Embed Size (px)
Citation preview

Eur. J. Biochem. 168,209-217 (1987) ((3 FEBS 1987
Biosynthesis of Lewis fucolipid antigens in human colorectal carcinoma cells Partial characterization of LcOse4Cer and H-1 fucolipid fucosyl transferase acceptor activities
Vis A. LIEPKANS' and Goran LARSON' ' Equipc de Recherche 277 du Centre National de la Recherche Scientifique, Institut de Recherches Scientifiques sur le Cancer, Villejuif
Department of Clinical Chemistry, Sahlgren's Hospital, University of Goteborg
(Received January 8/March 26, 1987) - EJB 87 0011
Purified glycolipids were tested for their ability to serve as acceptors of ['4C]fucose from GDP-['4C]fucose as catalyzed by cell-free extracts and purified membrane fractions of human colorectal carcinoma cells, SW1116, cultured in serum-free medium. Purified lactotetraosyl ceramide (GalP1+3GlcNAcPl+ 3GalP1+4Glc-Cer or LcOse4Cer) and H-1 glycolipid (Fucal+2Gal~1+3GlcNAc~1+3Gal~1+4Glc-Cer or IV2 FucaLcOse4Cer) stimulated incorporation of radioactivity into lipid-soluble glycolipid at a rate greater than ten times that of Lea glycolipid [Gal~1+3(Fucal+4)GlcNAc~1+3Gal~1+4Glc-Cer or 1114 FucaLcOse4Cer].
The enzymatic activities in crude and purified membrane fractions were optimized for substrate concentra- tions (glycolipid and GDP-fucose), detergent requirement (taurocholate), pH, time and protein. The radioac- tive product of H-1 fucosylation migrated as discrete and distinct bands on high-performance thin-layer chromatograms (HPTLC). Evidence for their identity with Leb fucolipid described previously [Fucal+2Gal~1-+3(Fucal+4)GlcNAc~1-+3Gal~1+4Glc-Cer or II141V2 (Fuca) LcOse4Cer] is presented.
The radioactive product of LcOse4Cer fucosylation was mainly Lea fucolipid as determined by co-migration with authentic Lea fucolipid in three HPTLC systems as native and acetylated derivatives. Our results also indicated a low level of H-1 and Leb glycolipid synthesis from LcOse4Cer.
On the basis of the optima, linearity for time, and enzyme-limiting conditions, we obtained a 12-19-fold purification of the LcOse4Cer and H-1 fucosyl transferase acceptor activities in three peaks of a sucrose gradient. The peak with the highest specific activity (peak 3) was highest in density and in Na', K', ATPase specific activity, although NADH - cytochrome-c reductase and UDP-GalNac transferase were also present in peak 3.
The apparent K , values of LcOse4Cer acceptor activity and H-1 acceptor activity in peak 3 were significantly different 0) < 0.01) by statistical tests, 2.4 pM and 0.5 pM, respectively. These apparent K, values were much lower (lo3 x ) and the pH optima were lower (4.8 - 5.3), than the corresponding properties reported for the a1 +3/ a1 4 4 fucosyl transferase purified from human milk. Our results suggest a role for the non-glycosidic moieties of the acceptors and/or the tissue-specific or primitive expression of these fucosyl transferase activities.
Fucose was first identified as a component of a glycolipid fraction in 1963 by Koscielak [l]. It was first reported to be associated with the antigenicity of red blood cells by Yamakawa et al. [2]. The first pure fucolipid was isolated from human gastric adenocarcinoma by Hakomori and Jeanloz in 1964 [3]. Since the advent of high-performance thin-layer chromatography (HPTLC), high-pressure- liquid chromatog- raphy and monoclonal antibody techniques, the structure of a number of complex fucolipids have been determined. Fucolipids can contain glucose (Glc), galactose (Gal),
Correspondence to V. A. Liepkans, Institut de Recherches Scientifiques sur le Cancer, Boite postale 8, F-94802 Villejuif Ckdex, France
Abbreviations. LcOse4Cer, lactotetraosylceramide (GalP1+3- GlcNAcPl +3GalP1+4Glc-Cer); H-1 glycolipid or fucolipid (Fucal+2GalPl+ 3GlcNAc~1+3Gal/31+4Glc-Cer); Lea glycolipid or fucolipid [Gal~1+3(Fucal+4) GlcNAc~l+3Gal~1-+4Glc-Cer]; Leb glycolipid or fucolipid [Fucal-t2GalP1+3(Fucal+4)- GlcNAc~l-+3Gal~1+4Glc-Cer]; GalNAc, n-acetylgalactosamine; Fuc, fucose (6-deoxy-~-galactose); Cer, ceramide; GgOse4Cer, gan- gliotetraosyl ceramide; GbOse4Cer, globoside; GbOse5Cer, Forssman antigen.
Some of these results were presented in abstract form at the 1986 meetings of the Society for Complex Carbohydrates (Charleston, SC, USA) and the American Society for Cell Biology (Washington, DC, USA).
N-acetylglucosamine (GlcNAc), N-acetylgalactosamine (GalNAc) and N-substituted neuraminic acids. All of the sugars so far reported in fucolipids are in glycosyl linkages in the pyranosyl form and of the D-series except fucose (6-deoxy- L-galactose). Most fucolipids are derivatives of 1acto-N- tetraosyl (LcOse,), lacto-N-neotetraosyl (nLcOse,) ganglio- tetraosyl (GgOse4) ceramide or the globo-series [4].
