-, 190 5 197 + 0,9 180 ,-~ 210 9 181 - 0,6 170 ,-~ 190
Rdaction dans le second champ 61ectri- que apr~s un repos de 25 seeondes apr~s
le premier Limite des Total des
Valeur moyenne valeurs ceufs
164 _+ 0,8 140 ,-., 180 73 175 _+ 1,0 150 ~ 200 46 167 + 0,9 150 ,-, IS0 56
(B) R6aetion dans le premier champ 61ectrique
2 174 + 0,6 160 ~ 190 5 196 + 0,6 180 ,-, 210 9 179 -4-__ 1,0 160 ,-, 200
R~action dans le second champ 61ectri- que apr6s un repos de 85 seeondes apr~s
le premier 173 + 0,5 160 ,-~ 190 60 193 + 0,9 170 N 210 59 178 + 1,0 160 ,~ 200 40
D 'apr~s le tab leau ci-dessus, il est clair que les valeurs dans le second champ ~lectrique sont plus basses que celles- observds dans le p remie r si la p~riode de repos est courte; les valeurs dans le second champ s ' approchent

L'expression des facteurs 16taux h6r6ditaires, etc. 27
des premi6res si le temps de repos est allong6. Ce ph6nombne signifie que la charge h la surface qui a 6td obtenue dans le champ dlectrique ne disparait pas tout de suite apr~s que le courant 6lectrique a dt4 arr6t6.
Discuss ion
Des observations sur la charge & la surface des (cuts existent dans le cas des oursins et des amphibiens, mats ce phgnom~ne n'avait pas encore 6t6 6tudi6 chez l'ceuf d'insecte.
Nous avons d$crit dans le travail pr6c6dent les dldments du proto- plasme de l'ceuf de drosophile et leur mouvement : rappelons que, au stade off le protoplasme s'est contract6, les noyaux se divisent g6ndralemeut dans la partie int6rieure de l'ceuf, ils sont envelopp~s d'un cytoplasme fiche en petits granules; les granules vitellins sont disperses ef isol6s du cyto- plasme. Lorsque le d6veloppement progresse, les noyaux, qui sont envelopp6s dans l'ilot protoplasmique, se meuvent vers la p~riph6rie; au coutraire, les granules vitellins se r6unissent/t l'int~rieur de l'(euf. Au stade de la formation du blastoderme, les noyaux, les petits granules et les granules vitellins sont stratifies r@uli~rement: en une seule couche de cel- lules, y compris les noyaux, ~t la surface, une couche constitute par de nombreux petits granules au milieu, et un groupe de granules vitellins ~t l'int6rieur.
Dans les cellules qui se sont tangles h la p~riph6rie, la masse de chromatine, le suc nucl~aire des noyaux et le cytoplasme des cellules sont aussi arraIrg~s r@uli~rement : le noyau occupe la place la plus ext6rieure de la cellule et la masse de la chromatine se localise dans la partie la plus ext~rieure du noyau. I1 existe donc un arrangement tr~s r@ulier des ~16ments du protoplasme & ce stade du blastoderme chez l'embryon normal; c'est pr6cis~ment /t stade que la valeur de voltage qui agit sur l'ceuf est la plus 61ev~e.
Or, il est connu que les amino-acides et les prot~ines ont les carac- t~ristiques physiques et chimiques suivantes:les amino-acides sont repr6- sent~s comme des cations par l a formule de HaN + . R . COOH en solution acide et comme des anions par la formule de H2N. R. C O O - en solution alcaline. Les prot6ines montrent essentiellement le m6me comportement que les amino-acides, mats elles en different principalement par la multi- plicit6 de leurs groupes anion e t cation. Donc, les prot~ines, tout comme
les amino-acides, sont charges ~lectriquement. On sait aussi (1) que dans une substance charg6e 61ectriquement, la
condition 61ectrique int~rieure est repr~sent~e comme une charge de la surface, si elle est conductrice. (2) Si une molecule polaire est plac~e dans un champ ~lectrique, elle peut ~tre soumise & la polarisation di61ec- trique et, par consequent, elle peut ~tre orient~e dans une direction d~finie. Des molecules non polaires typiques peuvent aussi devenir des dipoles quand elles sont plac~es dans un champ 61ectrique.
I1 semble que les r~gles physiques et chimiques ci-dessus sont aussi applicables h l'6tre vivant. D'ailleurs, il a 6t~ en observ~ par beaucoup

