Embed Size (px)
Citation preview

Infection, Genetics and Evolution 12 (2012) 1519–1523
Contents lists available at SciVerse ScienceDirect
Infection, Genetics and Evolution
journal homepage: www.elsevier .com/locate /meegid
Short communication
APOL1 expression is induced by Trypanosoma brucei gambiense infectionbut is not associated with differential susceptibility to sleeping sickness
Hamidou Ilboudo a, David Berthier b, Mamadou Camara c, Oumou Camara c, Jacques Kabore a,Mamadou Leno c, Sow Keletigui d, Isabelle Chantal b, Vincent Jamonneau a,f, Adrien Marie Gaston Belem e,Gérard Cuny f, Bruno Bucheton a,f,⇑a Centre International de Recherche-Développement sur l’Elevage en zone Subhumide (CIRDES), 01 BP 454 Bobo-Dioulasso 01, Burkina Fasob CIRAD, UMR INTERTRYP, Campus International de Baillarguet, F-34398 Montpellier, Francec Programme National de Lutte contre la Trypanosomose Humaine Africaine, BP 851 Conakry, Guinead Institut National de Santé Publique, Laboratoire National de Référence, BP 6366, Conakry, Guineae Université Polytechnique de Bobo-Dioulasso, 01 BP 1091 Bobo-Dioulasso 01, Burkina Fasof Institut de Recherche pour le Développement, Unité Mixte de Recherche IRD-CIRAD 177, TA 207/G, Campus International de Baillarguet, F-34398 Montpellier Cedex 5, France
a r t i c l e i n f o a b s t r a c t
Article history:Received 30 March 2012Received in revised form 21 May 2012Accepted 25 May 2012Available online 9 June 2012
Keywords:Human African trypanosomiasisApolipoprotein L-IExpressionResistance/susceptibility
1567-1348/$ - see front matter � 2012 Elsevier B.V. Ahttp://dx.doi.org/10.1016/j.meegid.2012.05.010
⇑ Corresponding author at: Institut de Recherche pMixte de Recherche IRD-CIRAD 177, TA 207/G, CampuF-34398 Montpellier Cedex 5, France. Tel.: +33 4 67 520.
E-mail address: [email protected] (B. Bucheto
Most African trypanosome species are sensitive to trypanolytic factors (TLFs) present in human serum. Try-panosome lysis was demonstrated to be associated with apolipoprotein L-I (APOL1). Trypanosoma brucei(T. b.) gambiense and Trypanosoma brucei rhodesiense, the two human infective trypanosome species, haveboth developed distinct resistance mechanisms to APOL1 mediated lysis. Whereas T. b. rhodesiense resis-tance is linked with the expression of the serum resistance associated (SRA) protein that interacts withAPOL1 inside the parasite lysosome, inhibiting its lytic action; T. b. gambiense resistance is rather controlledby a reduced expression of the parasite HpHb receptor, limiting APOL1 absorption by trypanosomes. Basedon this last observation we hypothesised that variation in the host APOL1 environment could significantlyalter T. b. gambiense growth and thus resistance/susceptibility to sleeping sickness. To test this hypothesis,we have measured blood APOL1 relative expression in HAT patients, uninfected endemic controls and sero-logically positive subjects (SERO TL+) that are suspected to control infection to parasitological levels thatare undetectable by the available test used in the field. All RNA samples were obtained from medical sur-veys led in the HAT mangrove foci of Coastal Guinea. Results indicate that APOL1 expression is a complextrait dependant on a variety of factors that need to be taken into account in the analysis. Nevertheless, mul-tivariate analysis showed that APOL1 expression levels were significantly higher in both HAT and SERO TL+
subject as compared to endemic controls (p = 0.006). This result suggests that APOL1 expression is likelyinduced by T. b. gambiense, but is not related to resistance/susceptibility in its human host.
� 2012 Elsevier B.V. All rights reserved.
1. Introduction
Apolipoprotein L-I (APOL1) is a human protein that plays a piv-otal role in the interaction of the human host with African trypano-somes conferring innate resistance to most African trypanosomesspecies that otherwise cause fatal disease in livestock (Pays andVanhollebeke, 2008). In human serum, innate resistance is associ-ated with two distinct particles, the trypanosome lytic factor-1(TLF-1), a minor subclass of high-density lipoprotein (HDL) andTLF-2, a large lipid poor complex (Vanhollebeke and Pays, 2010).Both contain APOL1, the trypanolytic component (Vanhamme
ll rights reserved.
our le Développement, Unités International de Baillarguet,9 39 19; fax: +33 4 67 59 39
n).
