Embed Size (px)
Citation preview

Chromosoma (Berl.) 48, 327--340 (1974) �9 by Springer-Verlag 1974
Analyse des 6changes de chromatides dans les cellules somatiques humaines
T r a i t e m e n t au B U D R (5 b r o m o d ~ o x y u r i d i n e )
e t f luorescence b ico lore p a r l ' A c r i d i n e Orange*
B. Dut r i l l aux , A. M. Fosse, M. Pr ieur et J . Lejeune
Institut de Proggn6se, Paris
Chromat id Exchanges in H u m a n Mitot ic Cells. BUDI~ T r e a t m e n t and Bichromat ic F luorescence b y Acr idine Orange
Abstract. A new technique (BUd~ treatment followed by acridine orange staining) allowing a differentiation of sister chromatids is described. A statistical analysis of 91 human karyotypes gives an estimate of the frequency of exchanges. The mean of sister chromatid exchanges is 27,3 and the minimum number is 11 per cell. - - The frequency of these exchanges is proportional to the relative length of each chromosome, and the accumulation of several exchanges in some segments evokes the possibility of a "negative interference". - - The analysis of endomitoses treated with BUdl~ during at least two generations is not in disagreement with the model of semi-conservative replication of chromosomal DNA, but the modifi- cations of the chromatids may result from a completely different process. - - The frequency of endomitoses is increased by the treatment. These endomitoses allow a very precise analysis of the evolution of the sister chromatid exchanges, during two successive cellular generations.
Introduction
Observ6s pa r Tay lo r et al., d~s 1957, les 6changes de chromat ides su rvenan t sur les chromosomes m6taphas iques ont 6t6 depuis re t rouv4s pa r de n o m b r e u x au teurs chez di /f6rentes esp6ces animalcs ou v6g6tales.
Deux m6thodes, aux pr incipes assez comparables , peuven t 4tre utilis6es. Elles consis tent b, subs t i tuer , ~ la t hymid ine normale de I 'ADN, soit de la t hymid ine tr i t i6e (Taylor , 1957), soit du B U D R ou 5 bromod6- oxyur id ine (Zakharov et Egol ina , 1972).
Le B U D R n~lodifie l ' a spec t des chromosomes m6taphas iques . In t ro - dui t d u r a n t les derni~res heures de culture, co r respondan t 5, la phase S t a rd ive , il crop@he la condensa t ion de cer ta ins segments (Palmer, 1970, Zakha rov et al., 1971), c t l e s deux chromat ides sceurs sont sym6tr iques .
Laiss6 en con tac t p e n d a n t un ou plusieurs jours avec les cellnles en culture, il i ndu i t une asym6tr ic en t re les chromat ides sceurs (Zakharov
* Travail de I'E.I~.A. N o 47 du C.N.R.S. et C.E.A. Contrat iN ~ 293.
"23*

328 B. Dutrillaux et al.
et Egolina, 1972), et l 'on remarque alors qu'il s 'est produi t des 6changes entre elles.
L 'aspec t cytologique des chromosomes ainsi trait6s peut 6tre sensible- men t am61ior6 par l 'emploi de colorants fluorescents, tels l 'acridine orange (Dutrillaux et al., 1973) ou le produi t 33258 Hoechst (Latt , 1973).
Dans ce travail, nous avons analys6 sys t6mat iquement les @changes de chromatides mis en 6vidence par un t ra i tement au B U D R suivi d 'une coloration par l 'acridine orange.
Pa r rappor t s la technique autoradiographique classique, cette m@thode a l ' avantage d'6tre beaucoup plus pr@eise, et exclut la possibilit6 de cassures chromatidiennes radio-induites. Cependant, on ignore si le B U D R n ' indui t pas chimiquement des cassures chromosomiques.
Materiel et m~thodes Toutes les analyses ont 6t4 effectu~es sur des leucocytes humains en culture,
apr~s stimulation par la phytoh4magglutinine. La technique cytologique de base est celle habituellement utilis6e au laboratoire (Turpin et Lejeune, 1965; Dutrillaux et Couturier, 1972).
Le temps de culture a vari6 de 2 s 6 jours. Le BUDI~ a St@ utilis~ s la dose de 200 tzg par ml de milieu et les temps de contact avec les cellules ont 4t~ 6tag,s de 20 heures s 4 jours.
La technique de coloration par ]'acridine orange et d'observation microscopique a ~t4 d6crite par Couturier et al. (1973). Pour mieux faire apparaltre les differences de coloration entre les 2 ehromatides, il est pr@f@rable d'irradier les cellules par la lumigre ultra-violette pendant quelques minutes.
Afin d'identifier les chromosomes, il est possible de les traiter par la m6thode de d~naturation thermique m@nag~e (Dutrillaux et Lejeune, 1971), avant coloration par l'acridine orange.
R~sultats
1. Conditions d' apparition des asymdtries chromatidiennes
Parmi les nombreux essais r6alis@s, nous avons rcmarqu6 l 'appari t ion d'asym@tries chromatidiennes pour des temps de culture en pr6sence de B U D R 6gaux ou sup@rieurs ~ 26 heures. La fr@quencc des mitoses pr6sentant cet aspect est faible de 26 & 30 heures. Elle augmente r@gu- li~rement avec ]e temps de contact , et apr~s plusieurs jours, plus de 50 % des mitoses sont ainsi marqn6es.
