Embed Size (px)
Citation preview

Acquisition of species-specific O-linked carbohydrate chains from oviducalmucins in Rana arvalisA case study
Alexandra Coppin, Emmanuel Maes, Christophe Flahaut, Bernadette Coddeville and Ge rard Strecker
Laboratoire de Chimie Biologique, Unite Mixte de Recherche du Centre National de la Recherche Scientifique n88576,
Universite des Sciences et Technologies de Lille (Flandres-Artois), Villeneuve d'Ascq, France
The extracellular matrix surrounding amphibian eggs is composed of mucin-type glycoproteins, highly
O-glycosylated and plays an important role in the fertilization process. Oligosaccharide-alditols were released
from the oviducal mucins of the anuran Rana arvalis by alkali-borohydride treatment in reduced conditions.
Neutral and acidic oligosaccharides were fractionated by ion-exchange chromatographies and purified by HPLC.
Each compound was identified by matrix assisted laser desorption ionization-time of flight (MALDI-TOF)
spectrometry, NMR spectroscopy, electrospray ionization-tandem mass spectroscopy (ESI-MS/MS) and
permethylation analyses. This paper reports on the structures of 19 oligosaccharide-alditols, 12 of which have
novel structures. These structures range in size from disaccharide to octasaccharide. Some of them are acidic,
containing either a glucuronic acid or, more frequently, a sulfate group, located either at the 6 position of GlcNAc
or the 3 or 4 positions of Gal. This latter sulfation is novel and has only been characterized in the species
R. arvalis. This structural analysis led to the establishment of several novel carbohydrate structures,
demonstrating the structural diversity and species-specificity of amphibian glycoconjugates.
Keywords: sulfated-oligosaccharides; NMR; amphibian; oviducal mucin.
The amphibian eggs are surrounded by oviducal jellies,which can be structurally and chemically distinct frombetween species [1]. The jelly coats are made up of severalconcentric layers composed of mucin-type glycoproteins,highly O-glycosylated. These glycoproteins have been shown toplay important roles in fertilization [2,3]. Many functions havebeen assigned to the egg jelly components such as spermbinding, sperm capacitation, induction of the acrosomereaction, prevention of species cross-fertilization, the block ofpolyspermy (anurans) and provision of a protective ionicenvironment for the developing embryo [4±7].
Previous studies have shown a species-specific structuraldiversity in carbohydrate chains of amphibian egg jelly coats[8±19]. Consequently, these carbohydrate chains represent newphenotypic markers of amphibian species and from a biologicalpoint of view, can be involved in species-specific interactiongametes or in the specific host±parasite interaction [20]. In thepresent study, we have isolated neutral and acidic carbohydratechains from the oviducal mucins of Rana arvalis, anddetermined the structure of 19 of them.
M A T E R I A L S A N D M E T H O D S
Sampling of jelly coat mucus
Eggs from R. arvalis, collected in France and Russia, wereobtained from natural spawns. The jelly coat material waslyophilized.
Isolation of oligosaccharide-alditols
O-linked oligosaccharides were released from the crudematerial by alkaline degradation in reduced conditions (1 mNaBH4) to obtain stable carbohydrate chains. Total materialwas submitted to a cationic exchange chromatography onDowex 50 � 2 (200±400 mesh, H+ form) to remove residualpeptides. Neutral oligosaccharide-alditols were separated fromacidic oligosaccharide-alditols by anionic exchange chromato-graphy on Dowex 1 � 2 (200±400 mesh, HCOO2 form). Afterwater elution of neutral compounds, acidic oligosaccharideswere desorbed with 50 mm and 100 mm solutions of pyridineacetate buffer (pH 6.7). Neutral fractions were subfractionatedon Bio-Gel P2 column (90 � 1.5 cm) to give three subfractionsFN-I to FN-III. The neutral and acidic compounds were isolatedby HPLC on a primary amine-bonded silica column (Supelco-syl LC-NH2; 4.6 � 250 mm; Supelco Inc, Bellefonte, PA,USA) using acetonitrile/water or acetonitrile/water/30 mmpotassium phosphate buffer (pH 5.2) as eluent, at a flow rateof 1 mL´min21. All the collected fractions were desalted by gelpermeation on a Bio-Gel P2 column (55 � 2 cm), andlyophilized [20]. The different steps of the purification areexplained in detail in many previous papers [12±17].
Eur. J. Biochem. 266, 370±382 (1999) q FEBS 1999
Correspondence to G. Strecker, Laboratoire de Chimie Biologique,
Unite Mixte de Recherche du Centre National de la Recherche Scientifique
n88576, Universite des Sciences et Technologies de Lille (Flandres-Artois),
F-59655 Villeneuve d'Ascq Cedex, France.
Abbreviations: MALDI-TOF, matrix assisted laser desorption
ionization-time of flight; COSY, correlation spectroscopy; ESI-MS/MS,
electrospray ionization-tandem mass spectroscopy; Fuc, fucose; Gal,
galactose; GlcNAc, N-acetylglucosamine; GalNAc, N-acetylgalactosamine;
GalNAc-ol, N-acetylgalactosaminitol; GlcA, glucuronic acid.
(Received 21 July 1999, revised 10 September 1999, accepted
13 September 1999)
q FEBS 1999 Species-specificity of O-glycan chains (Eur. J. Biochem. 266) 371
Carboxyl reduction
Before methylation of compound 50-10, the oligosaccharide-alditol was dissolved in dry dimethylsulfoxide (100 mL).Methyl iodide (100 mL) was added and the solution wasmixed well and kept for 2 h at room temperature in order toconvert the carboxylic acid group into a methyl ester. Afterlyophilization, the derivatized oligosaccharide-alditol wasdissolved in dry methanol (500 mL) and the methyl ester wasreduced using NaBD4 at room temperature for 12 h. After theaddition of acetic acid to pH 6, the sample was dried underreduced pressure. Boric acid was removed as methyl borate bycodistillation (5±6 times) with anhydrous methanol. The driedmaterial was further purified by gel-filtration chromatographyon a column (45 � 0.5 cm) of Sephadex G-10 (Pharmacia)equilibrated with deionized water. The compounds were elutedwith deionized water at a flow rate of 6 mL´h21, and fractionswere analysed for hexose by orcinol/H2SO4 coloration. Thecarbohydrate-containing fractions were pooled, concentratedand then lyophilized.
Methylation
The reduced oligosaccharide (200 mg) was methylatedaccording to Ciucanu and Kerek [21].
Gas chromatography/mass spectrometry (GC/MS)
The partially acetylated methylglycosides were analysed byGC/MS according to Fournet et al. [22].
Matrix-assisted laser desorption mass spectrometry(MALDI-MS)
Molecular masses of the acidic oligosaccharides weremeasured by matrix-assisted laser desorption ionization(MALDI) MS on a Vision 2000 time of flight (TOF)instrument (Finnigan Mat, Bremen, Germany) equipped witha 337-nm UV laser. Samples were dissolved in water at aconcentration of 50±100 pmol´mL21 and 1 mL of thesesolutions were mixed with 1 mL of matrix onto the target,then allowed to crystallize at room temperature. The structureswere analysed in the negative mode using 2,5-dihydroxy-benzoic acid matrix (10 mg´mL21 dissolved in water/ethanol,80 : 20). External calibration was performed using angiotensinI (1296.7 Da) purchased from Sigma. Between 10 and 20 shotswere accumulated for each analysis.
Electrospray mass spectrometry
All MS measurements were carried out in the negative-ionmode on a triple quadrupole instrument (Micromass Ltd,Altrincham, UK). A mixture of polypropylene glycol was usedto calibrate the quadrupole mass spectrometer. The sampleswere dissolved in methanol/water (50 : 50) at a concentrationof 10 pmol´mL21, with 1 mm ammomium acetate. Solutionswere infused using a Havard syringe pump at a flow rate of3 mL´min21. Quadrupole was scanned from 50 to 1200 Da with
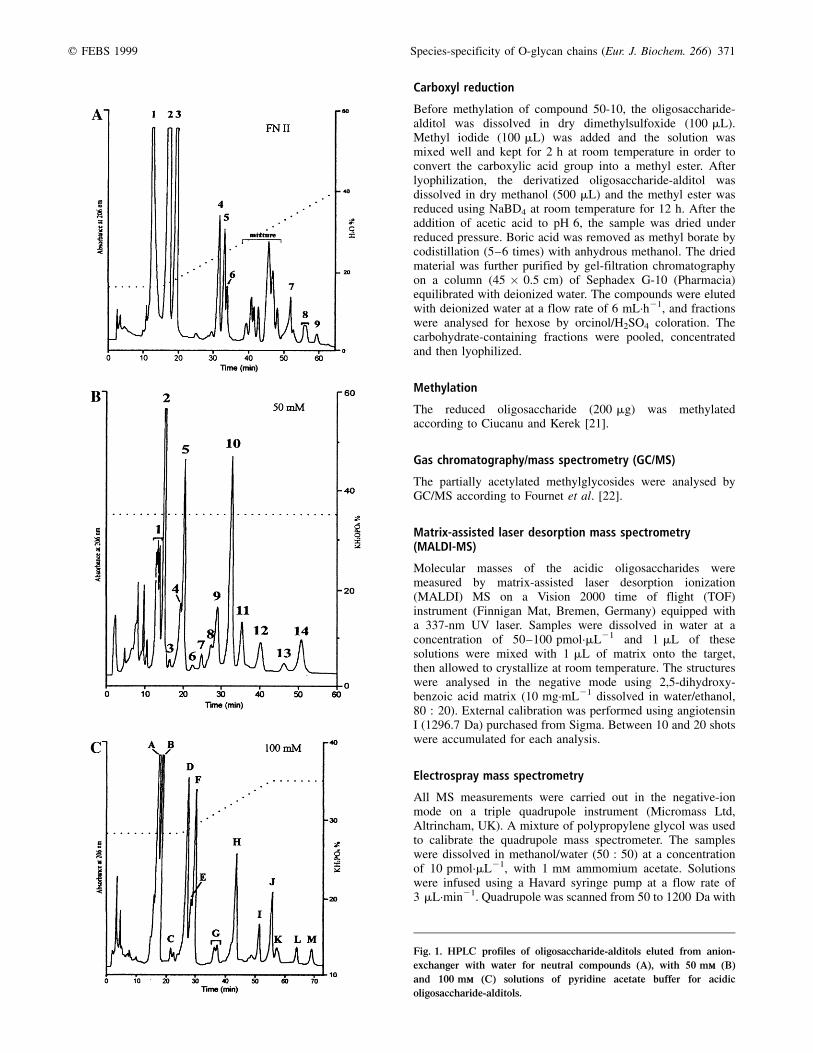
Fig. 1. HPLC profiles of oligosaccharide-alditols eluted from anion-
exchanger with water for neutral compounds (A), with 50 mmm (B)
and 100 mmm (C) solutions of pyridine acetate buffer for acidic
oligosaccharide-alditols.