et al., 2003), and the haptoglobin-related protein (HPR), which bindsfree haemoglobin in blood and facilitate the uptake of APOL1 via atrypanosome haptoglobin–haemoglobin (HbHp) receptor. Whereasrecognition of the HPR–haemoglobin complex by the HbHp receptorseems to be a major pathway for the uptake of TLF-1 (Vanhollebekeet al., 2008) other alternative APOL1 uptake pathways seem to oc-cur. TLF-2 is mainly taken up via another potent but uncharacterisedmechanism and other routes such as a lipoprotein receptor pathwayor fluid phase endocytosis may contribute to the delivery of APOL1to trypanosomes (Vanhollebeke and Pays, 2010; Bullard et al.,2012). Within the acidic lysosome, APOL1 is activated to form mem-brane pores, resulting in ion deregulation that leads to osmoticimbalance, parasite swelling and lysis (Perez-Morga et al., 2005;Vanhollebeke et al., 2007). Two subspecies, Trypanosoma brucei (T.b.) rhodesiense and T. b. gambiense have evolved mechanisms to es-cape lysis from human TLFs and are able to infect human causing

34
74
64
1520 H. Ilboudo et al. / Infection, Genetics and Evolution 12 (2012) 1519–1523
sleeping sickness (Pays et al., 2006). Resistance to human serum ismediated in T. b. rhodesisense by the expression of the serum-resis-tance-associated (SRA) protein which interact specifically with theC-terminal domain of APOL1 inhibiting its lytic action (Lecordieret al., 2009). The SRA gene is absent in T. b. gambiense and resistanceto human serum has been associated with a reduced expression ofthe trypanosome HpHb receptor gene, as compared with the otherT. b. brucei subspecies, which effect is to limit TLF-1 absorption bythe parasite (Kieft et al., 2010). However, T. b. gambiense seems tohave evolved multiple resistance mechanisms, independent of theHpHb receptor pathway, that may differ between group 1 and group2 T. b. gambiense (Capewell et al., 2011).
Whereas APOL1 is critical in determining infectivity to the differ-ent trypanosome species, as shown by the description of an Indiansubject with APOL1 deficiency found to be infected by Trypanosomaevansi (Vanhollebeke et al., 2006), it has also been suggested thatAPOL1 genetic polymorphisms may also influence resistance/suscepti-bility to human infective trypanosomes. A recent study has shownthat two APOL1 polymorphisms within haplotypes harbouring signa-ture of positive selection and located in the APOL1/SRA interactingdomain were associated with kidney diseases in Afro-Americans.Interestingly, sera from carriers of these polymorphisms had somedegree of lytic activity to T. b. rodhesiense. No effects were observedhowever for T. b. gambiense (Genovese et al., 2010). Because T. b.gambiense resistance to human serum seems in part mediated byits ability to avoid APOL1 uptake, we wanted to test here the relationbetween APOL1 expression levels in human populations and resis-tance/susceptibility to T .b. gambiense infections. Noteworthy, anold study performed in Congolese patients and endemic controls,has previously shown that HAT is associated with marked alterationsin the composition and level of host lipoproteins. Nevertheless, nei-ther TLFs nor APOL1 were quantified at that time (Huet et al., 1990).In the framework of the present study, we measured APOL1 relativeexpression from blood RNAs collected within medical surveys led inthe three major HAT foci of Coastal Guinea: Forecariah, Dubreka andBoffa. Previous studies on the genetic characterisation of trypano-somes infecting HAT patients and SERO TL+ individuals in Guineathese last 10 years have shown that all were group 1 T. b. gambiense(Koffi et al., 2009; Kabore et al., 2011). For this study we consideredthree categories of phenotypes according to their HAT infection sta-tus: (i) uninfected endemic controls, negative to the Card Agglutina-tion Test for Trypanosomiasis (CATT), negative to the immunetrypanolysis test (TL) highly specific of T. b. gambiense (Van Meirv-enne et al., 1995; Jamonneau et al., 2010); (ii) HAT patients, for whichthe trypanosome was evidenced by microscopy (mini-anion ex-change centrifugation test (mAECT) (Buscher et al., 2009) and/or di-rect examination of cervical lymph nodes), positive to TL and sortedinto the different disease stages (haemolymphatic stage, early neuro-logical stage, late neurological stage) according to white cell countsin the cerebrospinal fluid (CSF); (iii) individuals displaying strongserological response (CATT plasma dilution end titres 1/8 or higher),positive to TL but in whom parasitological tests were negative (SEROTL+). These individuals maintaining high and specific serological re-sponses to T. b. gambiense are found positive from time to time byPCR and are strongly suspected to control infection to levels thatare below the detection threshold of field parasitological tests (Buch-eton et al., 2011; Ilboudo et al., 2011).