2. Apparence des chromosomes asymdtriques
Pour des temps de contact in[@riev_rs & 30 heures, l 'aspect des chromo- somes asym6triques correspond ~ celui d@crit par Zakharov et Egolina (1972). La plus courte des deux chromatides scents cst for tement colorSe, et homog~ne, la plus longue porte des lacunes dont la localisation paral t

Echanges de chromatides dans les cellules humaines 329
comparable s celle observ@ sur les deux chromatides, apr~s un traitement bref (Zakharov et al., 1971, 1974; Dutrillaux et al., 1973).
L'aspect obtenu duns les cultures de cellules sanguines est sensible- ment diff6rent lorsque le temps de contact avec le BUDR est sup6rieur s 30 heures.
Apr~s coloration au Giemsa, une chromatide est homog~ne et fonc6e, l 'autre est 6galement homog~ne, mais un peu plus claire.
Apr~s coloration par l'acridine orange, la chromatide qui prenait fortement le Giemsa 6met une vive fluorescence verte, dont l'intensit6 s'accroit s mesure de l'irradiation ultra-violettc. L'autre chromatide 6met d 'abord une faible fluorescence vert pale, puis progressivement, la coloration rosit jusqu's devenir rouge bistre (fig. 1 a). Cet aspect est particuli~rement favorable pour 4tudier les @hanges de chromatides.
Des diff4rences de longueur semblent persister entre les denx chroma- tides sceurs, la plus courte 4rant celle qui 4met la fluorescence verte.
3. Apparence des chromosomes traitds par le BUDR, puis par la mdthode de ddnaturation thermique
Lorsque les pr6parations sont trait@s par la m6thode de d@aturation thermique m6nag6c, la coloration des chromosomes devient assez paradoxa]e.
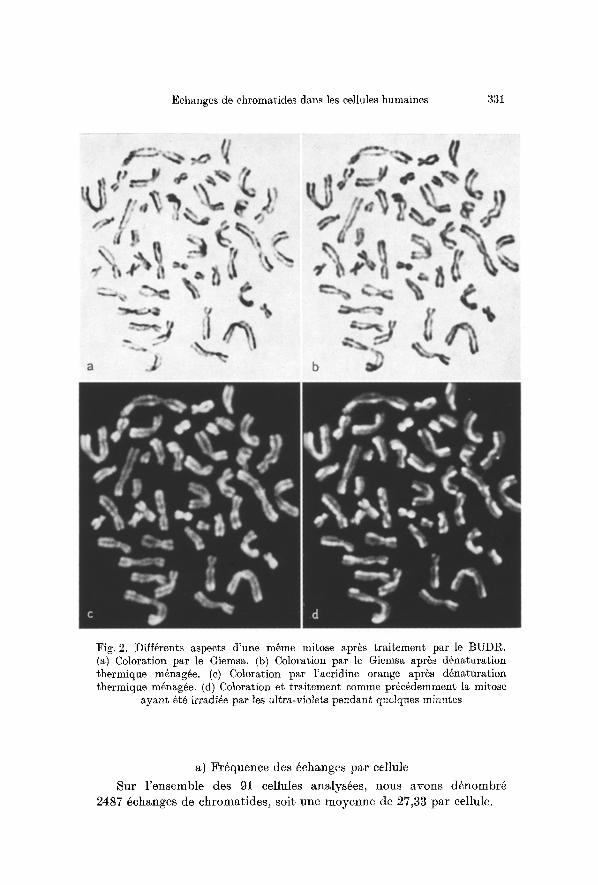
Ainsi, apr~s coloration par le Giemsa, la chromatide qui 6tait ]a plus courte et la plus color@ avant d6naturation devient pale et faible- ment marqn@. Par contre, la chromatide qui 6tait la plus longue et la plus pale porte des bandes R plus intenses et plus 6tendnes (fig. 2a, b).
Si l'on effectue ensuite une coloration par l'acridine orange, on obtient encore un marquage R, mais sym6trique. Puis tr~s rapidement, avec l'irradiation ultra-violette, la chromatide qui 6tait primitivement la plus color6e par ]e Giemsa devient homog~ne et de plus en plus fluorescente en vert. Inversement, l 'autre chromatide perd sa coloration verte et devient rouge et homog~ne (fig. 2 c, d).
Ces diff6rents aspects et ces r6aetions particuli~res au traitement thermique et aux colorations apportent des informations int6ressantes sur les m6canismes du marquage, mais ee point est l 'objet d'6tudes en c o u r s .
Tels qncls, ces aspects permettent une analyse pr6cise des chromatides et la localisation des points d'6changes 6ventuels.