372 A. Coppin et al. (Eur. J. Biochem. 266) q FEBS 1999
a scan duration of 2 s and a scan delay of 0.1 s. The sampleswere sprayed using a 3.2-kV needle voltage and the decluster-ing (cone) voltage was set at 40 V. For collision-induceddissociation (CID) experiments, the pressure of argon in thecell was 2.4 � 1023 mbar (1.8 � 1023 Torr) and the collisionenergy was set to values ranging from 40 to 70 eV.
400 MHz NMR spectroscopy1H-NMR experiments were performed on a Bruker ASX 400WB spectrometer with a 5-mm 1H-13C mixed probe headoperating in the pulse Fourier transform mode and controlledby an Aspect 3000 computer. Chemical shifts were expressedin p.p.m. downfield from internal sodium 4,4 0-dimethyl-4-silapentane-1-sulfonate but were actually measured byreference to internal acetone (d = 2.225 p.p.m. in D2O at27 8C). The two dimensional homonuclear correlated spectro-scopy (COSY) with simple and double relay transfer wereperformed using Bruker standard pulse sequences library[23,24].
R E S U L T S A N D D I S C U S S I O N
The fractionation of neutral oligosaccharide-alditols on Bio-GelP2 resulted in three subfractions FN-I to FN-III. The analysis ofFN-I revealed oligosaccharide-alditols with molecular massestoo high for an easy HPLC analysis; FN-III will be analysed ata later date. Figure 1 depicts HPLC profiles of neutral fractionFN-II and acidic fractions eluted with 50 mm (fraction 50 mm)and 100 mm (fraction 100 mm) solutions of pyridine acetatebuffer. The system of numbering of individual fractions takesinto account the concentration of buffer elution and also theappearance of HPLC profile [i.e. compound 50-10, oligo-saccharide-alditol desorbed from the anionic exchange resinwith 50 mm pyridine acetate buffer and corresponding to thetenth peak on the HPLC profile; compound FN II-4, neutraloligosaccharide-alditol (second subfraction after Gel filtration)
corresponding to the fourth peak on the HPLC profile]. Thirty-five peaks were isolated but only 19 were characterized.
Neutral compounds FN II-2 to FN II-6 and compound 50-5
These well known compounds are depicted in Fig. 2. Thechemical shifts of their structural groups are identical withthose observed for oligosaccharide-alditols described pre-viously [9,25].
Compound 50-10
The primary structure of this compound was established bycombining the results of methylation analysis, 2D 1H-NMRspectroscopy and MALDI-TOF MS. The MALDI-TOF MSanalysis shows a pseudomolecular ion [M-H]2 at m/z 868(Table 1), which is in agreement with the following molarproportion: HexNAc-ol, Hex, dHex, HexA (1 : 2 : 1 : 1). The2D 1H-NMR spectrum (Fig. 3, Table 2) shows the H-2 and H-5atom resonances of GalNAc-ol at 4.354 and 4.140 p.p.m.,respectively, which shows the existence of core class 1[Gal(b1±3)GalNAc-ol] [25]. According to Koerner et al.[26], the proton vicinal coupling constant pattern attests thepresence of two b-galacto configuration units [3J1,2 = 3J2,3 =large (. 5Hz) and 3J3,4 = 3J4,5 = small (, 5Hz)]. The signalsobserved at dH-1 = 5.379 p.p.m. (3J1,2 = 3.7 Hz), dH-5 =4.276 p.p.m. and dH-6 = 1.238 p.p.m. indicate unambiguouslythe presence of an a-Fuc unit. The NMR parameters (Table 2)of this unit are typical of the (a1±2) linkage to Gal [25]. Onthe basis of our previous work, we suspected that thehexuronic acid was a GlcA unit, sometimes found in otheramphibian carbohydrate molecules [9,13,14]. After carboxy-methylation, reduction with NaBD4, permethylation, meth-anolysis and acetylation, the GC/MS analysis showed thepresence of 6,6 0-D2-(2,3,4,6)-Me4-Glc, showing the existenceof GlcA in nonreducing terminal position. The b-configuration
Table 1. Monosaccharide composition data for oligosaccharide-alditols released from the mucin of the egg jelly coat. The composition of
oligosaccharide-alditols were deduced from [M-H]2 pseudomolecular ions obtained by negative-ion MALDI-TOF spectrometry. Monosaccharide units were
identified on the basis of their vicinal coupling constants by NMR.
Molar ratio
Compounds [M-H]2 GalNAc-ol Gal GlcNAc Fuc GlcA Sulfate or phosphate group
50-5 560 1 1 0 0 1 0
50-7 1121 1 2 1 2 0 1
50-10 868 1 2 0 1 1 0
50-11 1324 1 2 2 2 0 1
50-14 1486 1 3 2 2 0 1
1071 1 2 1 1 1 0
100-A 772 1 2 0 1 0 1
100-B 772 1 2 0 1 0 1
100-D 975 1 2 1 1 0 1
100-E 829 1 2 1 0 0 1
100-F 975 1 2 1 1 0 1
100-G 975 1 2 1 1 0 1
100-H 975 1 2 1 1 0 1
100-I 1121 1 2 1 2 0 1
100-J 1178 1 2 2 1 0 1
100-L 1324 1 2 2 2 0 1
100-M 1340 1 3 2 2 0 1

q FEBS 1999 Species-specificity of O-glycan chains (Eur. J. Biochem. 266) 373
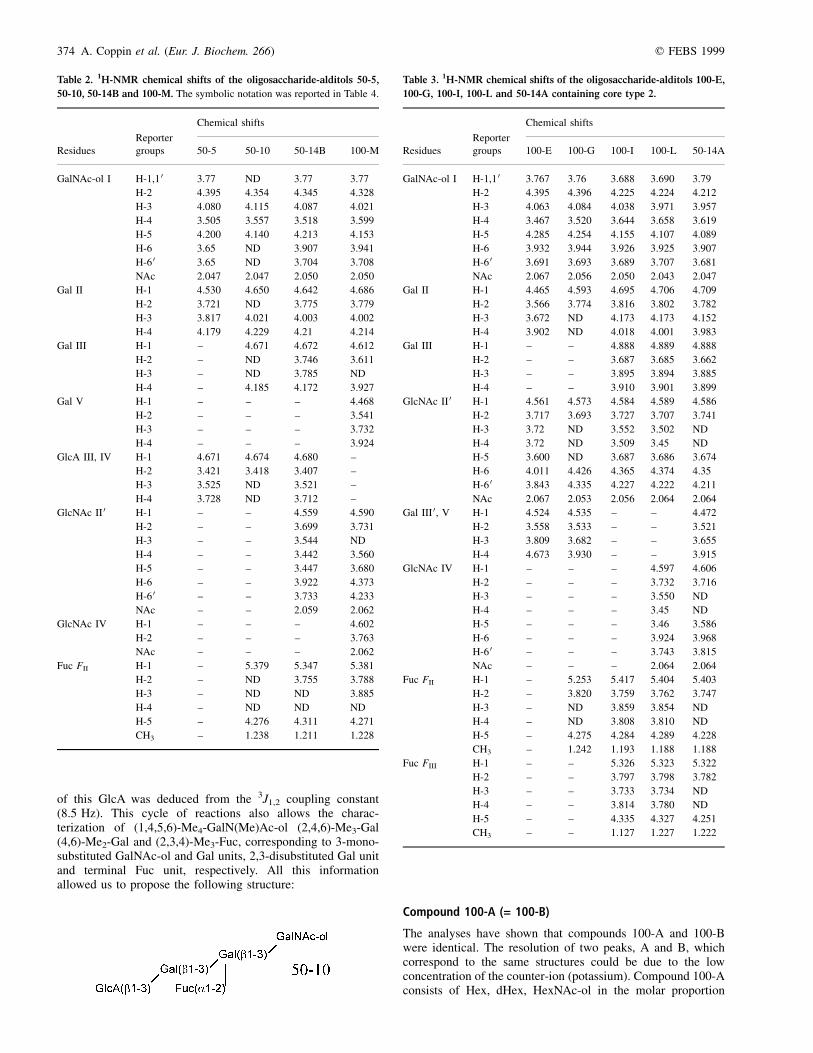
Fig. 2. Summary scheme of the primary structures which are characteristics of R. arvalis.