Fucolipids which carry antigenic determinants have been found associated with human tumor tissues [5], tissues in development [6], and human tumor cells in culture [7]. Promi- nent among these are the Lewis blood group antigens. Lewisa glycolipid [Gal~1+3(Fucal+4)GlcNAc~1~3Gal~1+4Glc- Cer or 1114 Fuca-LcOse4Cer] and Lewisb glycolipid [Fuc- a1 + 2GalP1 + 3(Fucal+4)GlcNAcPl+ 3GalPl+ 4Glc-Cer or II141V2 (F~ca)~LcOse,Cer] have been detected and isolated from canine intestine and human intestinal tumor cells in culture [S, 91.
To our knowledge there have been no biochemical studies on the cell-free biosynthesis of Lewisa and Lewisb glycolipids, using pure glycolipid substrates and human colorectal carcinoma cells. Prohaska et al. in 1978, in an immuno- chemical study presented evidence for the synthesis of Lewisa and Lewisb glycolipids on the surface of erythrocytes (which had adsorbed a glycolipid precursor of presumed structure) in the presence of human mucosal extracts [lo]. Erythrocytes from Lewisa-b- individuals were able to support the enzymatic

210
synthesis of Lea and Leb glycolipids. These authors also suggested that the Lea could be a precursor of Leb glycolipid. Prieels et al. have presented evidence using oligosaccharide substrates that a1 +3 and a1 +4 fucosyl transferase activities from human milk co-purify [l l] . Campbell and Stanley have described two, a1 -3 fucosyl transferase activities in Chinese hamster mutants which differ in glycoprotein acceptor speci- ficity [12]. Hakomori has presented an interesting hypothesis which predicts that 'aberrant' fucosylation of Lea to Leb may occur in relation to oncogenesis [13].
Study of many glycosyl transferases has been hampered by the unavailability of pure acceptor substrates. With the exception of the enzyme purified from human milk, the enzymatic properties of the a1 -4 fucosyl transferases have not been well characterized [14]. Tumor-associated fucosyl transferases which synthesize the Lewis family of fucolipids in colorectal carcinoma cells fall into this category. We have obtained highly purified glycolipid substrates carrying LcOse4, IV2Fuca-LcOse4 and II14Fuca-LcOse4 structures. The purpose of these studies was to test and quantify their ability to accept fucose from GDP-fucose in the presence of cell-free extracts of human intestinal tumor cells SW1116.
The results support the prediction that these intestinal tumor cells can synthesize Lea and Leb glycolipid from LcOse4Cer and H-1 glycolipid (IV2Fuca-LcOse4Cer) sub- strates, but do not support the hypothesis that aberrant fucosylation of Lea glycolipid [I31 occurs to any significant extent in these human intestinal tumor cells. We also present evidence for differences in the enzymatic properties of the LcOse4Cer and H-1 fucosyl transferase acceptor activities.
We found that a significant portion of these 1-4 fucosyl transferase activities was present in a purified membrane frac- tion which was high in Na+/K+-ATPase activity, a plasma membrane marker.
Partial characterization of these LcOse4Cer and H-I fucolipid acceptor activities indicate that their properties are significantly different from those reported for the purified a1 -t3/al-t4 transferase from human milk [l l] , and point to a need for studies with intact acceptors.
MATERIALS AND METHODS
Cell biology and purification of Lewis antigens
SW1116 cells were cultured in serum-free medium and cell pellets were washed and harvested as previously described [15]. Anti-Lewisa and anti-Lewisb monoclonal antibodies were obtained from Chembiomed Ltd (Edmonton, Alberta, Canada). Radioactive Lewis antigens, whether from whole cells or from cell-free incubations, were extracted and purified as previously described with the modification in the final step of Fractosil columns (120 - 130 mesh, Merck, Darmstadt, FRG) with a stepwise gradient of chloroform/methanol (CHC13/CH30H) (4/1, 2/1, 1/1) and methanol. The Lewis antigens were found in the CHC13/CH30H (1/1) fraction and/ or the methanol fraction depending on the column load and dimensions. For glycolipid derived from 500 pg cell protein, a Fractosil column of 0.5 x 5 cm with 25-ml fractions will elute about 90% of the lipid-soluble fucolipid in the methanol and CHC13/CH30H (1/1) fractions.
Assay for lipid-soluble fucolipid synthesis A cell pellet obtained from one confluent T 175-cm2 flask
was suspended in 1 ml cacodylate/sucrose/MgC12 (0.7 ml
0.2 M sodium cacodylate pH 5.4, 0.3 ml 2 M sucrose, 70 pl 0.05 M MgC12). After 100 strokes in a Wheaton B dounce homogenizer, the preparation was centrifuged at 500 x g for 30 min and the resulting supernatant was centrifuged again at 2000 x g for 30 min. This supernatant was free of whole cells and was the cell extract used in this study; it contained 3 - 4 pg protein/pl. It could be kept up to 6 months at - 30 "C with little or no loss in fucosyl transferase activities. Sucrose- density-purified membranes were stored at - 80°C.
Purification of the fucosyl transferase activities by sucrose density centrifugation was a procedure modified from Schachter et al. [16] and Knope et al. [17]. Cell pellets from six T 175-cm2 culture flasks (z 50000 cells/cmz) were suspended in buffer A (20% sucrose, 0.2 M cacodylate, 5 mM 2-mercaptoethanol, 1% dextran, pH 5.3) and sonicated on ice (medium setting, 15-s sonication, 15-s stop, repeat four time). The resulting suspension was then centrifuged at 1000 x g for 15 min. The supernatant was centrifuged at I00000 x g for 60 min. The resulting cell pellet was suspended in 5 ml buffer A pH 5.3 and applied to a discontinuous sucrose gradient: 5ml each of 25% sucrose, 30% sucrose, 35% sucrose, 40% sucrose, 45% sucrose and 55% sucrose (all buffered with Mg" cacodylate, pH 5.3) for 20 h in an SW 27 rotor at 27000 rpm. We recovered the fractions by sterile pipetting from the top, carefully avoiding cross-contamination; 1 -ml fractions were collected and every other tube was assayed for fucosyl transferase activities. The four or five tubes from each peak of activity were pooled and designated peaks 1, 2 and 3 according to their density (see Results). This material was regurgitated through a small-bore pipette until an even dis- tribution of the suspension was attained and assayed for pro- tein according to Bradford [18] in the presence and absence of detergent (taurocholate).