4. Analyse des dchanges de chromatides survenant en une gdndration
Pour connaitre la fr6quence des @hanges de chromatides, nous avons effectu6 le caryotype de 91 cellules, dont 44 de formule 46,XX, et 47 de formule 46,Xu
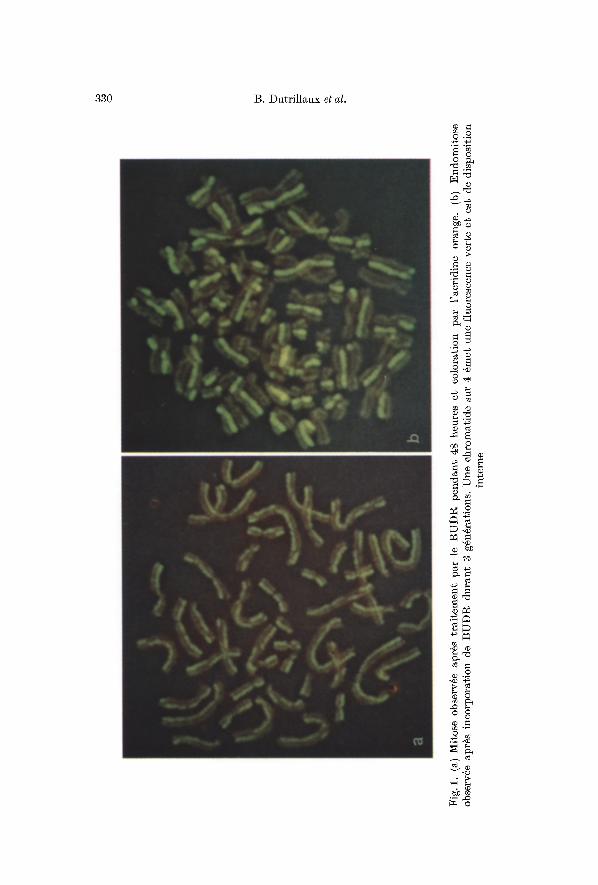
V$
N t
Fig
. 1.
(a
) M
itos
e ob
serv
~e
aprg
s tr
aite
men
t p
ar
le B
UD
R
pen
dan
t 48
heu
res
et
colo
rati
on
par
F
aeri
dine
or
ange
. (b
) E
nd
om
ito
se
obse
rv~e
apr
gs i
ncor
pora
tion
de
BU
DI~
d
ura
nt
3 g6
ngra
tion
s. U
ne
ehro
mat
ide
sur
4 gm
et u
ne f
luor
esee
nee
vert
e et
est
de
disp
osit
ion
inte
rne
Eehanges de ehromatides dans les eellules humaines 331
Fig, 2. Diff@ents aspects d'une m~me mitose aprgs traitement par le BUDR. (a) Coloration par le Giemsa. (b) Coloration par le Giemsa apr~s d6naturation thermique m6nag~e, (c) Coloration par l'aeridine orange aprgs d6naturation thermique m6nag~e, (d) Coloration et traitement comme pr6cgdemment la mitose
ayant ~t6 irradi4e par les ult)ra-violets pendant quelques minutes
a) Fr6quence des 6changes pa r cellule
Sur l ' ensemble des 9I cellules analys6es, nous avons d6nombr6 2487 6changes de ehromat ides , soit une moyenne de 27,33 par cellule;
332 B. Dutrillaux et al.
2 0
1 5
10
10 20 3 0 4 0 5 0 6 0 7 0 80 9 0
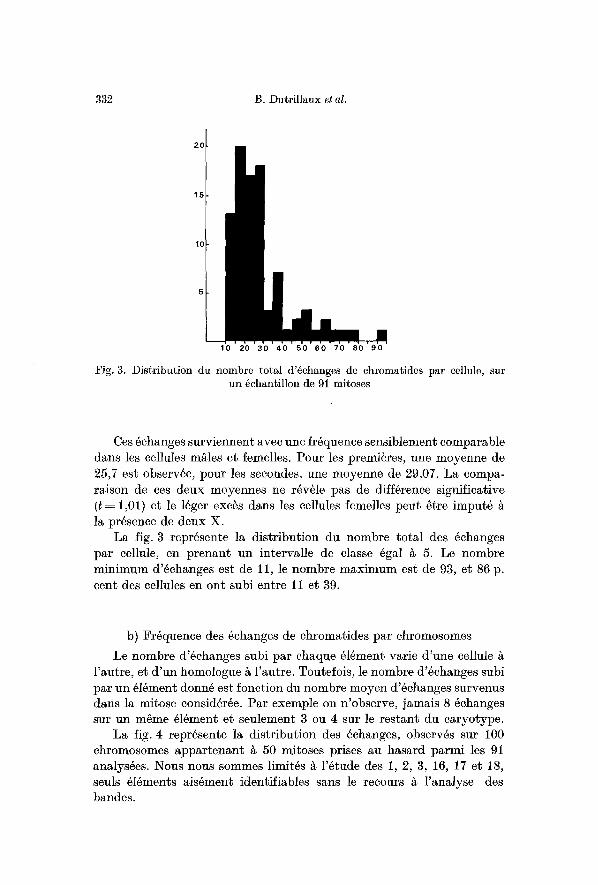
Fig. 3. Distribution du nombre total d'@hanges de chromatides par cellule, sur un 6chantillon de 91 mitoses
Ces 6changes survicnnent avee une fr6quence scnsiblement comparable dans ]es eellules ingles et femelles. Pour les premiSres, une moyenne de 25,7 est observ6e, pour les seeondes, une moyenne de 29,07. La compa- raison de ces deux moyennes ne r~v~le pas de diff6renee signifieative (t : 1,01) et le 16ger excgs dans les eellules femelles peut ~tre imput~ la presence de deux X.