374 A. Coppin et al. (Eur. J. Biochem. 266) q FEBS 1999
of this GlcA was deduced from the 3J1,2 coupling constant(8.5 Hz). This cycle of reactions also allows the charac-terization of (1,4,5,6)-Me4-GalN(Me)Ac-ol (2,4,6)-Me3-Gal(4,6)-Me2-Gal and (2,3,4)-Me3-Fuc, corresponding to 3-mono-substituted GalNAc-ol and Gal units, 2,3-disubstituted Gal unitand terminal Fuc unit, respectively. All this informationallowed us to propose the following structure:
Compound 100-A (= 100-B)
The analyses have shown that compounds 100-A and 100-Bwere identical. The resolution of two peaks, A and B, whichcorrespond to the same structures could be due to the lowconcentration of the counter-ion (potassium). Compound 100-Aconsists of Hex, dHex, HexNAc-ol in the molar proportion
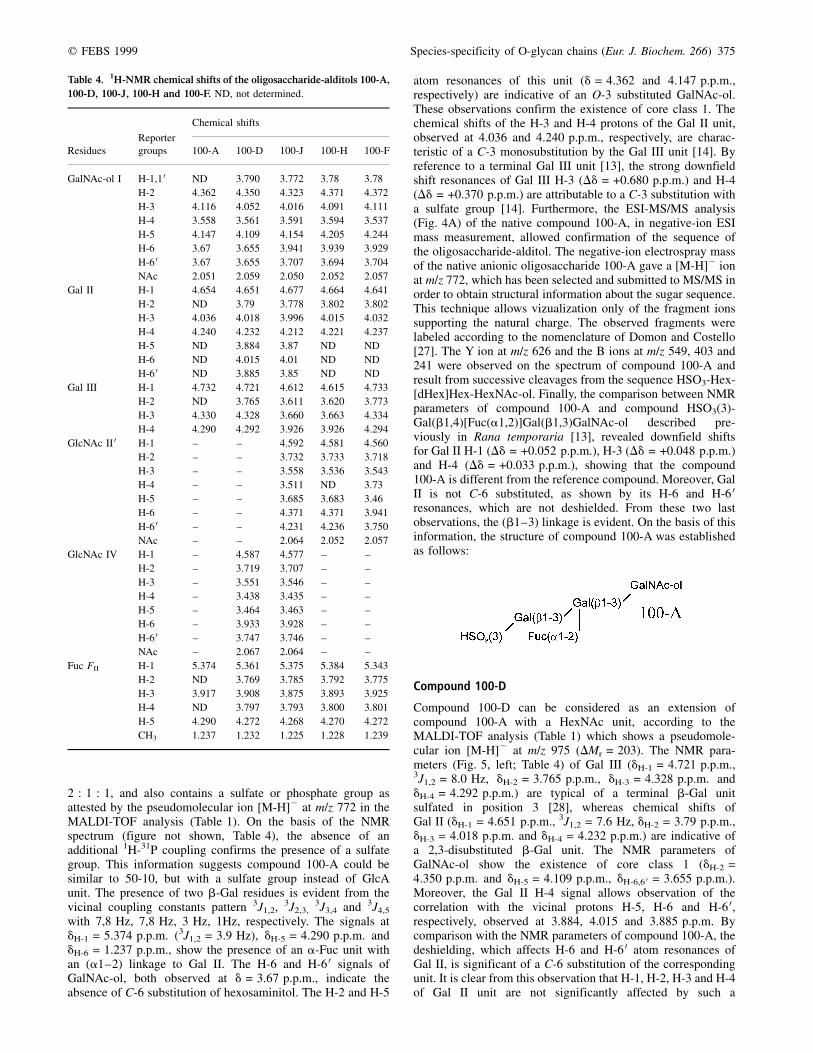
Table 2. 1H-NMR chemical shifts of the oligosaccharide-alditols 50-5,
50-10, 50-14B and 100-M. The symbolic notation was reported in Table 4.
Reporter
Chemical shifts
Residues groups 50-5 50-10 50-14B 100-M
GalNAc-ol I H-1,1 0 3�.77 ND 3�.77 3�.77
H-2 4.395 4�.354 4�.345 4�.328
H-3 4.080 4�.115 4�.087 4�.021
H-4 3.505 3�.557 3�.518 3�.599
H-5 4.200 4�.140 4�.213 4�.153
H-6 3.65 ND 3�.907 3�.941
H-6 0 3.65 ND 3�.704 3�.708
NAc 2.047 2�.047 2�.050 2�.050
Gal II H-1 4�.530 4�.650 4�.642 4�.686
H-2 3.721 ND 3�.775 3�.779
H-3 3.817 4�.021 4�.003 4�.002
H-4 4.179 4�.229 4�.21 4�.214
Gal III H-1 ± 4�.671 4�.672 4�.612
H-2 ± ND 3�.746 3�.611
H-3 ± ND 3�.785 ND
H-4 ± 4�.185 4�.172 3�.927
Gal V H-1 ± ± ± 4�.468
H-2 ± ± ± 3�.541
H-3 ± ± ± 3�.732
H-4 ± ± ± 3�.924
GlcA III, IV H-1 4�.671 4�.674 4�.680 ±
H-2 3.421 3�.418 3�.407 ±
H-3 3.525 ND 3�.521 ±
H-4 3.728 ND 3�.712 ±
GlcNAc II 0 H-1 ± ± 4�.559 4�.590
H-2 ± ± 3�.699 3�.731
H-3 ± ± 3�.544 ND
H-4 ± ± 3�.442 3�.560
H-5 ± ± 3�.447 3�.680
H-6 ± ± 3�.922 4�.373
H-6 0 ± ± 3�.733 4�.233
NAc ± ± 2�.059 2�.062
GlcNAc IV H-1 ± ± ± 4�.602
H-2 ± ± ± 3�.763
NAc ± ± ± 2�.062
Fuc FII H-1 ± 5�.379 5�.347 5�.381
H-2 ± ND 3�.755 3�.788
H-3 ± ND ND 3�.885
H-4 ± ND ND ND
H-5 ± 4�.276 4�.311 4�.271
CH3 ± 1�.238 1�.211 1�.228
Table 3. 1H-NMR chemical shifts of the oligosaccharide-alditols 100-E,
100-G, 100-I, 100-L and 50-14A containing core type 2.
Reporter
Chemical shifts
Residues groups 100-E 100-G 100-I 100-L 50-14A
GalNAc-ol I H-1,1 0 3�.767 3�.76 3�.688 3�.690 3�.79
H-2 4.395 4�.396 4�.225 4�.224 4�.212
H-3 4.063 4�.084 4�.038 3�.971 3�.957
H-4 3.467 3�.520 3�.644 3�.658 3�.619
H-5 4.285 4�.254 4�.155 4�.107 4�.089
H-6 3.932 3�.944 3�.926 3�.925 3�.907
H-6 0 3.691 3�.693 3�.689 3�.707 3�.681
NAc 2.067 2�.056 2�.050 2�.043 2�.047
Gal II H-1 4�.465 4�.593 4�.695 4�.706 4�.709
H-2 3.566 3�.774 3�.816 3�.802 3�.782
H-3 3.672 ND 4�.173 4�.173 4�.152
H-4 3.902 ND 4�.018 4�.001 3�.983
Gal III H-1 ± ± 4�.888 4�.889 4�.888
H-2 ± ± 3�.687 3�.685 3�.662
H-3 ± ± 3�.895 3�.894 3�.885
H-4 ± ± 3�.910 3�.901 3�.899
GlcNAc II 0 H-1 4�.561 4�.573 4�.584 4�.589 4�.586
H-2 3.717 3�.693 3�.727 3�.707 3�.741
H-3 3.72 ND 3�.552 3�.502 ND
H-4 3.72 ND 3�.509 3�.45 ND
H-5 3.600 ND 3�.687 3�.686 3�.674
H-6 4.011 4�.426 4�.365 4�.374 4�.35
H-6 0 3.843 4�.335 4�.227 4�.222 4�.211
NAc 2.067 2�.053 2�.056 2�.064 2�.064
Gal III 0, V H-1 4�.524 4�.535 ± ± 4�.472
H-2 3.558 3�.533 ± ± 3�.521
H-3 3.809 3�.682 ± ± 3�.655
H-4 4.673 3�.930 ± ± 3�.915
GlcNAc IV H-1 ± ± ± 4�.597 4�.606
H-2 ± ± ± 3�.732 3�.716
H-3 ± ± ± 3�.550 ND
H-4 ± ± ± 3�.45 ND
H-5 ± ± ± 3�.46 3�.586
H-6 ± ± ± 3�.924 3�.968
H-6 0 ± ± ± 3�.743 3�.815
NAc ± ± ± 2�.064 2�.064
Fuc FII H-1 ± 5�.253 5�.417 5�.404 5�.403
H-2 ± 3�.820 3�.759 3�.762 3�.747
H-3 ± ND 3�.859 3�.854 ND
H-4 ± ND 3�.808 3�.810 ND
H-5 ± 4�.275 4�.284 4�.289 4�.228
CH3 ± 1�.242 1�.193 1�.188 1�.188
Fuc FIII H-1 ± ± 5�.326 5�.323 5�.322
H-2 ± ± 3�.797 3�.798 3�.782
H-3 ± ± 3�.733 3�.734 ND
H-4 ± ± 3�.814 3�.780 ND
H-5 ± ± 4�.335 4�.327 4�.251
CH3 ± ± 1�.127 1�.227 1�.222
q FEBS 1999 Species-specificity of O-glycan chains (Eur. J. Biochem. 266) 375
2 : 1 : 1, and also contains a sulfate or phosphate group asattested by the pseudomolecular ion [M-H]2 at m/z 772 in theMALDI-TOF analysis (Table 1). On the basis of the NMRspectrum (figure not shown, Table 4), the absence of anadditional 1H-31P coupling confirms the presence of a sulfategroup. This information suggests compound 100-A could besimilar to 50-10, but with a sulfate group instead of GlcAunit. The presence of two b-Gal residues is evident from thevicinal coupling constants pattern 3J1,2, 3J2,3,
3J3,4 and 3J4,5