Na+/K+-ATPase and Mg + -ATPase specific activities were determined according to Solomonson et al. [I91 in the presence of taurocholate (7 mM). NADH - cytochrome-c reductase was assayed with pure NADH and beef heart cytochrome c (Sigma) according to Solomonson and Vennesland with E of 21 mM-' cm-' and in the presence of antimycin A [20]. UDP-GalNAc transferase was assayed essentially according to Fitzgerald et al. [21] and Coste et al. [22] as for galactose transferase except that UDP-[ 14C]GalNAc (NEC-592, specific activity 47.2 Ci/mol) was used and purified human intestinal much was the acceptor. Intestinal mucin was a gift of Dr J. Nardelli (University of California at San Francisco, V. A. Med. Ctr., Gastrointestinal Unit). It was filtered through Nalgene type S (0.2 pm) filters, dialyzed against sterile HzO and lyophilized prior to use, the final concentration being 10 pg/ml. We modified the assay to a scale of 2 - 5 pg protein in a total volume of 150 p1 and washed the phosphotungstic acid precipitates three times with 3 ml cold 0.5 M trichloroacetic acid and three times with 3 ml ethanol at - 20°C prior to counting.
Each fucosyl transferase assay incubation contained 100 - 250 pg sodium taurocholate (detergent), 5 - 10 pg purified glycolipid acceptor, 100 pmol cacodylate, 0.25 pmol MgC12 and 0.5- 1.0 nmol GDP-['4C]fucose (254 Ci/mol, New England Nuclear, Boston, MA, USA) and 2 - 23 pg cell-free protein in a total volume of 70 pl. The detergent in CHC13/ CH30H (2/1) and the glycolipid substrates (LcOse4Cer, H-I glycolipid or Lea glycolipid) in CHC13/CH30H (1/2), 3% H 2 0 were added first and the solvents evaporated prior to the addi- tion of the other components. The GDP-['4C]fucose, which was in H20/ethanol (l / l) , was dried to half volume or less to help eliminate EtOH and was added finally in no more than

21 1
5 p1. LcOse4Cer and Lea glycolipid had to be solubilized by alternate heating and vortex at 50°C before addition to the incubation tube. The incubations were run up to 60 min at 37 "C in conical assay tubes and terminated by the addition of 500 p1 CHC13/CH30H (1/2), 3% HzO. After 24-48 h, the total extract, excluding the precipitated protein, was chroma- tographed on paper (3MM Whatman) and 0.5 x 2.0 cm Florisil columns as previously described [23, 241. Glycolipids were isolated from the acetone/dichloroethane fractions of the Florisil columns. Samples were stored at f4"C under anhydrous conditions and were re-acetylated or de-acetylated prior to analysis by HPTLC. When four identical incubations in two separate experiments were performed, standard error was f 6 % .
The GDP-['4C]fucose substrate was stable for the I-h duration of the incubations as verified by paper chromatog- raphy of the water-soluble extracts of the incubations at pH 4.9 - 7.0. When the upper phases of the Folch partitions were chromatographed in ethanol/l M ammonium acetate (7/3) pH 4.0, 96-97% of the 14C radioactivity migrated with untreated GDP-['4C]fucose (NEN) and not with previously degraded I4C-labeled material or [3H]fucose (NEN, Boston, MA, USA).
Radiolabeling of LcOse4Cer glycosphingolipid
Radiolabeling of LcOse4 was performed according to a method modified from Leskawa et al. [25]. About 50 pg purified LcOse4Cer was dried down in a conical test tube with 100 pg sodium taurocholate. To this mixture was added 1 mg (45 units) galactose oxidase (from Polyporus circinatus, P-L Biochemicals, Milwaukee) in 50 pl 0.1 M sodium phosphate buffer pH 7.0. This suspension was incubated 8 h, then another 1 mg galactose oxidase was added and incubation continued overnight. An excess of CHC13/CH30H/H20 (120/ 30/5) was added and the material put through an SiOz (0.5 x 2.0 cm) column with three 5-ml washes. The dried lipid collected from the column was dissolved (with gentle warming) in 100 pl tetrahydrofuran/HzO (4/1) and a 100-fold molar excess of NaB[3H]4 (10 Ci/mmol) was added. This was incubated for 12 h at room temperature, another 100-fold molar excess of NaB[3H]4 added and incubated for another 12 h. The material was partitioned in 4 ml CHC13/CH30H (2/1) plus 1 ml 0.9% NaCl, three times and the lower phases were combined dried and analyzed by HPTLC. We obtained a [3H]LcOse4Cer preparation which was of 80% isotopic purity by HPTLC in two systems (CHC13/CH30H/H20, 60/ 35/8 and 60/30/5). Almost all of the isotopic impurity (95%) ran faster than standard LcOse4Cer and disappeared after Florisil column chromatography. We estimated the specific activity of this material to be about lo5 cpm/pg.