7 16
1 1 1 1
0 10 20 30 40 50 60 70 80 90 100 110
APOL1 expression levels
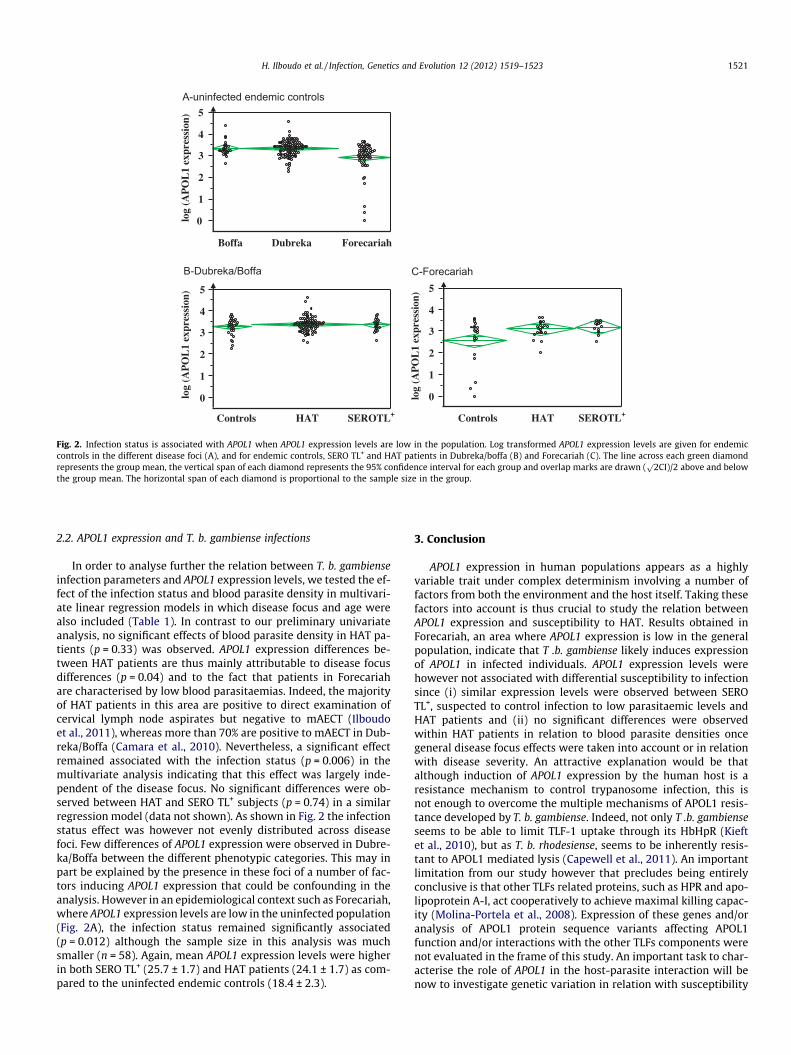
Fig. 1. Distribution of APOL1 expression levels in the study population. Distributionand outlier box plot of APOL1 expression levels (expressed in fold change relative tothe individual with lowest expression; fold change = 1) are shown. Numbers ofindividuals in each APOL1 expression categories are given above the histogram.
2. Result and discussion
2.1. APOL1 expression: a complex phenotypic trait
APOL1 expression levels in the study population were calculatedas the relative expression of APOL1 in comparison to the individualwith the lowest expression level (individual with highest Ct).
APOL1 expression appears to be highly variable across the studypopulation with relative expressions ranging from one to morethan 100 (fig. 1). Moreover, the APOL1 expression distribution inour study population was not normal with skewness toward higherlevels. Linear regressions were thus performed on log transformedAPOL1 expression levels. The univariate linear regression analysisof recorded covariates (gender, age, disease focus and infectioncovariates) as explanatory variables of APOL1 shows that APOL1expression is a complex trait controlled by multiple factors (Table1). Mean APOL1 expression levels were significantly (p = 0.0006)elevated in HAT patients (29.7 ± 1.2) and SERO TL+ (28.1 ± 1.3) ascompared to uninfected endemic controls (24.5 ± 1.6). WithinHAT patients higher expression levels were associated with pa-tients displaying the highest blood trypanosome densities(p = 0.008); >2 trypanosomes observed in the mAECT collector, cor-responding to blood parasitaemia >50 trypanosomes/ml (Buscheret al., 2009; Camara et al., 2010). No association was observed be-tween APOL1 expression levels and the different disease stages.Infection variables were however not the only ones to display sig-nificant effects on APOL1 expression variation. Among those we de-tected: (i) an age effect, with individuals less than 25 years olddisplaying significantly higher APOL1 expression levels than theother age groups (p = 0.002) and (ii) a strong disease focus effectthat was highly significant (p < 0.0001). Mean expression levelswere much lower in Forecariah (22.5 ± 1.2) as compared to thetwo other foci Dubréka (30.5 ± 1.1) and Boffa (30.5 ± 2.3) that arelocated 100 and 150 km along the Guinean cost. Furthermore,the focus effect was the most significant amongst uninfected ende-mic controls (p = 0.0004) indicating that in between foci differ-ences are largely independent of T. b. gambiense infectionparameters (Fig 2A). The reason underlying these APOL1 expressiondifferences between foci are unknown but could possibly be re-lated to differences in the nutritional regimen (lipid consumption,vitamin D. . .), to the existence of other infections inducing APOL1expression, to the prevalence of different pathologies associatedwith increased apolipoprotein levels or to genetic polymorphismsin lipid metabolism genes. Indeed, significant correlations havepreviously been observed between plasma apolipoprotein L con-centrations and triglycerides and cholesterol levels in normolipi-demic subjects. Plasma APOL proteins concentrations were alsofound to be elevated in hyperlipidemic and diabetic patients(Duchateau et al., 2000). Furthermore it was suggested that controlof APOL proteins expression is in part mediated at the transcriptionlevels through the presence of putative sterol regulatory elementspresent in the promoter of the APOL gene family (Duchateau et al.,2001).