with 7,8 Hz, 7,8 Hz, 3 Hz, 1Hz, respectively. The signals atdH-1 = 5.374 p.p.m. (3J1,2 = 3.9 Hz), dH-5 = 4.290 p.p.m. anddH-6 = 1.237 p.p.m., show the presence of an a-Fuc unit withan (a1±2) linkage to Gal II. The H-6 and H-6 0 signals ofGalNAc-ol, both observed at d = 3.67 p.p.m., indicate theabsence of C-6 substitution of hexosaminitol. The H-2 and H-5
atom resonances of this unit (d = 4.362 and 4.147 p.p.m.,respectively) are indicative of an O-3 substituted GalNAc-ol.These observations confirm the existence of core class 1. Thechemical shifts of the H-3 and H-4 protons of the Gal II unit,observed at 4.036 and 4.240 p.p.m., respectively, are charac-teristic of a C-3 monosubstitution by the Gal III unit [14]. Byreference to a terminal Gal III unit [13], the strong downfieldshift resonances of Gal III H-3 (Dd = +0.680 p.p.m.) and H-4(Dd = +0.370 p.p.m.) are attributable to a C-3 substitution witha sulfate group [14]. Furthermore, the ESI-MS/MS analysis(Fig. 4A) of the native compound 100-A, in negative-ion ESImass measurement, allowed confirmation of the sequence ofthe oligosaccharide-alditol. The negative-ion electrospray massof the native anionic oligosaccharide 100-A gave a [M-H]2 ionat m/z 772, which has been selected and submitted to MS/MS inorder to obtain structural information about the sugar sequence.This technique allows vizualization only of the fragment ionssupporting the natural charge. The observed fragments werelabeled according to the nomenclature of Domon and Costello[27]. The Y ion at m/z 626 and the B ions at m/z 549, 403 and241 were observed on the spectrum of compound 100-A andresult from successive cleavages from the sequence HSO3-Hex-[dHex]Hex-HexNAc-ol. Finally, the comparison between NMRparameters of compound 100-A and compound HSO3(3)-Gal(b1,4)[Fuc(a1,2)]Gal(b1,3)GalNAc-ol described pre-viously in Rana temporaria [13], revealed downfield shiftsfor Gal II H-1 (Dd = +0.052 p.p.m.), H-3 (Dd = +0.048 p.p.m.)and H-4 (Dd = +0.033 p.p.m.), showing that the compound100-A is different from the reference compound. Moreover, GalII is not C-6 substituted, as shown by its H-6 and H-6 0resonances, which are not deshielded. From these two lastobservations, the (b1±3) linkage is evident. On the basis of thisinformation, the structure of compound 100-A was establishedas follows:
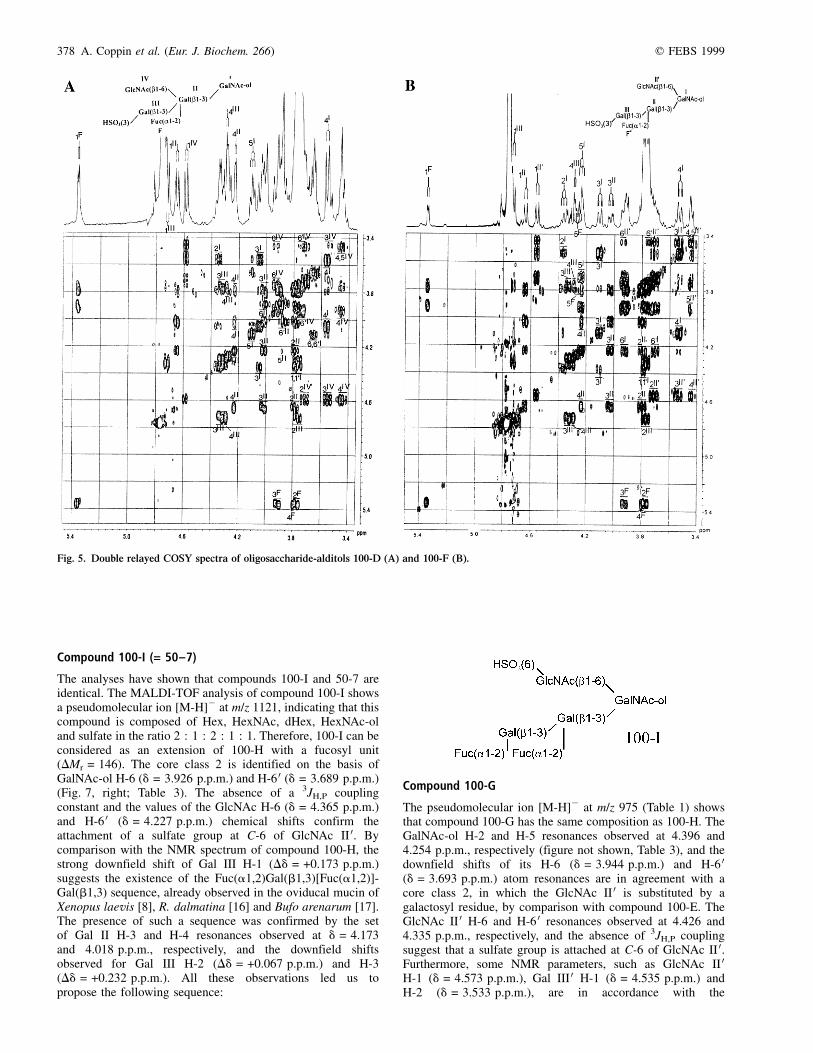
Compound 100-D
Compound 100-D can be considered as an extension ofcompound 100-A with a HexNAc unit, according to theMALDI-TOF analysis (Table 1) which shows a pseudomole-cular ion [M-H]2 at m/z 975 (DMr = 203). The NMR para-meters (Fig. 5, left; Table 4) of Gal III (dH-1 = 4.721 p.p.m.,3J1,2 = 8.0 Hz, dH-2 = 3.765 p.p.m., dH-3 = 4.328 p.p.m. anddH-4 = 4.292 p.p.m.) are typical of a terminal b-Gal unitsulfated in position 3 [28], whereas chemical shifts ofGal II (dH-1 = 4.651 p.p.m., 3J1,2 = 7.6 Hz, dH-2 = 3.79 p.p.m.,dH-3 = 4.018 p.p.m. and dH-4 = 4.232 p.p.m.) are indicative ofa 2,3-disubstituted b-Gal unit. The NMR parameters ofGalNAc-ol show the existence of core class 1 (dH-2 =4.350 p.p.m. and dH-5 = 4.109 p.p.m., dH-6,6 0 = 3.655 p.p.m.).Moreover, the Gal II H-4 signal allows observation of thecorrelation with the vicinal protons H-5, H-6 and H-6 0,respectively, observed at 3.884, 4.015 and 3.885 p.p.m. Bycomparison with the NMR parameters of compound 100-A, thedeshielding, which affects H-6 and H-6 0 atom resonances ofGal II, is significant of a C-6 substitution of the correspondingunit. It is clear from this observation that H-1, H-2, H-3 and H-4of Gal II unit are not significantly affected by such a
Table 4. 1H-NMR chemical shifts of the oligosaccharide-alditols 100-A,
100-D, 100-J, 100-H and 100-F. ND, not determined.
Reporter
Chemical shifts
Residues groups 100-A 100-D 100-J 100-H 100-F
GalNAc-ol I H-1,1 0 ND 3�.790 3�.772 3�.78 3�.78
H-2 4.362 4�.350 4�.323 4�.371 4�.372
H-3 4.116 4�.052 4�.016 4�.091 4�.111
H-4 3.558 3�.561 3�.591 3�.594 3�.537
H-5 4.147 4�.109 4�.154 4�.205 4�.244
H-6 3.67 3�.655 3�.941 3�.939 3�.929
H-6 0 3.67 3�.655 3�.707 3�.694 3�.704
NAc 2.051 2�.059 2�.050 2�.052 2�.057
Gal II H-1 4�.654 4�.651 4�.677 4�.664 4�.641
H-2 ND 3�.79 3�.778 3�.802 3�.802
H-3 4.036 4�.018 3�.996 4�.015 4�.032
H-4 4.240 4�.232 4�.212 4�.221 4�.237
H-5 ND 3�.884 3�.87 ND ND
H-6 ND 4�.015 4�.01 ND ND
H-6 0 ND 3�.885 3�.85 ND ND
Gal III H-1 4�.732 4�.721 4�.612 4�.615 4�.733
H-2 ND 3�.765 3�.611 3�.620 3�.773
H-3 4.330 4�.328 3�.660 3�.663 4�.334
H-4 4.290 4�.292 3�.926 3�.926 4�.294
GlcNAc II 0 H-1 ± ± 4�.592 4�.581 4�.560