High-performance thin layer chromatography and glycolipid references
High-performance thin-layer chromatography (HPTLC) was performed to separate and characterize fucolipid and glycolipid standards. HPTLC solvent systems that we used were CHC13/CH30H/H20 (60/35/8 and 60/30/5) for native glycolipids and CHC13/CH30H (94/6, 95/5 and 97/3) for acetylated derivatives. Forssman antigen (GbOse5Cer), Lea glycolipid, H-1 glycolipid, and LcOse4Cer were located by orcinol spray (7 pg each). Lea and Leb glycolipid were also located by an immunobinding assay modified from Magnani
et al. [26]. The RF values of the native glycolipids on silica gel 60 HPTLC aluminium plates (20 x 20 cm; Merck, Darmstadt, FRG) in the solvent system CHC13/CH30H/H20 (60/30/5) for 2.5 h in an unlined tank were GbOse4Cer, 0.80; LcOse4Cer, 0.71 ; GbOse5Cer, 0.61; H-1 glycolipid, 0.50; Lea glycolipid, 0.40 and Leb, 0.20 - 0.30 (doublet).
Lactotetraosyl ceramide (LcOse4Cer) and Lea glycolipid (II14Fuca-LcOse4Cer) were prepared from pooled blood group A meconia, H-I glycolipid (IV'Fuca-LcOse4Cer) and Leb-glycolipid [II14,1V2(Fuca)2-LcOse4Cer] from pooled blood group 0 meconia as previously reported [8, 27, 281. After obtaining non-acid glycolipid fractions, the glycolipids were further separated as acetylated derivatives using, initial- ly, Li Chro Prep Si 60 (1 5 - 25 pm, Merck, Darmstadt, FRG) and a stepwise elution with increasing concentrations of meth- anol in chloroform. Final purifications were obtained using Iatrobeads 6RS-8060, Silica gel (Iatron Labs. Tokyo, Japan) and gradient elutions with 0-2% and 2-6% methanol in chloroform for acetylated derivatives and CHC13/MeOH/ HzO mixtures (65/25/4,60/35/8,50/40/10,40/40/12, v/v/v) for native glycolipids. LcOse4Cer was extensively purified as the acetylated derivative also using increasing amounts of tert- butanol or methyl acetate in dichloromethane as eluants. Glycolipid preparations were analyzed by mass spectrometry as permethylated derivatives using a ZAB-HF mass spec- trometer (VG analytical, Wythenshire, England). Globoside was obtained from Supelco (Bellefonte, PA) and Forssman antigen was kindly donated by Dr J. Portoukalian (Centre Lkon Bkrard, Lyon, France).
Analyzes of the I4C-labeled product of cell-free incubations
Material from pooled assays was deacetylated after Florisil chromatography. About 2000 cpm, representing three or four incubations, was dried down and solubilized in 200 pl CHC13/CH30H (2/1), 50 pl of 0.5% Na'OCH; in CH30H was added and the mixture incubated at 25°C for 45 min. After addition of 10% acetic acid to neutralize, Fractosil chromatography was performed as above and the fractions containing radioactivity were chromatographed by HPTLC as above. After drying we sprayed the lanes supporting 14C- labeled cell-free products with enhancer spray (New England Nuclear, Boston, MA, USA) so as to perform fluorography for 7- 10 days at -80°C; 300- 500 dpm could be easily visualized as discrete bands by these methods.
Adjoining lanes which supported the migration of authentic standards were cut separately (aluminium HPTLC plates, Silica 60, Merck) and sprayed with orcinol/HzS04 spray. Adjoining lanes which were spotted with Lea and SW1116 extracts (containing Leb antigen) were cut and analyzed by immunobinding assays [25,26]. Another lane was spotted with lipid-soluble glycolipid obtained from whole cells which had been incubated 3 days in serum-free medium con- taining [3H]fucose [15].
Aliquots of the enzyme products were degraded in 0.5 M trichloroacetic acid for 2 h at 90°C and the degradation prod- ucts analyzed by paper chromatography in butanol/ethanol/ H 2 0 (10/1/2) for 70 h [29] for separation of ['4C]fucose from galactose and N-acetylglucosamine; 90 - 95% of the radioac- tivity migrated with the fucose band on paper chromatog- raphy. Sugars (Sigma, St Louis, USA) were located by dipping in AgN03/ethanol [30]. The 14C-labeled glycolipid products were treated with beef epididymis fucosidase (Sigma, St Louis, MO, USA) to test for lability; about 2.5 pg glycolipid contain- ing 450 dpm was treated with 0.2 unit enzyme preparation in

212
0.1 M citrate, 0.01% NaN3, pH 6.2 in a total volume of 250 p1 for 24 h at 37°C [30].
RESULTS
The migration on HPTLC of the native glycolipids which we used in this study as acceptors for fucosylation and as reference standards is shown in Fig. 1. Mass spectrometric data of the different fractions confirmed the LcOse4, H-1, Lea and Leb carbohydrate sequences and established the dominating ceramides of all fractions as phytosphingosine with hydroxy 24:O or 22:O (N-acylatcd) fatty acyl groups.