0
1
2
3
4
5
Boffa Dubreka Forecariah
0
1
2
3
4
5
Controls HAT SEROTL+
0
1
2
3
4
5
A-uninfected endemic controls
B-Dubreka/Boffa C-Forecariah
log
(AP
OL
1 ex
pres
sion
)
Controls HAT SEROTL+
log
(AP
OL
1 ex
pres
sion
)
log
(AP
OL
1 ex
pres
sion
)
Fig. 2. Infection status is associated with APOL1 when APOL1 expression levels are low in the population. Log transformed APOL1 expression levels are given for endemiccontrols in the different disease foci (A), and for endemic controls, SERO TL+ and HAT patients in Dubreka/boffa (B) and Forecariah (C). The line across each green diamondrepresents the group mean, the vertical span of each diamond represents the 95% confidence interval for each group and overlap marks are drawn (
p2CI)/2 above and below
the group mean. The horizontal span of each diamond is proportional to the sample size in the group.
H. Ilboudo et al. / Infection, Genetics and Evolution 12 (2012) 1519–1523 1521
2.2. APOL1 expression and T. b. gambiense infections
In order to analyse further the relation between T. b. gambienseinfection parameters and APOL1 expression levels, we tested the ef-fect of the infection status and blood parasite density in multivari-ate linear regression models in which disease focus and age werealso included (Table 1). In contrast to our preliminary univariateanalysis, no significant effects of blood parasite density in HAT pa-tients (p = 0.33) was observed. APOL1 expression differences be-tween HAT patients are thus mainly attributable to disease focusdifferences (p = 0.04) and to the fact that patients in Forecariahare characterised by low blood parasitaemias. Indeed, the majorityof HAT patients in this area are positive to direct examination ofcervical lymph node aspirates but negative to mAECT (Ilboudoet al., 2011), whereas more than 70% are positive to mAECT in Dub-reka/Boffa (Camara et al., 2010). Nevertheless, a significant effectremained associated with the infection status (p = 0.006) in themultivariate analysis indicating that this effect was largely inde-pendent of the disease focus. No significant differences were ob-served between HAT and SERO TL+ subjects (p = 0.74) in a similarregression model (data not shown). As shown in Fig. 2 the infectionstatus effect was however not evenly distributed across diseasefoci. Few differences of APOL1 expression were observed in Dubre-ka/Boffa between the different phenotypic categories. This may inpart be explained by the presence in these foci of a number of fac-tors inducing APOL1 expression that could be confounding in theanalysis. However in an epidemiological context such as Forecariah,where APOL1 expression levels are low in the uninfected population(Fig. 2A), the infection status remained significantly associated(p = 0.012) although the sample size in this analysis was muchsmaller (n = 58). Again, mean APOL1 expression levels were higherin both SERO TL+ (25.7 ± 1.7) and HAT patients (24.1 ± 1.7) as com-pared to the uninfected endemic controls (18.4 ± 2.3).