La fig. 3 repr6sente la distribution du nombre total des ~changes par celluIe, en prenant un intervalle de classe ~gal ~ 5. Le hombre minimum d'~ehanges est de 11, le nombre maximum est de 93, et 86 p. cent des cellules an ont subi entre l i et 39.
b) Fr~qnence des ~changes de ehromatides par chromosomes
Le nombre d'~changes subi par chaq~te 616ment varie d'une cellule l'autre, at d 'un homologue s l'autre. Toutefois, le nombre d'6changes subi par un ~lgment donn6 est fonetion du nombre moyen d'~changes survenus dans la mitose consid~rge. Par exemple on n'observe, jamais 8 4changes sur un m6me 416ment et seulement 3 ou 4 sur le restant du earyotype.
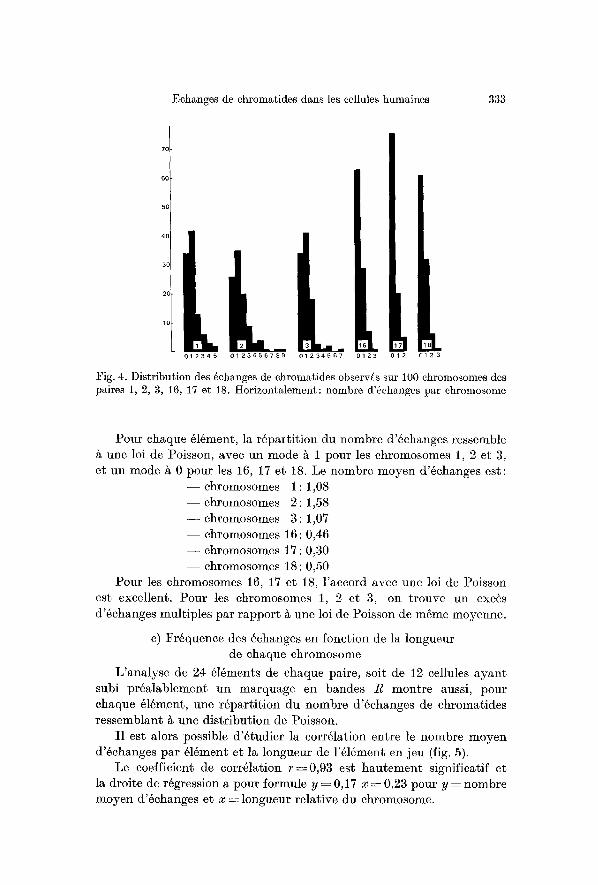
La fig. 4 repr6sente la distribution des @hangcs, observes sur 100 chromosomes appartenant s 50 mitoses prises au hasard parmi les 91 analys~es. Nous nous sommes limit,s g~ l'4tude des 1, 2, 3, 16, 17 et 18, seuls ~16ments ais6ment identifiables sans le recours s l'analyse des bandes.
Eeh~nges de chromatides dans les cellules humaines 333
[[ 0 1 2 3 4 5 0 1 2 3 4 8 6 7 8 9 0 1 2 3 4 8 6 7
Fig. 4. Distribution des 6changes de ehromatides observ(s sur 100 chromosomes des paires 1, 2, 3, 16, 17 et 18. Horizontalement: nombre d'~changes par chromosome
Pour chaque 616ment, la r6pa~t i t ion du nombre d '6ehanges ressemble 5~ une loi de Poisson, avee un mode ~ 1 pour les chromosomes 1, 2 et 3, et un mode/~ 0 pour les 16, 17 et 18. Le hombre moyen d '6changes es t :
- - chromosomes 1 : 1,08 - - chromosomes 2 : 1 , 5 8 - - chromosomes 3 :1 ,07 - - chromosomes 16:0 ,46 - - chromosomes 17:0 ,30 - - chromosomes 18 :0 ,50
Pour les chromosomes 16, i7 et 18, l ' aeeord avee une loi de Poisson est excel lent . Pour les chromosomes 1, 2 et 3, on t rouve un exe6s d '6ehanges mul t ip les pa r r a p p o r t s une loi de Poisson de m4me moyenne .
e) Fr6quenee des 6changes en fonet ion de la longueur de ehaque chromosome
L ' ana ly se de 24 616ments de ehaque paire , soit de 12 eellules a y a n t subi p r6a lab lemen t tin marquage en bandes R mont re aussi, pour ehaque 616ment, une r6par t i t ion du nombre d '6changes de ehromat ides r e s semblan t s une d i s t r ibu t ion de Poisson.
I1 est alors possible d '6 tud ie r la corr61ation ent re le hombre moyen d '6ehanges p a r 616ment et la longueur de l'616ment en jeu (fig. 5).
Le coefficient de eorr61ation r = 0 , 9 3 est h a u t e m e n t s ignif icat if e t la droi te de r6gression a pour formule y = 0,17 x = 0,23 pour y = n o m b r e moyen d '6changes et x = longueur re la t ive du chromosome.