Table 1 shows a comparison of fucosyl transferase specific activities for three possible acceptors in a crude membrane extract and two peaks from a sucrose density centrifugation as described in Methods. It is a summary of a series of re- produced experiments to determine the extent of apparent purification obtained. In practice, maximal rates of acceptor activity within 1 h of incubation giving a useful amount of radioactivity can be obtained with concentrations of 25 - 55 pM glycolipid acceptor, 7 - 12 pM GDP-fucose and 3 - 7 mM taurocholate. Addition of higher concentrations of these
1 2 3 4 Fig. 1. High-performance thin-layer chromatography of the glycolipid substrates used in this study. Lane 1, LcOseCer; lane 2, H-1 fucolipid; lane 3, Lea fucolipid; lane 4, Leb fucolipid reference standard. Solvent system: chloroform/methanol/water (60/35/8) on Merck Si60 HPTLC plates in sandwich chamber
components did not significantly enhance incorporation of I4C into glycosphingolipid. We used 17-23 pg protein for crude membranes and 2- 5 pg protein for purified mem- branes (per 70-p1 incubation). Saturating levels of GDP- fucose substrate were determined from substrate dependency curves (not shown) and the significant presence of GDP- fucose pyrophosphatase activity was excluded by chromatog- raphy of the water-soluble extracts of the incubations (see Methods). The purified Lewis" glycolipid was not a good acceptor for [14C]fucose in the range of parameters that we tried and is therefore a good control for the effect of carrier in the series of chromatographic steps in the assay. There was no absolute dependency on Mg2+ ion but in initial studies with crude membranes some stimulation was observed (10- 40%) so we have chosen to retain MgZf under standard conditions of assay throughout purification. Table 1 demonstrates the conditions for linearity with respect to time and protein and with saturation of glycolipid acceptor and GDP-['4C]fucose; however, we also found that an over-abun- dant concentration of glycosphingolipid ( 2 100 pM) tended to alter linearity, especially with LcOse4Cer. All samples were measured with respect to incubations containing no glycolipid (which were in turn measured with respect to boiled enzyme controls) to determine the activity dependent on exogenous purified glycosphingolipid; generally endogenous activity was very low (in the range of not more than twice backround for 2 pg protein). Radioactivity in each sample was determined for 30 min in a programmed scintillation counter (LKB beta- rack) which automatically subtracted control values and endogenous acceptor activity from each incubation and had a visual display of the window spectra including chemilumi- nescence.
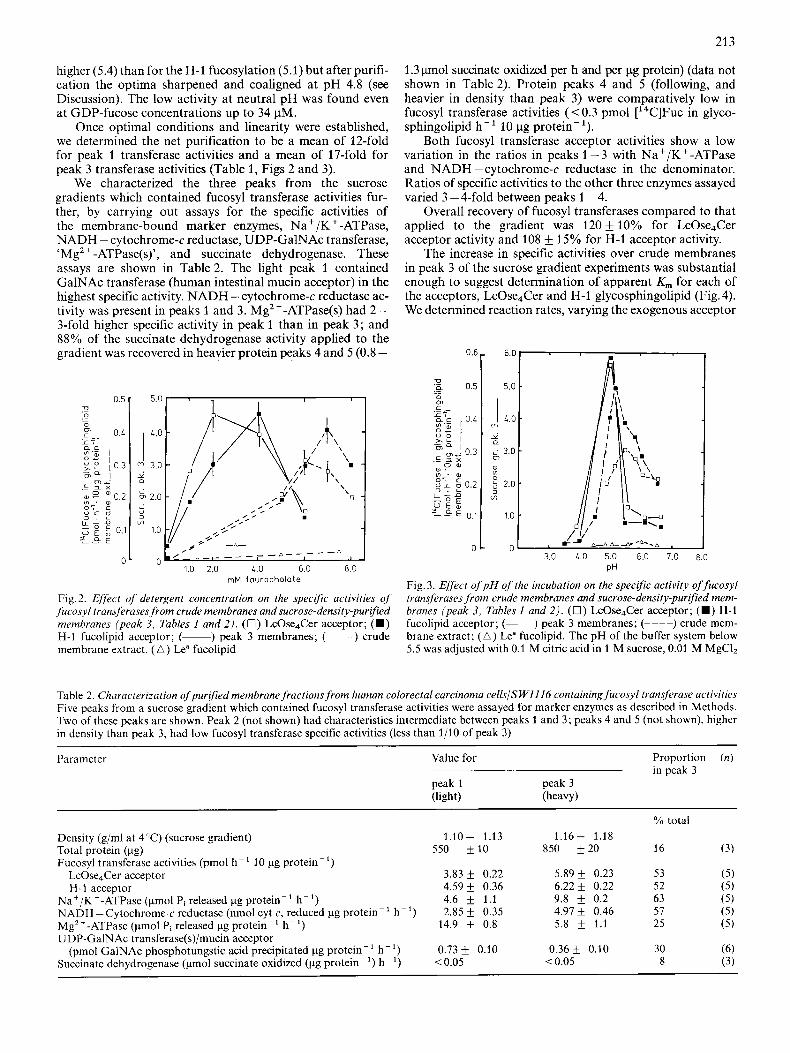
Overall, the optimal requirements for taurocholate detergent were lower (2-4 mM) in peak 3 than in the crude membrane extracts (6 - 7.5 mM) (Fig. 2). H-1 fucolipid had a requirement for more detergent in order to obtain maximal rates than LcOse4Cer in the crude membranes and in the purified membranes in terms of the mean but we were unable to achieve statistical significance ( P > 0.05) for this relation- ship.