3. Conclusion
APOL1 expression in human populations appears as a highlyvariable trait under complex determinism involving a number offactors from both the environment and the host itself. Taking thesefactors into account is thus crucial to study the relation betweenAPOL1 expression and susceptibility to HAT. Results obtained inForecariah, an area where APOL1 expression is low in the generalpopulation, indicate that T .b. gambiense likely induces expressionof APOL1 in infected individuals. APOL1 expression levels werehowever not associated with differential susceptibility to infectionsince (i) similar expression levels were observed between SEROTL+, suspected to control infection to low parasitaemic levels andHAT patients and (ii) no significant differences were observedwithin HAT patients in relation to blood parasite densities oncegeneral disease focus effects were taken into account or in relationwith disease severity. An attractive explanation would be thatalthough induction of APOL1 expression by the human host is aresistance mechanism to control trypanosome infection, this isnot enough to overcome the multiple mechanisms of APOL1 resis-tance developed by T. b. gambiense. Indeed, not only T .b. gambienseseems to be able to limit TLF-1 uptake through its HbHpR (Kieftet al., 2010), but as T. b. rhodesiense, seems to be inherently resis-tant to APOL1 mediated lysis (Capewell et al., 2011). An importantlimitation from our study however that precludes being entirelyconclusive is that other TLFs related proteins, such as HPR and apo-lipoprotein A-I, act cooperatively to achieve maximal killing capac-ity (Molina-Portela et al., 2008). Expression of these genes and/oranalysis of APOL1 protein sequence variants affecting APOL1function and/or interactions with the other TLFs components werenot evaluated in the frame of this study. An important task to char-acterise the role of APOL1 in the host-parasite interaction will benow to investigate genetic variation in relation with susceptibility
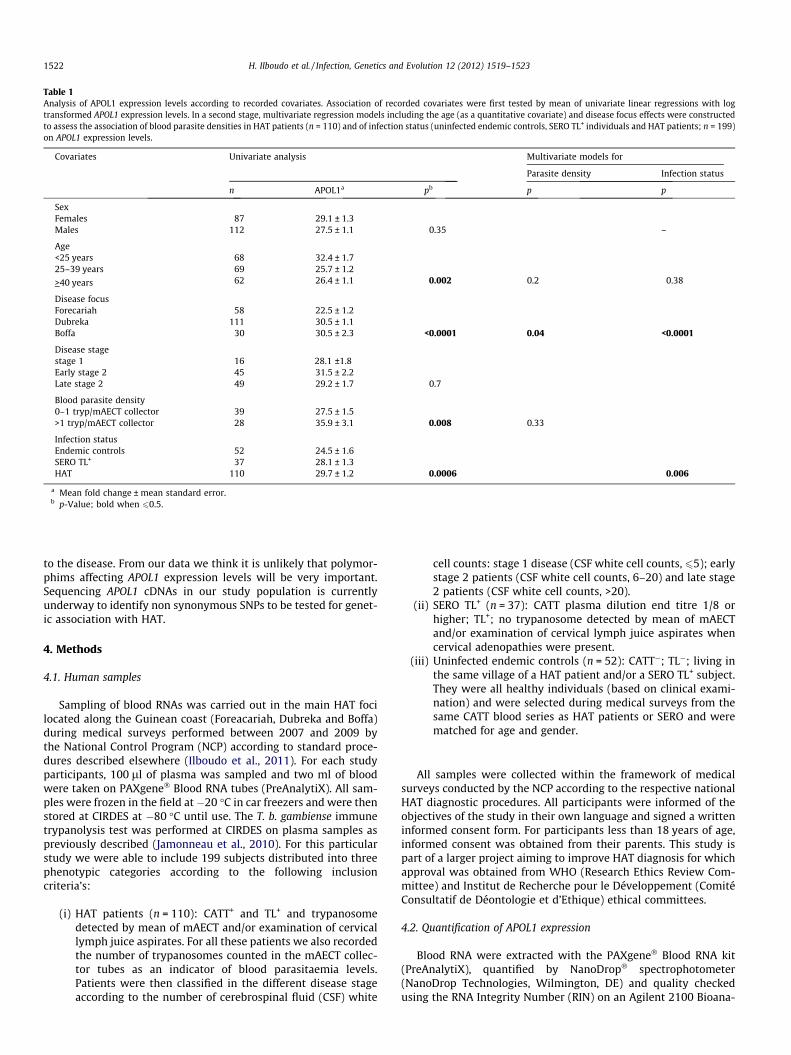
Table 1Analysis of APOL1 expression levels according to recorded covariates. Association of recorded covariates were first tested by mean of univariate linear regressions with logtransformed APOL1 expression levels. In a second stage, multivariate regression models including the age (as a quantitative covariate) and disease focus effects were constructedto assess the association of blood parasite densities in HAT patients (n = 110) and of infection status (uninfected endemic controls, SERO TL+ individuals and HAT patients; n = 199)on APOL1 expression levels.
Covariates Univariate analysis Multivariate models for
Parasite density Infection status
n APOL1a pb p p
SexFemales 87 29.1 ± 1.3Males 112 27.5 ± 1.1 0.35 –
Age<25 years 68 32.4 ± 1.725–39 years 69 25.7 ± 1.2
>40 years 62 26.4 ± 1.1 0.002 0.2 0.38
Disease focusForecariah 58 22.5 ± 1.2Dubreka 111 30.5 ± 1.1Boffa 30 30.5 ± 2.3 <0.0001 0.04 <0.0001
Disease stagestage 1 16 28.1 ±1.8Early stage 2 45 31.5 ± 2.2Late stage 2 49 29.2 ± 1.7 0.7
Blood parasite density0–1 tryp/mAECT collector 39 27.5 ± 1.5>1 tryp/mAECT collector 28 35.9 ± 3.1 0.008 0.33
Infection statusEndemic controls 52 24.5 ± 1.6SERO TL+ 37 28.1 ± 1.3HAT 110 29.7 ± 1.2 0.0006 0.006
a Mean fold change ± mean standard error.b p-Value; bold when 60.5.