H-2 ± ± 3�.732 3�.733 3�.718
H-3 ± ± 3�.558 3�.536 3�.543
H-4 ± ± 3�.511 ND 3�.73
H-5 ± ± 3�.685 3�.683 3�.46
H-6 ± ± 4�.371 4�.371 3�.941
H-6 0 ± ± 4�.231 4�.236 3�.750
NAc ± ± 2�.064 2�.052 2�.057
GlcNAc IV H-1 ± 4�.587 4�.577 ± ±
H-2 ± 3�.719 3�.707 ± ±
H-3 ± 3�.551 3�.546 ± ±
H-4 ± 3�.438 3�.435 ± ±
H-5 ± 3�.464 3�.463 ± ±
H-6 ± 3�.933 3�.928 ± ±
H-6 0 ± 3�.747 3�.746 ± ±
NAc ± 2�.067 2�.064 ± ±
Fuc FII H-1 5�.374 5�.361 5�.375 5�.384 5�.343
H-2 ND 3�.769 3�.785 3�.792 3�.775
H-3 3.917 3�.908 3�.875 3�.893 3�.925
H-4 ND 3�.797 3�.793 3�.800 3�.801
H-5 4.290 4�.272 4�.268 4�.270 4�.272
CH3 1.237 1�.232 1�.225 1�.228 1�.239
376 A. Coppin et al. (Eur. J. Biochem. 266) q FEBS 1999
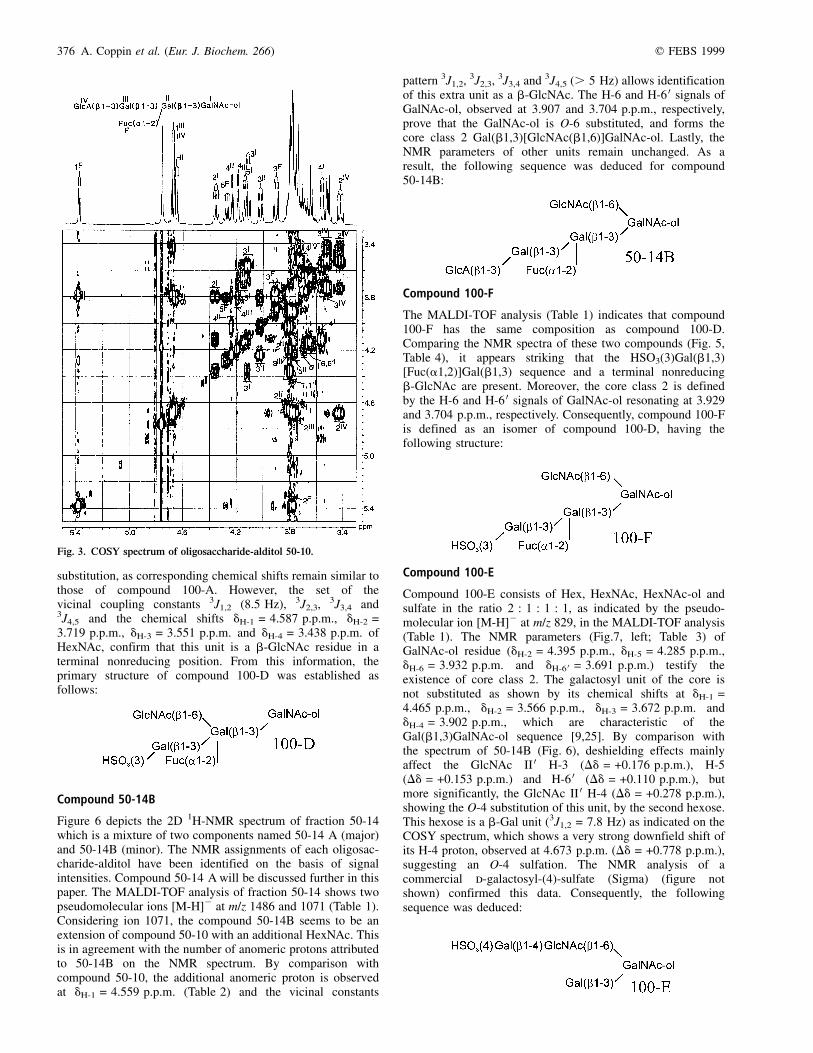
substitution, as corresponding chemical shifts remain similar tothose of compound 100-A. However, the set of thevicinal coupling constants 3J1,2 (8.5 Hz), 3J2,3, 3J3,4 and3J4,5 and the chemical shifts dH-1 = 4.587 p.p.m., dH-2 =3.719 p.p.m., dH-3 = 3.551 p.p.m. and dH-4 = 3.438 p.p.m. ofHexNAc, confirm that this unit is a b-GlcNAc residue in aterminal nonreducing position. From this information, theprimary structure of compound 100-D was established asfollows:
Compound 50-14B
Figure 6 depicts the 2D 1H-NMR spectrum of fraction 50-14which is a mixture of two components named 50-14 A (major)and 50-14B (minor). The NMR assignments of each oligosac-charide-alditol have been identified on the basis of signalintensities. Compound 50-14 A will be discussed further in thispaper. The MALDI-TOF analysis of fraction 50-14 shows twopseudomolecular ions [M-H]2 at m/z 1486 and 1071 (Table 1).Considering ion 1071, the compound 50-14B seems to be anextension of compound 50-10 with an additional HexNAc. Thisis in agreement with the number of anomeric protons attributedto 50-14B on the NMR spectrum. By comparison withcompound 50-10, the additional anomeric proton is observedat dH-1 = 4.559 p.p.m. (Table 2) and the vicinal constants
pattern 3J1,2, 3J2,3, 3J3,4 and 3J4,5 (. 5 Hz) allows identificationof this extra unit as a b-GlcNAc. The H-6 and H-6 0 signals ofGalNAc-ol, observed at 3.907 and 3.704 p.p.m., respectively,prove that the GalNAc-ol is O-6 substituted, and forms thecore class 2 Gal(b1,3)[GlcNAc(b1,6)]GalNAc-ol. Lastly, theNMR parameters of other units remain unchanged. As aresult, the following sequence was deduced for compound50-14B:
Compound 100-F
The MALDI-TOF analysis (Table 1) indicates that compound100-F has the same composition as compound 100-D.Comparing the NMR spectra of these two compounds (Fig. 5,Table 4), it appears striking that the HSO3(3)Gal(b1,3)[Fuc(a1,2)]Gal(b1,3) sequence and a terminal nonreducingb-GlcNAc are present. Moreover, the core class 2 is definedby the H-6 and H-6 0 signals of GalNAc-ol resonating at 3.929and 3.704 p.p.m., respectively. Consequently, compound 100-Fis defined as an isomer of compound 100-D, having thefollowing structure:
Compound 100-E
Compound 100-E consists of Hex, HexNAc, HexNAc-ol andsulfate in the ratio 2 : 1 : 1 : 1, as indicated by the pseudo-molecular ion [M-H]2 at m/z 829, in the MALDI-TOF analysis(Table 1). The NMR parameters (Fig.7, left; Table 3) ofGalNAc-ol residue (dH-2 = 4.395 p.p.m., dH-5 = 4.285 p.p.m.,dH-6 = 3.932 p.p.m. and dH-6 0 = 3.691 p.p.m.) testify theexistence of core class 2. The galactosyl unit of the core isnot substituted as shown by its chemical shifts at dH-1 =4.465 p.p.m., dH-2 = 3.566 p.p.m., dH-3 = 3.672 p.p.m. anddH-4 = 3.902 p.p.m., which are characteristic of theGal(b1,3)GalNAc-ol sequence [9,25]. By comparison withthe spectrum of 50-14B (Fig. 6), deshielding effects mainlyaffect the GlcNAc II 0 H-3 (Dd = +0.176 p.p.m.), H-5(Dd = +0.153 p.p.m.) and H-6 0 (Dd = +0.110 p.p.m.), butmore significantly, the GlcNAc II 0 H-4 (Dd = +0.278 p.p.m.),showing the O-4 substitution of this unit, by the second hexose.This hexose is a b-Gal unit (3J1,2 = 7.8 Hz) as indicated on theCOSY spectrum, which shows a very strong downfield shift ofits H-4 proton, observed at 4.673 p.p.m. (Dd = +0.778 p.p.m.),suggesting an O-4 sulfation. The NMR analysis of acommercial d-galactosyl-(4)-sulfate (Sigma) (figure notshown) confirmed this data. Consequently, the followingsequence was deduced:
Fig. 3. COSY spectrum of oligosaccharide-alditol 50-10.
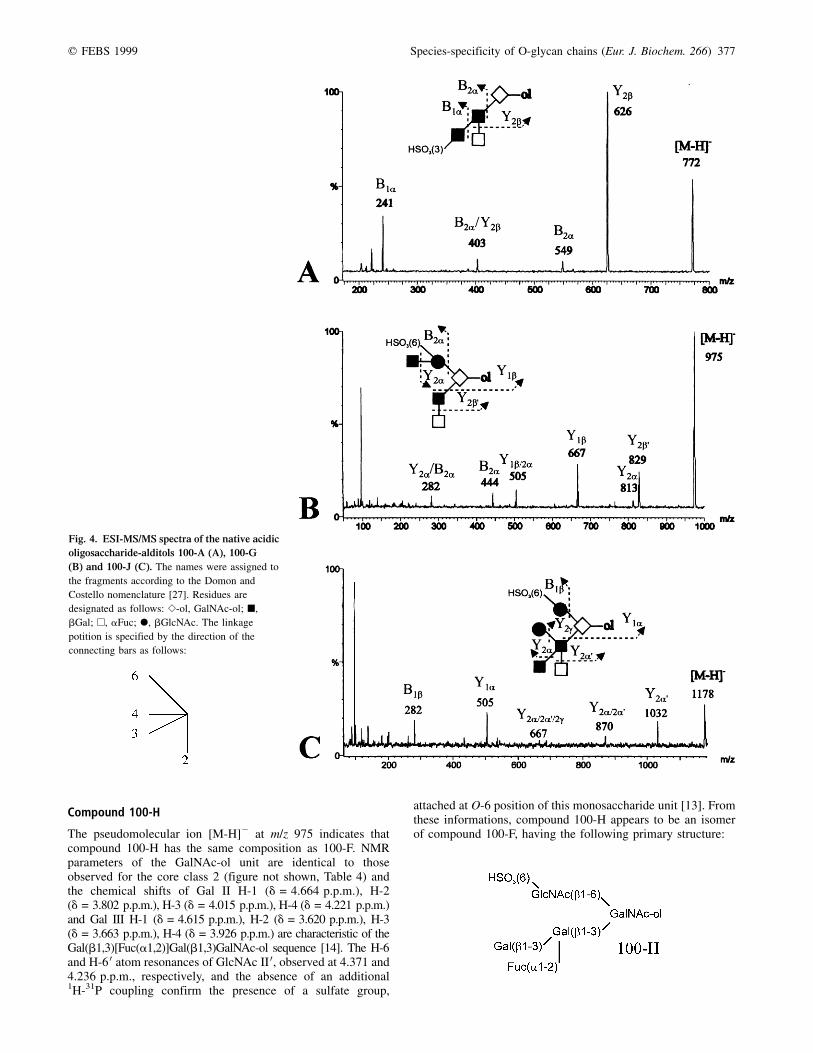
q FEBS 1999 Species-specificity of O-glycan chains (Eur. J. Biochem. 266) 377
Compound 100-H