Determination of the pH optima is shown in Fig. 3. In the crude extract the optimum for LcOse4Cer fucosylation was
Table 1. Fucosyl transferase glycolipid acceptor activities from human colorectal carcinoma cells ( S W1116) Data is presented as [ ''C]fucose in glycolipids isolated as described in Methods, except that incubations with crude membranes contained 17-23 pg protein, 6.5 mM taurocholate at pH 5.3; incubations with purified membranes contained 2-4 pg protein, 3.0 mM taurocholate at pH 4.8 (total volume of all incubations was 70 pl). Control values have a maximum standard error of ,6%. n.d. = not determined
System Crude membrane extract Sucrose gradient purified membranes
LcOse4Cer H-1 peak 1 peak 3
LcOse4Cer H-I LcOse4Cer H-I
Complete (control) Half glycosphingolipid Half protein +Mn2+ Double time (2 h) Double GDP-Fuc
- LcOse4Cer, - H-1 + Lea No glycolipid
~
pmoI ['4C]fucose hK' (10 pg protein)-'
0.32 0.43 0.20 0.32 0.12 0.21 n.d. n.d. 0.60 0.84 0.35 0.38
0.03 0 (blank ref.)
3.96 4.87 6.18 6.25 3.66 3.69 5.87 5.97 2.20 2.34 2.35 3.27 4.49 4.36 4.46 5.45 6.67 11.50 11.18 12.33 4.15 3.96 6.14 6.91
0.09 0
213
higher (5.4) than for the H-1 fucosylation (5.1) but after purifi- cation the optima sharpened and coaligned at pH 4.8 (see Discussion). The low activity at neutral pH was found even at GDP-fucose concentrations up to 34 pM.
Once optimal conditions and linearity were established, we determined the net purification to be a mean of 12-fold for peak 1 transferase activities and a mean of 17-fold for peak 3 transferase activities (Table 1, Figs 2 and 3).
We characterized the three peaks from the sucrose gradients which contained fucosyl transferase activities fur- ther, by carrying out assays for the specific activities of the membrane-bound marker enzymes, Na+/K+-ATPase, NADH - cytochrome-c reductase, UDP-GalNAc transferase, 'Mg2 + -ATPase(s)', and succinate dehydrogenase. These assays are shown in Table 2. The light peak 1 contained GalNAc transferase (human intestinal mucin acceptor) in the highest specific activity. NADH - cytochrome-c reductase ac- tivity was present in peaks 1 and 3. Mg2+-ATPase(s) had 2- 3-fold higher specific activity in peak 1 than in peak 3; and 88% of the succinate dehydrogenase activity applied to the gradient was recovered in heavier protein peaks 4 and 5 (0.8 -
0.5 r 5.0 I I , I I
1.0 2.0 1.0 6.0 8.0 mM tourocholote
Fig. 2. Effect of detergent concentration on the specific activities of fucosyl transferases from crude membranes and sucrose-density-purqied membranes (peak 3, Tables 1 and 2 ) . (0 ) LcOse4Cer acceptor; ( W ) H-1 fucolipid acceptor; (-) peak 3 membranes; (----) crude membrane extract. ( A ) Lea fucolipid
1.3 pmol succinate oxidized per h and per pg protein) (data not shown in Table 2). Protein peaks 4 and 5 (following, and heavier in density than peak 3) were comparatively low in fucosyl transferase activities (< 0.3 pmol [14C]Fuc in glyco- sphingolipid h-' 10 pg protein-').
Both fucosyl transferase acceptor activities show a low variation in the ratios in peaks 1-3 with Na+/K+-ATPase and NADH - cytochrome-c reductase in the denominator. Ratios of specific activities to the other three enzymes assayed varied 3 - 4-fold between peaks 1 - 4.
Overall recovery of fucosyl transferases compared to that applied to the gradient was 120 f 10% for LcOse4Cer acceptor activity and 108 k 15% for H-1 acceptor activity.
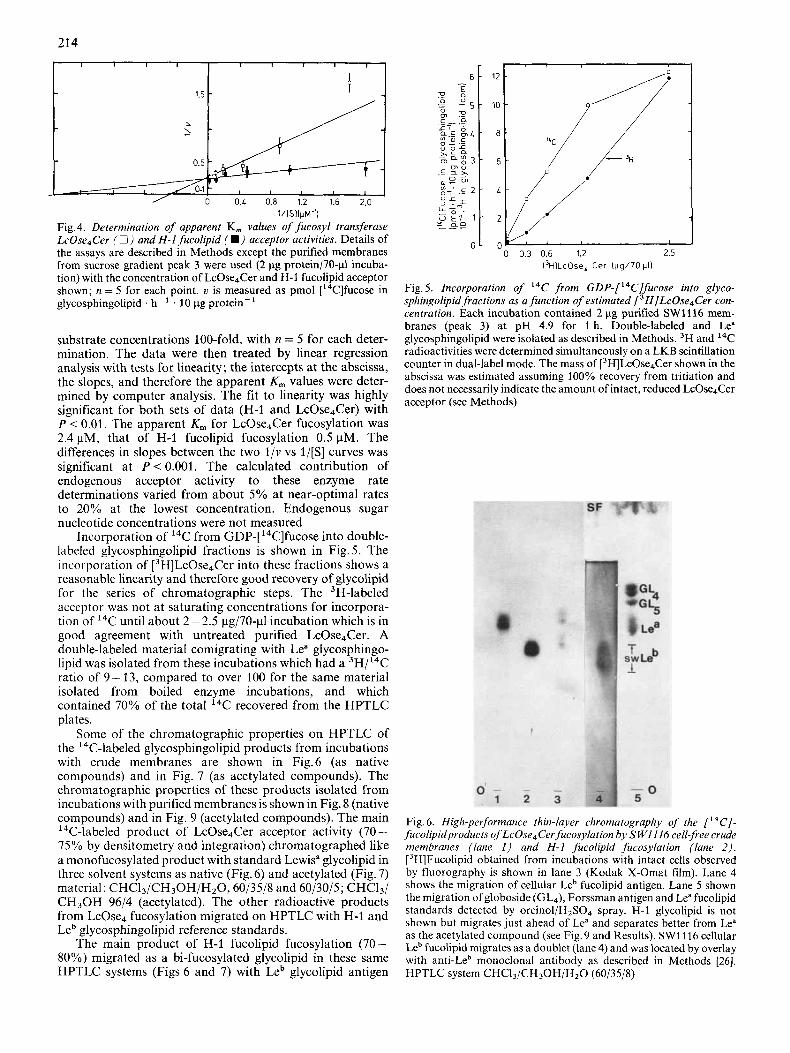
The increase in specific activities over crude membranes in peak 3 of the sucrose gradient experiments was substantial enough to suggest determination of apparent K , for each of the acceptors, LcOse4Cer and H-1 glycosphingolipid (Fig.4). We determined reaction rates, varying the exogenous acceptor
0.6,- 6.0 I I
/A' , t A - A A - d . - A , - A , 1 3.0 L.0 5.0 6.0 7.0 8.0
0
PH
Fig. 3. Effect of p H of the incubation on the specific activity of fucosyl transferases from crude membranes and sucrose-density-purqied mem- branes (peak 3, Tables 1 and 2) . ( 0 ) LcOse4Cer acceptor; (m) H-1 fucolipid acceptor; (-) peak 3 membranes; (----) crude mem- brane extract; ( A ) Lea fucolipid. The pH of the buffer system below 5.5 was adjusted with 0.1 M citric acid in 1 M sucrose, 0.01 M MgClz