1522 H. Ilboudo et al. / Infection, Genetics and Evolution 12 (2012) 1519–1523
to the disease. From our data we think it is unlikely that polymor-phims affecting APOL1 expression levels will be very important.Sequencing APOL1 cDNAs in our study population is currentlyunderway to identify non synonymous SNPs to be tested for genet-ic association with HAT.
4. Methods
4.1. Human samples
Sampling of blood RNAs was carried out in the main HAT focilocated along the Guinean coast (Foreacariah, Dubreka and Boffa)during medical surveys performed between 2007 and 2009 bythe National Control Program (NCP) according to standard proce-dures described elsewhere (Ilboudo et al., 2011). For each studyparticipants, 100 ll of plasma was sampled and two ml of bloodwere taken on PAXgene� Blood RNA tubes (PreAnalytiX). All sam-ples were frozen in the field at�20 �C in car freezers and were thenstored at CIRDES at �80 �C until use. The T. b. gambiense immunetrypanolysis test was performed at CIRDES on plasma samples aspreviously described (Jamonneau et al., 2010). For this particularstudy we were able to include 199 subjects distributed into threephenotypic categories according to the following inclusioncriteria’s:
(i) HAT patients (n = 110): CATT+ and TL+ and trypanosomedetected by mean of mAECT and/or examination of cervicallymph juice aspirates. For all these patients we also recordedthe number of trypanosomes counted in the mAECT collec-tor tubes as an indicator of blood parasitaemia levels.Patients were then classified in the different disease stageaccording to the number of cerebrospinal fluid (CSF) white
cell counts: stage 1 disease (CSF white cell counts, 65); earlystage 2 patients (CSF white cell counts, 6–20) and late stage2 patients (CSF white cell counts, >20).
(ii) SERO TL+ (n = 37): CATT plasma dilution end titre 1/8 orhigher; TL+; no trypanosome detected by mean of mAECTand/or examination of cervical lymph juice aspirates whencervical adenopathies were present.
(iii) Uninfected endemic controls (n = 52): CATT�; TL�; living inthe same village of a HAT patient and/or a SERO TL+ subject.They were all healthy individuals (based on clinical exami-nation) and were selected during medical surveys from thesame CATT blood series as HAT patients or SERO and werematched for age and gender.
All samples were collected within the framework of medicalsurveys conducted by the NCP according to the respective nationalHAT diagnostic procedures. All participants were informed of theobjectives of the study in their own language and signed a writteninformed consent form. For participants less than 18 years of age,informed consent was obtained from their parents. This study ispart of a larger project aiming to improve HAT diagnosis for whichapproval was obtained from WHO (Research Ethics Review Com-mittee) and Institut de Recherche pour le Développement (ComitéConsultatif de Déontologie et d’Ethique) ethical committees.
4.2. Quantification of APOL1 expression
Blood RNA were extracted with the PAXgene� Blood RNA kit(PreAnalytiX), quantified by NanoDrop� spectrophotometer(NanoDrop Technologies, Wilmington, DE) and quality checkedusing the RNA Integrity Number (RIN) on an Agilent 2100 Bioana-

H. Ilboudo et al. / Infection, Genetics and Evolution 12 (2012) 1519–1523 1523
lyzer� with the RNA 600 Nano LabChip (Agilent Technologies).cDNA synthesis was carried out with the High-Capacity cDNA Re-verse transcription kit� (Applied Biosystems) according to themanufacturer’s instructions. Primers for the reference or ‘‘house-keeping’’ gene (G3PDH) and for the target gene (APOL1) were de-signed, using Primer3 software (http://www.frodo.wi.mit.edu/primer3/), to span two exons (G3PDH f-AGATCATCAGCAATGCCTC,G3PDH r-GACTGTGGTCATGAGTCCTTC; APOL1 f-CTGCATCTGGAT-GAGTGC; APOL1 r-AGATACTGCTCTCTGGGTCC) in order to avoidthe amplification of contaminant DNA. We also checked that nopolymorphisms were described at primer loci in public databases.Primers were then commercially synthesized (Eurogentec, France).Briefly, real time quantitative RT-PCR (qRT-PCR) reactions werecarried out in a total volume of 25 ll with theBrillant� SYBR� GreenQPCR Core Reagent kit (Stratagen Corp) with 15 ng of cDNA and 1 llof each primer. Optimal primer concentrations determined for thetwo primer pairs were: 600 nM for each primer (G3PDH) and900 nM for each primer (APOL1). Real time qRT-PCR was performedusing a MX3000P� quantitative PCR system (Stratagen Corp) withthe following cycling parameters: 95 �C for 10 min (PCR hot start),followed by 40 cycles of 95 �C for 30 s, 58 �C for 30 s and 72 �C for45 s, followed by a final melting curve program to assess amplifica-tion specificity.