The pseudomolecular ion [M-H]2 at m/z 975 indicates thatcompound 100-H has the same composition as 100-F. NMRparameters of the GalNAc-ol unit are identical to thoseobserved for the core class 2 (figure not shown, Table 4) andthe chemical shifts of Gal II H-1 (d = 4.664 p.p.m.), H-2(d = 3.802 p.p.m.), H-3 (d = 4.015 p.p.m.), H-4 (d = 4.221 p.p.m.)and Gal III H-1 (d = 4.615 p.p.m.), H-2 (d = 3.620 p.p.m.), H-3(d = 3.663 p.p.m.), H-4 (d = 3.926 p.p.m.) are characteristic of theGal(b1,3)[Fuc(a1,2)]Gal(b1,3)GalNAc-ol sequence [14]. The H-6and H-6 0 atom resonances of GlcNAc II 0, observed at 4.371 and4.236 p.p.m., respectively, and the absence of an additional1H-31P coupling confirm the presence of a sulfate group,
attached at O-6 position of this monosaccharide unit [13]. Fromthese informations, compound 100-H appears to be an isomerof compound 100-F, having the following primary structure:
Fig. 4. ESI-MS/MS spectra of the native acidic
oligosaccharide-alditols 100-A (A), 100-G
(B) and 100-J (C). The names were assigned to
the fragments according to the Domon and
Costello nomenclature [27]. Residues are
designated as follows: S-ol, GalNAc-ol; B,
bGal; A, aFuc; X, bGlcNAc. The linkage
potition is specified by the direction of the
connecting bars as follows:
378 A. Coppin et al. (Eur. J. Biochem. 266) q FEBS 1999
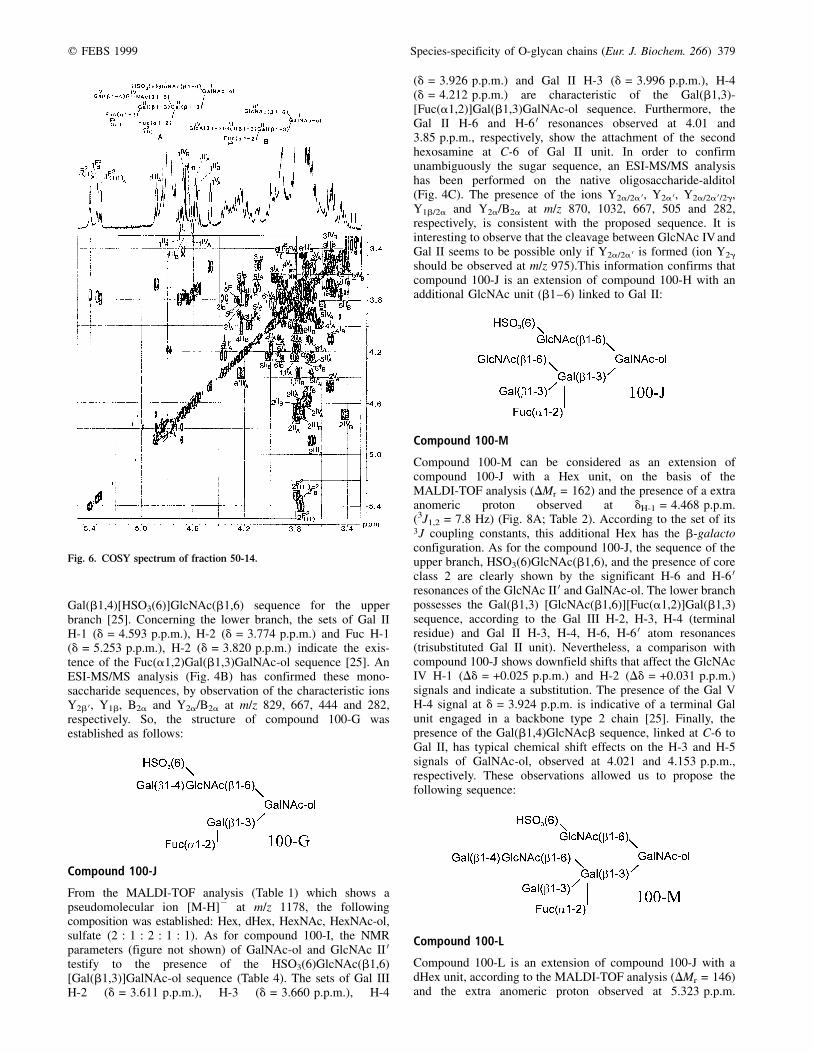
Compound 100-I (= 50±7)
The analyses have shown that compounds 100-I and 50-7 areidentical. The MALDI-TOF analysis of compound 100-I showsa pseudomolecular ion [M-H]2 at m/z 1121, indicating that thiscompound is composed of Hex, HexNAc, dHex, HexNAc-oland sulfate in the ratio 2 : 1 : 2 : 1 : 1. Therefore, 100-I can beconsidered as an extension of 100-H with a fucosyl unit(DMr = 146). The core class 2 is identified on the basis ofGalNAc-ol H-6 (d = 3.926 p.p.m.) and H-6 0 (d = 3.689 p.p.m.)(Fig. 7, right; Table 3). The absence of a 3JH,P couplingconstant and the values of the GlcNAc H-6 (d = 4.365 p.p.m.)and H-6 0 (d = 4.227 p.p.m.) chemical shifts confirm theattachment of a sulfate group at C-6 of GlcNAc II 0. Bycomparison with the NMR spectrum of compound 100-H, thestrong downfield shift of Gal III H-1 (Dd = +0.173 p.p.m.)suggests the existence of the Fuc(a1,2)Gal(b1,3)[Fuc(a1,2)]-Gal(b1,3) sequence, already observed in the oviducal mucin ofXenopus laevis [8], R. dalmatina [16] and Bufo arenarum [17].The presence of such a sequence was confirmed by the setof Gal II H-3 and H-4 resonances observed at d = 4.173and 4.018 p.p.m., respectively, and the downfield shiftsobserved for Gal III H-2 (Dd = +0.067 p.p.m.) and H-3(Dd = +0.232 p.p.m.). All these observations led us topropose the following sequence:
Compound 100-G
The pseudomolecular ion [M-H]2 at m/z 975 (Table 1) showsthat compound 100-G has the same composition as 100-H. TheGalNAc-ol H-2 and H-5 resonances observed at 4.396 and4.254 p.p.m., respectively (figure not shown, Table 3), and thedownfield shifts of its H-6 (d = 3.944 p.p.m.) and H-6 0(d = 3.693 p.p.m.) atom resonances are in agreement with acore class 2, in which the GlcNAc II 0 is substituted by agalactosyl residue, by comparison with compound 100-E. TheGlcNAc II 0 H-6 and H-6 0 resonances observed at 4.426 and4.335 p.p.m., respectively, and the absence of 3JH,P couplingsuggest that a sulfate group is attached at C-6 of GlcNAc II 0.Furthermore, some NMR parameters, such as GlcNAc II 0H-1 (d = 4.573 p.p.m.), Gal III 0 H-1 (d = 4.535 p.p.m.) andH-2 (d = 3.533 p.p.m.), are in accordance with the
Fig. 5. Double relayed COSY spectra of oligosaccharide-alditols 100-D (A) and 100-F (B).
q FEBS 1999 Species-specificity of O-glycan chains (Eur. J. Biochem. 266) 379
Gal(b1,4)[HSO3(6)]GlcNAc(b1,6) sequence for the upperbranch [25]. Concerning the lower branch, the sets of Gal IIH-1 (d = 4.593 p.p.m.), H-2 (d = 3.774 p.p.m.) and Fuc H-1(d = 5.253 p.p.m.), H-2 (d = 3.820 p.p.m.) indicate the exis-tence of the Fuc(a1,2)Gal(b1,3)GalNAc-ol sequence [25]. AnESI-MS/MS analysis (Fig. 4B) has confirmed these mono-saccharide sequences, by observation of the characteristic ionsY2b 0, Y1b, B2a and Y2a/B2a at m/z 829, 667, 444 and 282,respectively. So, the structure of compound 100-G wasestablished as follows:
Compound 100-J
From the MALDI-TOF analysis (Table 1) which shows apseudomolecular ion [M-H]2 at m/z 1178, the followingcomposition was established: Hex, dHex, HexNAc, HexNAc-ol,sulfate (2 : 1 : 2 : 1 : 1). As for compound 100-I, the NMRparameters (figure not shown) of GalNAc-ol and GlcNAc II 0testify to the presence of the HSO3(6)GlcNAc(b1,6)[Gal(b1,3)]GalNAc-ol sequence (Table 4). The sets of Gal IIIH-2 (d = 3.611 p.p.m.), H-3 (d = 3.660 p.p.m.), H-4