Table 2. Characterization of purified membrane fractions from human colorectal carcinoma cells/S WI 116 containing fucosyl transferase activities Five peaks from a sucrose gradient which contained fucosyl transferase activities were assayed for marker enzymes as described in Methods. Two of these peaks are shown. Peak 2 (not shown) had characteristics intermediate between peaks 1 and 3; peaks 4 and 5 (not shown), higher in density than peak 3, had low fucosyl transferase specific activities (less than 1/10 of peak 3)
Parameter Value for Proportion (n ) in peak 3
peak 1 peak 3 (light) (heavy)
YO total Density (g/ml at 4°C) (sucrose gradient) Total protein (pg) Fucosyl transferase activities (pmol h- ' 10 pg protein-')
LcOse4Cer acceptor H-l acceptor
Na+/K'-ATPase (pmol Pi released pg protein-' h-') NADH - Cytochrome-c reductase (nmol cyt c, reduced pg protein-' h- ') Mg*+-ATPase (pmol Pi released pg protein- h- ') IJDP-GalNAc transferase(s)/mucin acceptor
(pmol GalNAc phosphotungstic acid precipitated pg protein- ' h- ') Succinate dehydrogenase (pmol succinate oxidized (pg protein- ') h- ')
1.10- 1.13 550 k10
3.83 & 0.22 4.59 f 0.36 4.6 f 1.1 2.85 f 0.35
14.9 f 0.8
0.73 f 0.10 < 0.05
1.16- 1.18 850 +20
5.89 f 0.23 6.22 f 0.22 9.8 f 0.2 4.97 f 0.46 5.8 1.1
0.36 & 0.10 < 0.05
16
53 52 63 57 25
30 8
21 4
l/lSllpM-’) Fig. 4. Determination of apparent K, values of fucosyl transferase LcOse4Cer (0) and H-1 fucolipid ( W ) acceptor activities. Details of the assays are described in Methods except the purified membranes from sucrose gradient peak 3 were used (2 pg protein/7O-pl incuba- tion) with the concentration of LcOse,Cer and H-1 fucolipid acceptor shown; n = 5 for each point. u is measured as pmol [14C]fucose in glycosphingolipid . h- . 10 pg protein-
substrate concentrations 100-fold, with n = 5 for each deter- mination. The data were then treated by linear regression analysis with tests for linearity; the intercepts at the abscissa, the slopes, and therefore the apparent K, values were deter- mined by computer analysis. The fit to linearity was highly significant for both sets of data (H-1 and LcOse4Cer) with P < 0.01. The apparent K,,, for LcOse4Cer fucosylation was 2.4 pM, that of H-1 fucolipid fucosylation 0.5 pM. The differences in slopes between the two l / v vs l/[S] curves was significant at P < 0.001. The calculated contribution of endogenous acceptor activity to these enzyme rate determinations varied from about 5% at near-optimal rates to 20% at the lowest concentration. Endogenous sugar nucleotide concentrations were not measured
Incorporation of 14C from GDP-[14C]fucose into double- labeled glycosphingolipid fractions is shown in Fig. 5. The incorporation of [3H]LcOse4Cer into these fractions shows a reasonable linearity and therefore good recovery of glycolipid for the series of chromatographic steps. The 3H-labeled acceptor was not at saturating concentrations for incorpora- tion of 14C until about 2 - 2.5 pg/70-p1 incubation which is in good agreement with untreated purified LcOse4Cer. A double-labeled material comigrating with Lea glycosphingo- lipid was isolated from these incubations which had a 3H/14C ratio of 9-13, compared to over 100 for the same material isolated from boiled enzyme incubations, and which contained 70% of the total 14C recovered from the HPTLC plates.
Some of the chromatographic properties on HPTLC of the 14C-labeled glycosphingolipid products from incubations with crude membranes are shown in Fig.6 (as native compounds) and in Fig. 7 (as acetylated compounds). The chromatographic properties of these products isolated from incubations with purified membranes is shown in Fig. 8 (native compounds) and in Fig. 9 (acetylated compounds). The main l4C-labeled product of LcOse4Cer acceptor activity (70 - 75% by densitometry and integration) chromatographed like a monofucosylated product with standard Lewisa glycolipid in three solvent systems as native (Fig.6) and acetylated (Fig. 7) material: CHCI3/CH30H/H20, 60/35/8 and 60/30/5; CHC13/ CH30H 96/4 (acetylated). The other radioactive products from LcOse4 fucosylation migrated on HPTLC with H-1 and Leb glycosphingolipid reference standards.