4.3. Statistical analyses
APOL1 expression levels were calculated by the 2�DDCT method(Livak and Schmittgen, 2001) and results are given as mean foldchanges, relative to the individual with highest APOL1 Ct (used asthe calibrator). As APOL1 expression levels were not normally dis-tributed (p 6 0.5 by the Shapiro–Wilk W test), APOL1 expressionlevels were log transformed prior to carrying linear regressionanalysis. Association of APOL1 expression levels with recordedcovariates (disease focus, age, sex, blood parasite density and infec-tion status) were analysed in univariate or multivariate linearregressions by the standard least square method with the JMP7.0 software (SAS Institute Inc.). p-values <0.05 were consideredsignificant.
Acknowledgments
We would like to thank all study subjects for their kindnesswith the field team and their active participation in the organisa-tion of the field work. We are also very grateful to the Forecariah,Dubreka and Boffa District Health Department (DPS) medicalteams who participated actively in all the field activities organisedwithin this study. Finally we are also grateful to André Garcia forhelpful comments in the process of carrying out statistical analy-ses. This study was supported by the Institut de Recherche pourle Développement (IRD), the French Ministry of Foreign Affairsthrough the FSP/REFS project and the World Health Organization(WHO).
References
Bucheton, B., MacLeod, A., Jamonneau, V., 2011. Human host determinantsinfluencing the outcome of Trypanosoma brucei gambiense infections. ParasiteImmunol. 33, 438–447.
Bullard, W., Kieft, R., Capewell, P., Veitch, N.J., Macleod, A., Hajduk, S.L., 2012.Haptoglobin–hemoglobin receptor independent killing of African trypanosomesby human serum and trypanosome lytic factors. Virulence 3, 72–76.
Buscher, P., Mumba Ngoyi, D., Kabore, J., Lejon, V., Robays, J., Jamonneau, V.,Bebronne, N., Van der Veken, W., Bieler, S., 2009. Improved models of minianion exchange centrifugation technique (mAECT) and modified singlecentrifugation (MSC) for sleeping sickness diagnosis and staging. PLoS Negl.Trop. Dis. 3, e471.
Camara, M., Camara, O., Ilboudo, H., Sakande, H., Kabore, J., N’Dri, L., Jamonneau, V.,Bucheton, B., 2010. Sleeping sickness diagnosis: use of buffy coats improves thesensitivity of the mini anion exchange centrifugation test. Trop. Med. Int. Health15, 796–799.
Capewell, P., Veitch, N.J., Turner, C.M., Raper, J., Berriman, M., Hajduk, S.L., MacLeod,A., 2011. Differences between Trypanosoma brucei gambiense groups 1 and 2 intheir resistance to killing by trypanolytic factor 1. PLoS Negl. Trop. Dis. 5, e1287.
Duchateau, P.N., Movsesyan, I., Yamashita, S., Sakai, N., Hirano, K., Schoenhaus, S.A.,O’Connor-Kearns, P.M., Spencer, S.J., Jaffe, R.B., Redberg, R.F., Ishida, B.Y.,Matsuzawa, Y., Kane, J.P., Malloy, M.J., 2000. Plasma apolipoprotein Lconcentrations correlate with plasma triglycerides and cholesterol levels innormolipidemic, hyperlipidemic, and diabetic subjects. J. Lipid Res. 41, 1231–1236.
Duchateau, P.N., Pullinger, C.R., Cho, M.H., Eng, C., Kane, J.P., 2001. Apolipoprotein Lgene family: tissue-specific expression, splicing, promoter regions; discovery ofa new gene. J. Lipid Res. 42, 620–630.
Genovese, G., Friedman, D.J., Ross, M.D., Lecordier, L., Uzureau, P., Freedman, B.I.,Bowden, D.W., Langefeld, C.D., Oleksyk, T.K., Uscinski Knob, A.L., Bernhardy, A.J.,Hicks, P.J., Nelson, G.W., Vanhollebeke, B., Winkler, C.A., Kopp, J.B., Pays, E.,Pollak, M.R., 2010. Association of trypanolytic ApoL1 variants with kidneydisease in African Americans. Science 329, 841–845.
Huet, G., Lemesre, J.L., Grard, G., Noireau, F., Boutignon, F., Dieu, M.C., Jannin, J.,Degand, P., 1990. Serum lipid and lipoprotein abnormalities in human Africantrypanosomiasis. Trans. R. Soc. Trop. Med. Hyg. 84, 792–794.
Ilboudo, H., Jamonneau, V., Camara, M., Camara, O., Dama, E., Leno, M., Ouendeno, F.,Courtin, F., Sakande, H., Sanon, R., Kabore, J., Coulibaly, B., N’Dri, L., Diarra, A.,N’Goran, E., Bucheton, B., 2011. Diversity of response to Trypanosoma bruceigambiense infections in the Forecariah mangrove focus (Guinea): perspectivesfor a better control of sleeping sickness. Microbes Infect. 13, 943–952.