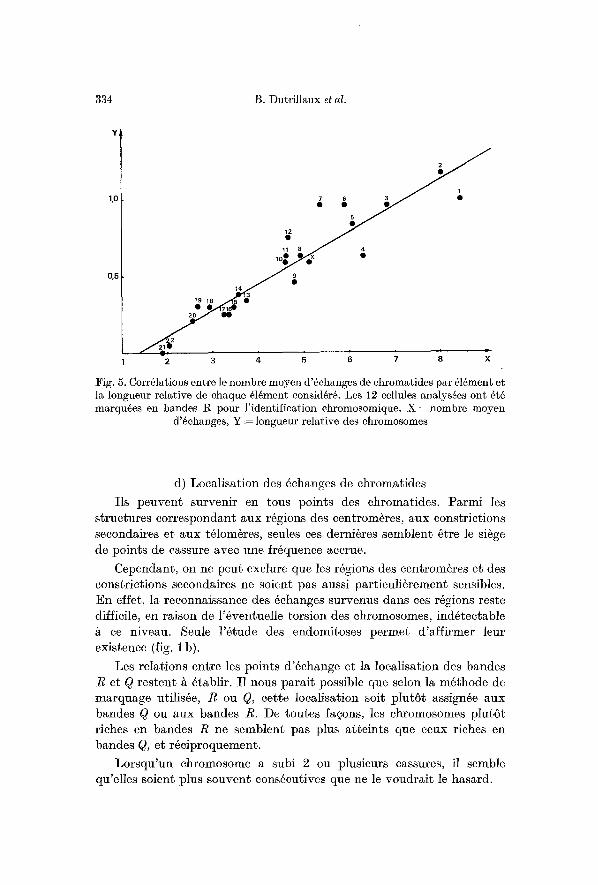
334 B. Dutrillaux et al.
YJ
1,O
0,5
11 8
14 3 19 1 8 ~
Fig. 5. Correlations entre le nombre moyen d'6ehanges de ehromatides par ~16ment et la longueur relative de ehaque ~16ment consid6rg. Les 12 cellules analys@s ont 6t6 marquees en bandes R pour l'identification chromosomique. X =nombre moyen
d'~ehanges, Y ~ longueur relative des chromosomes
d) Loca l i sa t ion des 6changes de chromat ides
I ls peuvcn t surveni r en tous poin ts des chromat ides . P a r m i les s t ruc tures cor respondan t aux r6gions des centrom6res, aux constr ic t ions secondaires et aux t61om6res, seules ees derni6res semblen t 6tre le si6ge de po in ts de cassure avec une fr6quencc accrue.
Cependant , on ne peu t exclure que les r6gions des centrom6res et des constr ic t ions secondaires ne soient pas aussi par t icu l i6 rement sensibles. En effet, la reconnaissance des 6changes survenus duns ces r6gions reste diffieile, en ra ison de l '6ventuel le tors ion des chromosomes, ind6tee table g~ ce niveau. Seule l '6 tude des endomitoses pc rme t d ' a f f i rmer leur exis tence (fig. 1 b).
Les re la t ions ent re les poin ts d 'gehange et la local isat ion des bandcs R et Q res ten t ~ 6tablir . I1 nous pa ra i t possible que selon la m6thode de marquage utilis6e, R ou Q, cet te loeal isat ion soit p lu t6 t assign6e aux bandes Q ou aux bandes R. De tou tes fagons, les chromosomes p lu t6 t r iches en bandes R ne scmblen t pas plus a t t e in t s que ccux riches en bandes Q, e t r6c iproquemcnt .
Lo r squ 'un chromosome a subi 2 ou plusieurs cassures, il semblc qu'el les soient p lus souvent cons6cutives que ne le voudra i t le hasard .

Echanges de chromatides dans les cellules humaines 335
5. Analyse des endoreduplications La fr6quence des endomitoses semble net tement accrue par la
pr6senee de BUDR dans les cultures. Ainsi, pour certaines pr6parations, le sixi6me des cellules en division est endomitotique.
Leur 6rude se r6vble int6ressante, car elle permet de suivre l'6volution des chromatides snr deux g6n6rations successives, avec une pr6cision remarquable.
a) Aspect des endoreduplications apr6s incorporation de BUDt~ durant nne ou deux g6n6rations
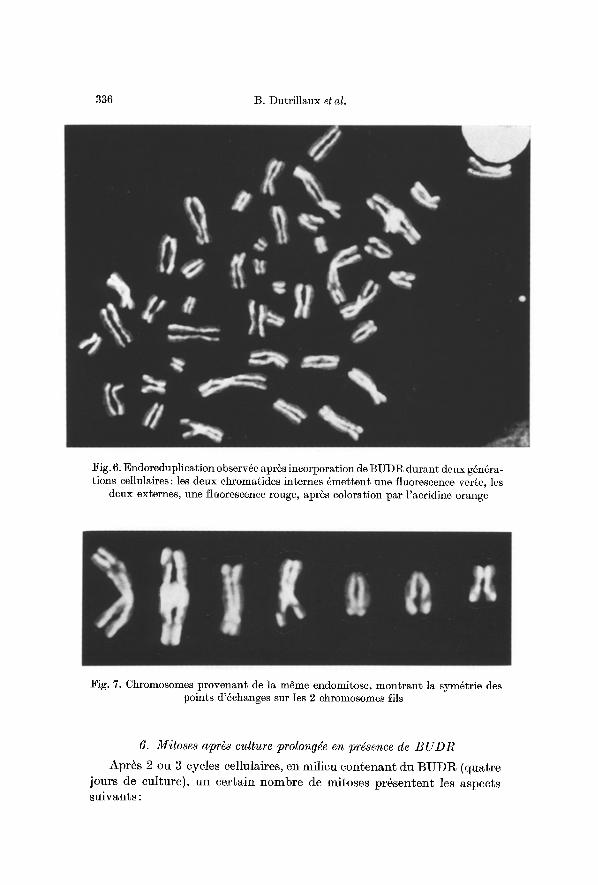
L'aspect des chromosomes (fig. 6) est comparable, dans routes les cellules analys6es. Chaeun des chromosomes formant les t6trades est constitu6 d'une ehromatide verge et d'une ehromatide rouge. Dans la grande majorit6 des eas, les 2 ehromatides verges song internes et ac- eol6es, les rouges s6par6es et externes.