28 T. tmaizumi
d'investigateurs de la physiologie cellulaire que le protoplasme de la cellule vivante est en g6n6ral charg6 ~lectriquement. Puisque le protoplasme est constitu6 en grande partie par les protdines et tes nucldoprotdines, la cellule est ndcessairement chargde dlectriquement elle-m~me; il dolt en r6sulter une charge /~ la surface de la cellule. I1 est donc probable que les variations de la charge "~ ta surface au cours du d4veloppement r~sultent des mouvements actifs causds par la synth~se, la d~composition et la transformation des substances internes de l'(euf qui sont charg6s 61ectrique- ment. Ces variations ne sont toutefois pas la cause, mais le r~sultat de ces mouvements actifs.
I1 est impossible de tirer des conclusions d6finitives des donn6es signal~es ci-dessus; on pout cependant discuter tes points suivants. I1 semble que l'~16vation de la valeur du voltage /~ taquelle l'(euf rdagit repr6sente ta diminution de la charge/~ ta surface de l'r car si la deusit6 de la charge /~ la surface est faible, t'~euf ne peut r6agir qu'~t un voltage 6lev& Cela signifirait que la condition d'association des moldcules des matdriaux constituant le protoplasme int6rieur est maintenue de far plus stable. Le fair que la valeur du voltage est maximal au stade de la formation du blastoderme chez l 'embryon normal, au moment oth la con- dition de la distribution des dl6ments du protoplasme esf r@l~e, plaide dans ce sens. En o utre, la comparaison de l 'embryon 16tal au stade LBD avec l 'embryon normal ~t stade 5, ne montre gu~re de differences entre les deux. I1 taut en conclure que les mouvements mdtaboliques du protoplasme chez l 'embryon ldtal seraient exdcut~s jusqu'/~ ce stade, bien que les noyaux ne se d~placent pas r@uli~rement vers la p6riph~rie.
R 6 s u m 6
Les variations de la charge ~ la surface ont ~t6 observdes dans un champ $lectrique constant form~ dans de la paraffine liquide pure. L'ceuf est charg6 n@ativement par rapport ~t ta paraffine liquide. La valeur du voltage/t laquelle l'o~uf r~agit est 6levde au stade de la premiere contraction de volmne, lots de la formation du blastoderme, atteint un maximum ~t ce stade et baisse ensuite tors de la formation de la go utti~re ventrale. Au stade de la formation de la bande germinale, elle s'approche de la valeur initiale. La valeur du voltage ~t laquelle l'o~uf vierge ou tu~ par l'eau bouillante r6agissent est presque ~gale/~ celle du stade initial de l 'embryon fdcondg. Au stade ofa le volume du protoplasme atteint son maximum, la r~action chez l 'embryon l~tal de D. melanogaster est presque @ale h celle trouvde au stade de la formation du blastoderme chez l 'embryon normal.
Travaux eit6s
H a u r o w i t z, F., 1950: Chemistry and biology of proteins.
I m a i z u m i , T., et K. N a k a m u r a , t951: La charge ~lectrique h la surface chez l'ceuf de Drosophila pendant le d6veloppement initial (on japonais). Zool. Mag. 60, 53--54.

L'expression des facteurs 16taux h6r6ditaires, etc. 29
N a k a m u r a , K., and T. I m a i z u m i , 1949: Electric charge at the surface of developing embryos in D. oirilis. Drosophila Informat ion Service 23, 94.
- - - 1950: Sur la charge 61ectrique ~t la couche de surface chez l 'embryon de Drosophila pendant le d6veloppement initial (en japonais). Zool. Mag. 59, iS.
S o n n e n b 1 i ck , B. P., 1947: Synchronous mitoses in Drosophila, their intensely rapid rate, and sudden appearance of the nucleolus. Rec. Gen. Soc. Amer; 16, 52.
- - t950: The ear ly embryology of Drosophila melanogaster. Biology of Droso- phila 62--167.