Jamonneau, V., Bucheton, B., Kabore, J., Ilboudo, H., Camara, O., Courtin, F., Solano, P.,Kaba, D., Kambire, R., Lingue, K., Camara, M., Baelmans, R., Lejon, V., Buscher, P.,2010. Revisiting the immune trypanolysis test to optimise epidemiologicalsurveillance and control of sleeping sickness in West Africa. PLoS Negl. Trop.Dis. 4, e917.
Kabore, J., Macleod, A., Jamonneau, V., Ilboudo, H., Duffy, C., Camara, M., Camara, O.,Belem, A.M., Bucheton, B., De Meeus, T., 2011. Population genetic structure ofGuinea Trypanosoma brucei gambiense isolates according to host factors. Infect.Genet. Evol. 11, 1129–1135.
Kieft, R., Capewell, P., Turner, C.M., Veitch, N.J., MacLeod, A., Hajduk, S., 2010.Mechanism of Trypanosoma brucei gambiense (group 1) resistance to humantrypanosome lytic factor. Proc. Natl. Acad. Sci. USA 107, 16137–16141.
Koffi, M., De Meeus, T., Bucheton, B., Solano, P., Camara, M., Kaba, D., Cuny, G., Ayala,F.J., Jamonneau, V., 2009. Population genetics of Trypanosoma brucei gambiense,the agent of sleeping sickness in Western Africa. Proc. Natl. Acad. Sci. USA 106,209–214.
Lecordier, L., Vanhollebeke, B., Poelvoorde, P., Tebabi, P., Paturiaux-Hanocq, F.,Andris, F., Lins, L., Pays, E., 2009. C-terminal mutants of apolipoprotein L-Iefficiently kill both Trypanosoma brucei brucei and Trypanosoma bruceirhodesiense. PLoS Pathog. 5, e1000685.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2(�Delta Delta C(T)) Method. Methods 25,402–408.
Molina-Portela, M.P., Samanovic, M., Raper, J., 2008. Distinct roles of apolipoproteincomponents within the trypanosome lytic factor complex revealed in a noveltransgenic mouse model. J. Exp. Med. 205, 1721–1728.
Pays, E., Vanhollebeke, B., 2008. Mutual self-defence. the trypanolytic factor story.Microbes Infect. 10, 985–989.
Pays, E., Vanhollebeke, B., Vanhamme, L., Paturiaux-Hanocq, F., Nolan, D.P., Perez-Morga, D., 2006. The trypanolytic factor of human serum. Nat. Rev. Microbiol. 4,477–486.
Perez-Morga, D., Vanhollebeke, B., Paturiaux-Hanocq, F., Nolan, D.P., Lins, L.,Homble, F., Vanhamme, L., Tebabi, P., Pays, A., Poelvoorde, P., Jacquet, A.,Brasseur, R., Pays, E., 2005. Apolipoprotein L-I promotes trypanosome lysis byforming pores in lysosomal membranes. Science 309, 469–472.
Van Meirvenne, N., Magnus, E., Buscher, P., 1995. Evaluation of variant specifictrypanolysis tests for serodiagnosis of human infections with Trypanosomabrucei gambiense. Acta Trop. 60, 189–199.
Vanhamme, L., Paturiaux-Hanocq, F., Poelvoorde, P., Nolan, D.P., Lins, L., Van DenAbbeele, J., Pays, A., Tebabi, P., Van Xong, H., Jacquet, A., Moguilevsky, N., Dieu,M., Kane, J.P., De Baetselier, P., Brasseur, R., Pays, E., 2003. Apolipoprotein L-I isthe trypanosome lytic factor of human serum. Nature 422, 83–87.
Vanhollebeke, B., Pays, E., 2010. The trypanolytic factor of human serum: manyways to enter the parasite, a single way to kill. Mol. Microbiol. 76, 806–814.
Vanhollebeke, B., Truc, P., Poelvoorde, P., Pays, A., Joshi, P.P., Katti, R., Jannin, J.G.,Pays, E., 2006. Human Trypanosoma evansi infection linked to a lack ofapolipoprotein L-I. N. Engl. J. Med. 355, 2752–2756.
Vanhollebeke, B., Lecordier, L., Perez-Morga, D., Amiguet-Vercher, A., Pays, E., 2007.Human serum lyses Trypanosoma brucei by triggering uncontrolled swelling ofthe parasite lysosome. J. Eukaryot. Microbiol. 54, 448–451.
Vanhollebeke, B., De Muylder, G., Nielsen, M.J., Pays, A., Tebabi, P., Dieu, M., Raes, M.,Moestrup, S.K., Pays, E., 2008. A haptoglobin–hemoglobin receptor conveysinnate immunity to Trypanosoma brucei in humans. Science 320, 677–681.





