Dans les eas faisant exception, il est ais6 de voir qu'il s'esg produit un, ou des 6changes de chromatides qui ont d6plac6 des segments verts. Cependant, mgme dans ces eas, les 616ments ont tendance s se tordre, de fagon s conserver l 'aceolement des chromatides vertes.
Cette disposition partieuli6re montre que les ehromatides les moins modifi6es, qui auraieng peut-gtre incorpor6 le BUDR sur un seul brin d 'ADN, ont, entre elles, des liaisons plus fortes que les ehromatides les plus modifi6es, qui auraient incorpor6 le BUDt~ sur leurs 2 brins d 'ADN.
Les points d'6changes de chromatides sent souvent tr6s sym6triques sur ehaeun des 2 chromosomes fils, appartenant ~ la m~me t6trade (fig. 7). Cette sym6trie n'existe pas entre chromosomes homologues.
b) Aspect des endoreduplieations apr6s incorporation de BUDR durant 2 ou 3 cycles
Dans ce type de cellule, qui est observ6 pour des temps de culture de 3 ~ 4 jours, en pr6sence de BUDR, l 'aspect est diff6rent du pr6c6dent, car sur les 4 ehromatides de la t6trade, il n 'en persiste plus qu'une poss6dant une fluorescence verge, les 3 autres 6tang de la mgme coloration rouge (fig. 1 b).
Ceci confirme done que les chromatides vertes song bien celles qui ont 6t6 les moins modifi6es par le BUDI~, et qui auraieng conserv6 un brin d 'ADN sans BUDR, scion le mode de replication semi-conservatif.
La chromatide verge est tr6s g6n6ralement interne, eomme dans le eas pr6c6dent, sauf apr6s 6change de chromatides, et l 'on peug alors souvent observer une torsion du chromosome.
336 B. Dutrillaux et al.
Fig. 6. Endoreduplication observge apr6s incorporation de BUDR durant deux g6n6ra- tions cellulaires: les deux chromatides internes 6mettent une fluorescence verte, les
deux externes, une fluorescence rouge, apr~s coloration par l'acridine orange
:Fig. 7. Chromosomes provenant de la mSme endomitose, montrant la sym~trie des points d'~changes sur les 2 chromosomes ills
6. Mitoses apr~s culture prolougde en prdsence de B U D R
Apr~s 2 ou 3 cycles cellulMres, en milieu con tenan t du B U D R (quatre jours de culture), u n certain nombre de mitoses pr6sentent les aspects suivants :

Ech~nges de chromatides duns les cellules humaines 337
Pour chaque paire chromosomique, l 'un des 616ments est constitu6 de deux chromatides uniform6ment rouges, ]'atttre 616ment peut ~tre, soit enti@ement rouge, soit compos6 d'une chromatide bistre et d'une chromatide verte, compl~tement ou partiellement.
Ces images correspondent exaetement au r6sultat d 'une s6gr6gation normale des chromatides asym6triques de ]a g6n6ration ce]lnlaire ant6rieure, et s 'accordent avec une r6plication semi-conservative.
Discussion
1. Les dchanges de chromatides des cellules mitotiques
La r6alit6 des 4changes de chromatides spontan4s est encore discut~e. En effct, prcsque tous les t ravaux effectu~s h ce sujet font intervenir,
pour mettre en 6videnee ces 6changes, l ' incorporation d 'un pr6curseur radio-actif de I 'ADN, comme la thymidine triti4e. On suit que l 'une des propri6t~s des radio-61~mcnts est d'induire des cass~res chromosomiques, qui peuvent entrainer, soit des ~changes de chromatides sceurs, soit d 'autres remaniements tels que des translocations et d614tions.
Ainsi, pour Gibson et Prescott (1972), le hombre d'4changes de chromatides sceurs observ6s 4rant directement proportionnel s la quantit6 de thymidine triti4e incorpor6e, tous les 4changes seraient radio-induits.
Pour d'autres, il existerait un scull au dessus duquel le hombre des 4changes s'accroitrait proportionnellement ~ la dose de pr4cursear radio-actif, et au-dessous duque] le hombre d'~changes serait statisti- quement ind6pendant de la dose.
On ne peut exclure que le BUDR soit capable d'induire les 4changes de chromatides. Duns ce eas, l ' irradiation ne pourrait 4tre invoqu4e, mais un trouble de la replication poarrai t ~tre la cause de (~ changements de copie ~) et non pus de (( cassures-recollements )).