(d = 3.926 p.p.m.) and Gal II H-3 (d = 3.996 p.p.m.), H-4(d = 4.212 p.p.m.) are characteristic of the Gal(b1,3)-[Fuc(a1,2)]Gal(b1,3)GalNAc-ol sequence. Furthermore, theGal II H-6 and H-6 0 resonances observed at 4.01 and3.85 p.p.m., respectively, show the attachment of the secondhexosamine at C-6 of Gal II unit. In order to confirmunambiguously the sugar sequence, an ESI-MS/MS analysishas been performed on the native oligosaccharide-alditol(Fig. 4C). The presence of the ions Y2a/2a 0, Y2a 0, Y2a/2a 0/2g,Y1b/2a and Y2a/B2a at m/z 870, 1032, 667, 505 and 282,respectively, is consistent with the proposed sequence. It isinteresting to observe that the cleavage between GlcNAc IV andGal II seems to be possible only if Y2a/2a 0 is formed (ion Y2g
should be observed at m/z 975).This information confirms thatcompound 100-J is an extension of compound 100-H with anadditional GlcNAc unit (b1±6) linked to Gal II:
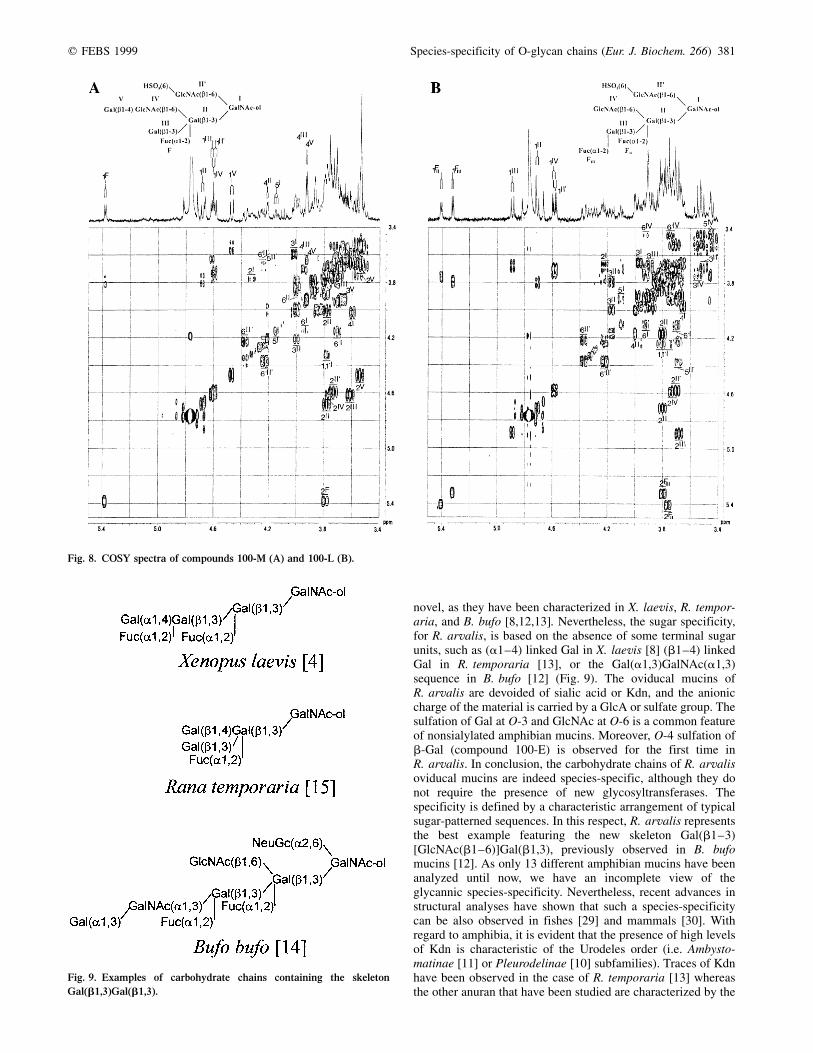
Compound 100-M
Compound 100-M can be considered as an extension ofcompound 100-J with a Hex unit, on the basis of theMALDI-TOF analysis (DMr = 162) and the presence of a extraanomeric proton observed at dH-1 = 4.468 p.p.m.(3J1,2 = 7.8 Hz) (Fig. 8A; Table 2). According to the set of its3J coupling constants, this additional Hex has the b-galactoconfiguration. As for the compound 100-J, the sequence of theupper branch, HSO3(6)GlcNAc(b1,6), and the presence of coreclass 2 are clearly shown by the significant H-6 and H-6 0resonances of the GlcNAc II 0 and GalNAc-ol. The lower branchpossesses the Gal(b1,3) [GlcNAc(b1,6)][Fuc(a1,2)]Gal(b1,3)sequence, according to the Gal III H-2, H-3, H-4 (terminalresidue) and Gal II H-3, H-4, H-6, H-6 0 atom resonances(trisubstituted Gal II unit). Nevertheless, a comparison withcompound 100-J shows downfield shifts that affect the GlcNAcIV H-1 (Dd = +0.025 p.p.m.) and H-2 (Dd = +0.031 p.p.m.)signals and indicate a substitution. The presence of the Gal VH-4 signal at d = 3.924 p.p.m. is indicative of a terminal Galunit engaged in a backbone type 2 chain [25]. Finally, thepresence of the Gal(b1,4)GlcNAcb sequence, linked at C-6 toGal II, has typical chemical shift effects on the H-3 and H-5signals of GalNAc-ol, observed at 4.021 and 4.153 p.p.m.,respectively. These observations allowed us to propose thefollowing sequence:
Compound 100-L
Compound 100-L is an extension of compound 100-J with adHex unit, according to the MALDI-TOF analysis (DMr = 146)and the extra anomeric proton observed at 5.323 p.p.m.
Fig. 6. COSY spectrum of fraction 50-14.
380 A. Coppin et al. (Eur. J. Biochem. 266) q FEBS 1999
(Fig. 8B, Table 3). This additional unit is an (a1±2)linked Fuc residue, according to its NMR parameters(dH-1 = 5.323 p.p.m. (3J1,2 = 4.1 Hz), dH-5 = 4.327 p.p.m. anddH-6 = 1.227 (p.p.m.). By comparison with compound100-J, the strong downfield shift of Gal III H-1(Dd = +0.277 p.p.m.) and the NMR parameters of the twoa-Fuc residues indicate the existence of a Fuc(a1,2)-Gal(b1,3)[Fuc(a1,2)]Gal(b1,3) sequence (see compound 100-I).Furthermore, the presence of a terminal nonreducing GlcNAc IVis evident from the set of its H-2, H-3 and H-4 signals observed,respectively, at 3.732, 3.550 and 3.45 p.p.m. Consequently,compound 100-L has the following primary structure:
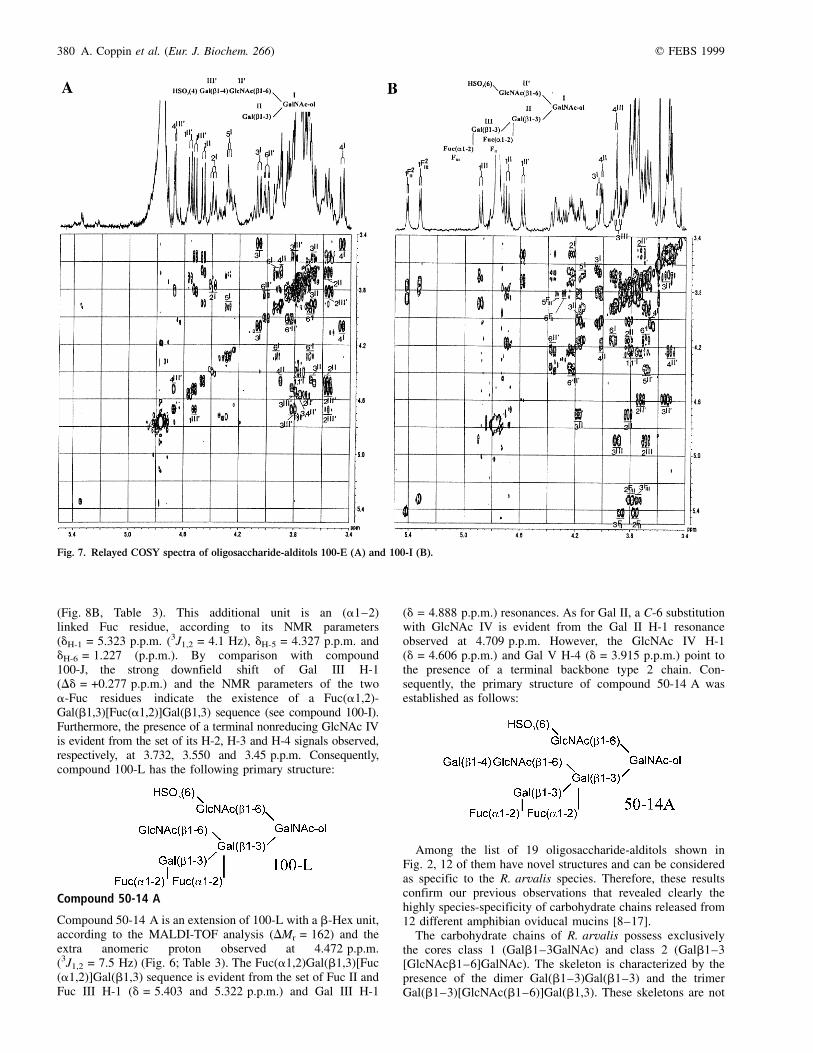
Compound 50-14 A
Compound 50-14 A is an extension of 100-L with a b-Hex unit,according to the MALDI-TOF analysis (DMr = 162) and theextra anomeric proton observed at 4.472 p.p.m.(3J1,2 = 7.5 Hz) (Fig. 6; Table 3). The Fuc(a1,2)Gal(b1,3)[Fuc(a1,2)]Gal(b1,3) sequence is evident from the set of Fuc II andFuc III H-1 (d = 5.403 and 5.322 p.p.m.) and Gal III H-1
(d = 4.888 p.p.m.) resonances. As for Gal II, a C-6 substitutionwith GlcNAc IV is evident from the Gal II H-1 resonanceobserved at 4.709 p.p.m. However, the GlcNAc IV H-1(d = 4.606 p.p.m.) and Gal V H-4 (d = 3.915 p.p.m.) point tothe presence of a terminal backbone type 2 chain. Con-sequently, the primary structure of compound 50-14 A wasestablished as follows:
Among the list of 19 oligosaccharide-alditols shown inFig. 2, 12 of them have novel structures and can be consideredas specific to the R. arvalis species. Therefore, these resultsconfirm our previous observations that revealed clearly thehighly species-specificity of carbohydrate chains released from12 different amphibian oviducal mucins [8±17].