En effet, si le BUDR induisait des cassures, on devrait s 'at tendre observer des remaniements chromosomiques; o1" nous n 'en avons
reconnu aucun sur les 91 caryotypes, dont 12 avee marquage en bandes R, clue nous avons analys6s.
Une comparaison du nombre des 6changes de chromatides observ6s en autor~diographie et apr~s tr~itement par le BUDR et coloration par l 'acridine orange ser~it possible, cn th4orie, mais resterait sans grande signification; le pottvoir analytique de cette derni@e m6thode semblant bien sup@ieur.
Duns l 'hypoth~se off ces 4changes de chromatides ne sont p~s artifi- ciels, il pourrait s'agir d 'un ph~nom~ne constant, voire n4cessaire. En effet, duns notre experience, aucune cellule n 'est exempte d'4changes.

338 B. Dutrillaux et al.
Leur nombre varie de 11 ~ 93, avec une moyenne proche de 27. La survenue de ces 6changes pourrait d6pendre d'une eommande cellulaire, puisque, dans une cellule donn6e, l 'at teinte de chaque chromosome est proportionnelle ~ l 'at teinte de tout le caryotype.
La loealisation des 6changes reste difficile 5~ pr6ciser, en relation avec les structures chromosomiques. Ainsi, la fr6quence de eeux survenant dans les bandes R parait comparable h celle survenant dans les bandes G. De mgme les chromosomes fortement marqu6s en R ne semblent pas atteints plus que eeux fortement marqu6s en G.
Ceci oppose done les 6changes de chromatides aux remaniements ehromosomiques, r6sultant de eassures radio-induites qui si6geraient de pr6f6renee dans les bandes R et qui atteindraient surtout les chromosomes fortement marqu6s en R (I-Iolmberg et Jonasson, 1973 ; Seabright, 1973).
Enfin, il ne semble pas y avoir d 'at teinte privil6gi6e des zones de replication tardive ou pr6coce. Par exemple, dans les cellules ~ deux X, il n 'y a pas d 'atteinte, pr6f6rentielle de l 'un d'eux.
Les seules particularit6s que nous ayons relev6es actuellement sont d'une part, la fr6quence accrue des 6changes de chromatides/~ proximit6 des t61ombres ; d 'autre part , la proximit6 remarquable des points d'6chan- ges accumul6s sur un mgme 616ment.
Ce dernier ph6nom6ne pourrait correspondre ~ u n m6canisme d'<dnterf6rence n6gative)), ce qui opposerait les 6changes de chromatides sceurs, dans les cellules somatiques, s ceux survenant entre chromosomes homologues, dans les cellules germinales.
2. Les dchanges de chromatides des cellules endomitotiques
La fr6quence des endoreduplications est sensiblement accrue par les trai tements au BUDR. De plus, la disposition des chromatides se r6v61e non al6atoire; celles 6mcttant une fluorescence verte, qui sont done les moins modifi6es, sont toujours internes par rapport aux chromatides bistres (fig. l b et 6).
Ces deux ph6nom6nes peuvent avoir une m6me origine: le BUDR emp@herait la s6paration des ehromatides sceurs lors de l 'anaphase. L' interpr6tation de ce m6canisme sera reprise ult6rieurement.
Aprbs incorporation pr6sum6e pendant deux cycles, deux chroma- tides sont vertes, deux sont rouge bistre. On peut alors analyscr les 6changes de chromatides survenus lors de la derni6re g6n6ration cellulaire.
I1 en ressort qu'il existe une tr6s forte tendance ~ la sym6trie d 'un chromosome fils b, l 'autre, s l 'int6rieur d'une m6me t6trade (fig. 7).
Par contre, cette sym6trie ne semble pas exister si l 'on compare les bivalents homologues.

Echanges de chromatides duns les eellules humaines 339
L'analyse des endoreduplieations oh les chromosomes song suppos6s avoir ineorpor6 le BUDR durant trois g6n~rations (une ehromatide verte et trois rouges) semble eonfirmer indireetement cette tendanee. En effet, il est possible de reeonnaitre les 6changes survenus 5~ l 'avant derni@e g6n6ration (la ehromatide verge se r@art i t entre les deux ehromatides sceurs). Si les points d'6ehanges si6geaient au hasard, on devrait pouvoir d6nombrer autant d'6ehanges s ehaque g6n@ation. Au eongraire, si les points d'6ehanges restaieng les mgmes, il ne se produirait en deuxi~me g~n@ation aucune fragmentation de la ehromatide verge. De ee fair, ees 6changes passeraient inapergus. Bien qu 'un gr~s petit nombre d'endo- mitoses de ee type air 6t6 analys6, il semble qua la fr6quenee des 6changes d6teetables soit beaucoup plus faible en derni@e g6n6ration. Par exemple, le seul eas o/1 le nombre total d'6ehanges est inf6rieur s 11 se situe duns eette s@ie.
On peut done snpposer qu'il existe, pour un chromosome donn6, un certain nombre de structures qui permettent , ou favorisent mgme, la survenue des 6changes de ehromatides.
Ces structures se maintiendraient durant deux, plusieurs, voire toutes les g6n@ations eellulaires, de serge que duns un el6ne donn6, tous les chromosomes paternels auraient tendanee ~ subir des 6changes en eertains points; et gous les chromosomes magernels en d 'autres points. Des exp6riences sent en tours pour v@ifier ou infirmer eette hypoth~se.
Enfin, nous avons admis que nous observions l'6volution eellulaire durant une, deux eg trois g6n@ations. Duns cette hypogh~se, la fluores- cence rouge des ehromatides r6sulterait d 'une incorporation de BUDI~ duns les deux brins d 'ADN alors qu'apr~s incorporation s u r u n seal brin, les ehromatides 6mettraient une fluorescence verge. Les modifi- cations ehromosomiques observ6es r6sulgeraient alors de la propri6t6 de replication semi-eonservatriee de I 'ADN, ee qui serait en accord avec les rgsnltats de Taylor et al. (1957).
Des exp@ienees routes r6centes laissent s penser que les modifications tinetoriales observ6es ne correspondent peut gtre pus s un proeessus semi-conservagif. La coloration rouge r6sulterait alors vraisemblablement d'une modification de la structure des prot6ines chromosomiques et non de l'ineorporagion du BUDR duns I 'ADN. Ces exp@ienees serong d6erites duns le d6gail ult@ieurement.
En conclusion, il reste impossible d'affirmer que les ~changes de ehromatides soient un ph6nomgne spontan6, survenant duns les eellules mitotiques.
Cependant leur survenue r6guli~re et proportionnelle s la longueur des chromosomes, en l 'absence de tout ph~nomgne d'irradiation fair penser que le t rai tement au BUDR r6v~le le ph6nomgne d'6change mais ne le provoque pus direetement.

340 B. Dutrillaux et al.
Bibliographie Couturier, J., Dutrillaux, B., Lejeune, J. : Etude des fluorescences sp@ifiques des
bandes R et des bandes Q des chromosomes humains. C. R. Aead. Sci. (Paris) 276, 339--342 (1973)
Dutrillaux, B., Couturier, J. : Techniques d'analyses chromosomiques. In: Biologie g~n6t. Expansion Scient. p. 5--12 (1972)
Dutrillaux, B., Laurent, C., Couturier, J., Lejeune, J.: Coloration par l'acridine orange de chromosomes pr~alablement trait6s par le 5 bromod~oxyuridine (BUDR). C. R. Acad. Sei. (Paris) 276, 3179--3181 (1973)
Dutrillaux, B., Lejeune, J. : Sur une nouvelle technique d'analyse du caryotype humain. C. R. Acad. Sci. (Paris) 272, 2638--2640 (1971)
Gibson, D. A., Prescott, D. H. : Induction of sister ehromatid exchanges in chromo- somes of l~at kangaroo cells by tritium incorporated into DNA. Exp. Cell Res. 74, 3 9 7 ~ 0 2 (1972)
Holmberg, M., Jonasson, J.: Preferential location of X-ray induced chromosome breakage in the l~-bands of human chromosomes. Hereditas (Lund) 74, 57--68 (1973)
Latt, S.A.: Microfluorometric detection of deoxyribonucleic acid replication in human metaphase chromosomes. Proc. nat. Acad. Sci. (Wash.) 70, 3395--3399 (1973)
Palmer, C. G. : 5 bromodeoxyuridine induced constrictions in human chromosomes. Canad. J. Genet. Cytol. 12, 816--830 (1970)
Seabright, M. : High resolution studies on the pattern of induced exchanges in the human karyotype. Chromosoma (Berl.) 40, 33--346 (1973)
Taylor, J. H., Woods, P. S., Hughes, W. L.: The organization and duplication of chromosomes as revealed by auto-radiographic studies using tritium labelled thymidine. Proe. nat. Acad. Sci. (Wash.) 43, 122--127 (1957)
Turpin, R., Lejeune, J. : Les chromosomes humains. Paris: Gauthier Villars Edit. 1965
Zakharov, A.F., Baranovskaya, L.I. , Benjusch, V.A., Demintseva, V.S., Oblapenko, N. G. : Differential spiralization along mammalian mitotic chromo- somes. II. 5-bromodeoxyuridine and 5 bromodeoxycitidine revealed differenti- ation in human chromosomes. Chromosoma (Berl.) 44, 343--360 (1974)
Zakharov, A.F., Egolina, N.A.: Differential spiralization along mammalian mitotic chromosomes I. BUdR revealed differentiation in Chinese hamster chromosomes. Chromosoma (Berl.) 38, 341--365 (1972)
Zakharov, A. F., Seleznev, J. V., Benjusch, V. A., Baranovskaya, L. I., Demintseva, V. S. : Differentiation along human chromosomes in relation to their identi- fication. Exc. Med. Int. Congr. Series 233, 193 (1971)
Rein le Juin 10, 1974 / Aceept~ par H. Bauer Bon • imprimer Juillet 15, 1974
Dr. B. Dutrillaux, Mlle A. M. Fosse, Prof. J. Lejeune, Dr. M. Prieur Chaire de G6n6tique Fondamentale Institut de Prog6n~se 15, rue de l'Ecole de M6decine 75006 Paris France





