The carbohydrate chains of R. arvalis possess exclusivelythe cores class 1 (Galb1±3GalNAc) and class 2 (Galb1±3[GlcNAcb1±6]GalNAc). The skeleton is characterized by thepresence of the dimer Gal(b1±3)Gal(b1±3) and the trimerGal(b1±3)[GlcNAc(b1±6)]Gal(b1,3). These skeletons are not
Fig. 7. Relayed COSY spectra of oligosaccharide-alditols 100-E (A) and 100-I (B).
q FEBS 1999 Species-specificity of O-glycan chains (Eur. J. Biochem. 266) 381
novel, as they have been characterized in X. laevis, R. tempor-aria, and B. bufo [8,12,13]. Nevertheless, the sugar specificity,for R. arvalis, is based on the absence of some terminal sugarunits, such as (a1±4) linked Gal in X. laevis [8] (b1±4) linkedGal in R. temporaria [13], or the Gal(a1,3)GalNAc(a1,3)sequence in B. bufo [12] (Fig. 9). The oviducal mucins ofR. arvalis are devoided of sialic acid or Kdn, and the anioniccharge of the material is carried by a GlcA or sulfate group. Thesulfation of Gal at O-3 and GlcNAc at O-6 is a common featureof nonsialylated amphibian mucins. Moreover, O-4 sulfation ofb-Gal (compound 100-E) is observed for the first time inR. arvalis. In conclusion, the carbohydrate chains of R. arvalisoviducal mucins are indeed species-specific, although they donot require the presence of new glycosyltransferases. Thespecificity is defined by a characteristic arrangement of typicalsugar-patterned sequences. In this respect, R. arvalis representsthe best example featuring the new skeleton Gal(b1±3)[GlcNAc(b1±6)]Gal(b1,3), previously observed in B. bufomucins [12]. As only 13 different amphibian mucins have beenanalyzed until now, we have an incomplete view of theglycannic species-specificity. Nevertheless, recent advances instructural analyses have shown that such a species-specificitycan be also observed in fishes [29] and mammals [30]. Withregard to amphibia, it is evident that the presence of high levelsof Kdn is characteristic of the Urodeles order (i.e. Ambysto-matinae [11] or Pleurodelinae [10] subfamilies). Traces of Kdnhave been observed in the case of R. temporaria [13] whereasthe other anuran that have been studied are characterized by the
Fig. 8. COSY spectra of compounds 100-M (A) and 100-L (B).
Fig. 9. Examples of carbohydrate chains containing the skeleton
Gal(b1,3)Gal(b1,3).

382 A. Coppin et al. (Eur. J. Biochem. 266) q FEBS 1999
presence of NeuAc, NeuGc, sulfate or GlcA. From a phylo-genetic point of view, the relationships between differentamphibian species should be studied in terms of evolution ofglycosyltransferase genes, leading to the expression of newenzymatic activities or their suppression. As long as thespecies-specificity of carbohydrate chains can be confirmed,the significance of this finding should be come within thecontext of the symbiotic role of carbohydrates during host±parasite interactions [31], or the species-specificity in gameterecognition.
A C K N O W L E D G E M E N T S
This research was supported by the Centre National de la Recherche
Scientifique (UMR 8576, Director Professor Andre Verbert) and by the
MinisteÁre de l'Enseignement SupeÂrieur et de la Recherche.
R E F E R E N C E S
1. Salthe, S.N. (1963) The egg capsules in the amphibia. J. Morphol. 113,
161±171.
2. Gwatkin, R.B.L. (1976) The cell surface in animal embryogenesis and
development. In Fertilization (Poste, G. & Nicolson, G.L., eds),
pp. 1±54. Elsevier/North-Holland Biomedical Press, Amsterdam.
3. Lopo, A.C. (1983) Sperm±egg interactions in invertebrates. In
Mechanism and Control of Animal Fertilization (Hartman, J.F.,
ed.), pp. 269±324. Academic Press, New York.
4. Jego, P., Jolly, J. & Boisseau, C. (1980) Les gangues ovulaires des
amphibiens (proteÂines seÂcreÂteÂes par l'oviducte) et leurs roÃles dans la
feÂcondation. Reproduct. Nutr. Develop. 20, 557±567.
5. Katagari, C. (1987) Role of oviducal secretions in mediating gamete
fusion in anuran amphibians. Zool. Sci. 4, 1±14.
6. Hedrick, J.L. & Nishihara, T. (1991) Structure and function of the
extracellular matrix of anuran eggs. J. Electron Microsc. Technical
17, 319±335.
7. Shimoda, Y., Kitajima, K., Inoue, S. & Inoue, Y. (1994) Isolation,
structural determination and calcium-binding properties of the
major glycoprotein present in Bufo japonicus. Eur. J. Biochem.
223, 223±231.
8. Strecker, G., Wieruszeski, J.M., Plancke, Y. & Boilly, B. (1995)
Primary structure of 12 neutral oligosaccharide-alditols released
from the jelly coats of the anuran Xenopus laevis by reductive
b-elimination. Glycobiology 5, 137±146.
9. Morelle, W. & Strecker, G. (1997) Structural analysis of the
oligosaccharide-alditols released by reductive b-elimination from
the jelly coat of Rana utricularia eggs. Biochem. J. 321, 879±887.
10. Plancke, Y., Wieruszeski, J.M., Boilly, B. & Strecker, G. (1994)
Primary structure of seven new acidic oligosaccharide-alditols from
the egg jelly coats of Axolotl mexicanum and Pleurodeles waltl.
Cienc. Cult. 46, 273±279.
11. Maes, E., Plancke, Y., Delplace, F. & Strecker, G. (1995) Primary
structure of acidic oligosaccharide-alditols derived from the jelly coat
of Ambystoma tigrinum. Eur. J. Biochem. 230, 146±156.
12. Morelle, W. & Strecker, G. (1997) Structural analysis of oligo-
saccharide-alditols released by reductive b-elimination fom oviducal
mucins of Bufo bufo. Glycobiology 7, 777±790.
13. Maes, E., Florea, D., Delplace, F., Lemoine, J., Plancke, Y. &
Strecker, G. (1997) Structural analysis of the oligosaccharide-
alditols released by reductive b-elimination from oviducal mucins
of Rana temporaria. Glycoconjugate J. 14, 127±146.
14. Florea, D., Maes, E. & Strecker, G. (1997) Primary structure of seven
sulfated oligosaccharide-alditols released by reductive b-elimination
from oviducal mucins of Rana temporaria. Carbohydr. Res. 302,
179±189.
15. Maes, E., Florea, D., Coppin, A. & Strecker, G. (1999) Structural
analysis of 20 oligosaccharide-alditols released from the jelly coat of
Rana palustris eggs by reductive b-elimination. Eur. J. Biochem. 263,
1±14.
16. Morelle, W., GuyeÂtant, R. & Strecker, G. (1998) Structural analysis
of oligosaccharide-alditols released by reductive b-elimination
from oviducal mucins of Rana dalmatina. Carbohydr. Res. 306,
435±443.
17. Morelle, W., Cabada, M.O. & Strecker, G. (1998) Structural analysis of
oligosaccharide-alditols released by reductive b-elimination from
the jelly coats of the anuran Bufo arenarum. Eur. J. Biochem.
252, 253±260.
18. Van Halbeek, P.M. (1980) A 360 MHz 1H-NMR study of three
oligosaccharides isolated from cow kappa-casein. Biochem. Biophys.
Acta 623, 295±300.
19. Breg, J., Van Halbeek, H., Vliegenthart, J.F.G., Lamblin, G.,
Houvenaghel, M.C. & Roussel, P. (1987) Structure of sialyl-
oligosaccharides isolated from bronchial mucus glycoproteins of
patients (blood group O) suffering from cystic fibrosis. Eur. J.
Biochem. 168, 57±68.
20. Montreuil, J., Bouquelet, S., Debray, H., Lemoine, J., Michalski, J.C.,
Spik, G. & Strecker, G. (1994) Glycoproteins. In Carbohydrate
Analysis, a Practical Approach (Chaplin, M.F. & Kennedy, J.F., eds),
pp. 181±293. IRL Press, Oxford University Press, Oxford, UK.
21. Ciucanu, I. & Kerek, F. (1984) A simple and rapid method for the
permethylation of carbohydrates. Carbohydr. Res. 131, 209±217.
22. Fournet, B., Strecker, G., Leroy, Y. & Montreuil, J. (1981) Gas-liquid
chromatography and mass spectrometry of methylated and acetylated
methyl glycosides. Application to the structural analysis of
glycoprotein glycans. Anal. Biochem. 116, 489±502.
23. Vliegenthart, J.F.G., Dorland, L. & van Halbeek, H. (1983) High-
resolution, H1-nuclear magnetic resonance spectroscopy as a tool in
the structural analysis of carbohydrates related glycoproteins. Adv.
Carbohydr. Chem. Biochem. 41, 209±374.
24. Bax, A.D. & Davis, D.G. (1985) Practical aspects of two-dimensional
transvers NOE spectroscopy. J. Magn. Reson. 63, 207±213.
25. Kamerling, J.P. & Vliegenthart, J.F.G. (1992) High-resolution1H-nuclear magnetic resonance spectroscopy of oligosaccharide-
alditols released from mucin-type O-glycoproteins. Biol. Magn.
Reson. 10, 1±194.
26. Koerner, T.A.W., Prestegard, J.H. & Yu, R.K. (1987) Oligosaccharide
structure by two-dimensional proton Nuclear Magnetic Resonance
Spectroscopy. Methods Enzymol. 138, 38±59.
27. Domon, B. & Castello, C.E. (1988) A systematic nomenclature for
carbohydrate fragmentations in FAB-MS/MS spectra of glyco-
conjugate. Glycoconjugate J. 5, 397±409.
28. Capon, C., Leroy, Y., Wieruszeski, J.M., Ruant, G., Strecker, G.,
Montreuil, J. & Fournet, B. (1989) Structures of O-glycosidically
linked oligosaccharides isolated from human meconium glyco-
proteins. Eur. J. Biochem. 182, 139±152.
29. Inoue, S., Kanamori, A., Kitajima, K. & Inoue, Y. (1988) Kdn-
glycoprotein: a novel deaminated neuraminic acid-rich glycoprotein
isolated from vitelline envelope of rainbow trout eggs. Biochem.
Biophys. Res. Commun. 153, 172±176.
30. Martensson, S., Levery, S.B., Fang, T.T. & Bendiak, B. (1998) Neutral
noyau oligosaccharides of bovin submaxillary mucin. Use of lead
tetracetate in the cold for establishing branch positions. Eur. J.
Biochem. 258, 603±622.
31. Varki, A. (1993) Biological roles of oligosaccharides: all of the theories
are correct. Glycobiology 3, 97±130.





