The main product of H-1 fucolipid fucosylation (70- SOYO) migrated as a bi-fucosylated glycolipid in these same HPTLC systems (Figs 6 and 7) with Leb glycolipid antigen
I3H1LcOse, Cer 1pg/70 p l l
Fig. 5. Incorporation of I4C from GDP-[‘4C]fucose into glyco- sphingolipid fractions as a function of estimated [3H]LcOse4Cer con- centration. Each incubation contained 2 pg purified SW1116 mem- branes (peak 3) at pH 4.9 for 1 h. Double-labeled and Lea glycosphingolipid were isolated as described in Methods. 3H and I4C radioactivities were determined simultaneously on a LKB scintillation counter in dual-label mode. The mass of [3H]LcOse4Cer shown in the abscissa was estimated assuming 100% recovery from tritiation and does not necessarily indicate the amount of intact, reduced LcOse4Cer acceptor (see Methods)
Fig. 6. High-performance thin-layer chromatography of the [ ‘“Cl - fucolipidproducts of LcOse4Cer fucosylation by S WI I16 cell-free crude membranes (lane 1 ) and H-I fucolipid fucosylation (lane 2 ) . [3H]Fucolipid obtained from incubations with intact cells observed by fluorography is shown in lane 3 (Kodak X-Omat film). Lane 4 shows the migration of cellular Leb fucolipid antigen. Lane 5 shown the migration of globoside (GL,), Forssman antigen and Lea fucolipid standards detected by orcinol/H2S04 spray. H-1 glycolipid is not shown but migrates just ahead of Lea and separates better from Lea as the acetylated compound (see Fig.9 and Results). SW1116 cellular Leb fucolipid migrates as a doublet (lane 4) and was located by overlay with anti-Leb monoclonal antibody as described in Methods [26]. HPTLC system CHCI3/CH30H/H,O (60/35/8)
21 5
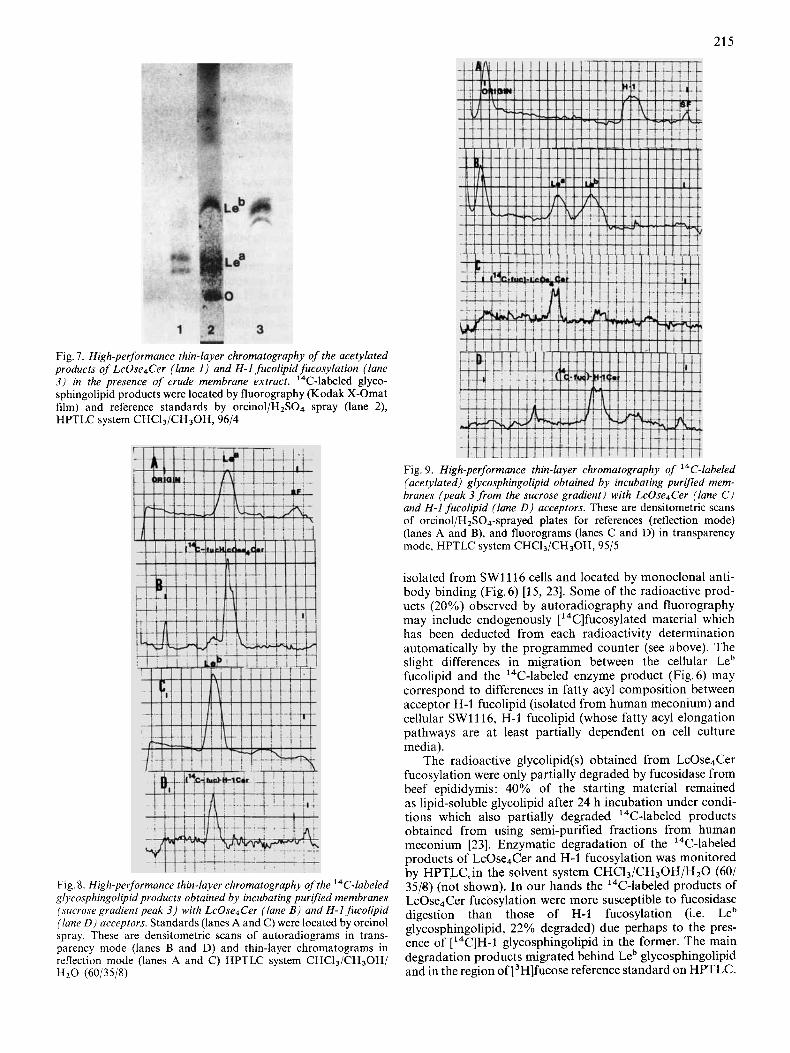
Fig. I . High-performance thin-layer chromatography of the acetylated products of LcOse4Cer (lane 1 ) and H-1 fucolipid fucosylation (lane 3) in the presence of crude membrane extract. 14C-labeled glyco- sphingolipid products were located by fluorography (Kodak X-Omat film) and reference standards by orcinol/HzS04 spray (lane 2), HPTLC system CHCI3/CH30H, 96/4
Fig 8 High-performance thin-layer chromatography of the I4C-labeled glycosphingolipid products obtained by incubating purlfed membranes (sucrose gradient peak 3) with LcOse4Cer (lane B ) and H-1 fucolipid (lane D ) acceptors Standards (lanes A and C) were located by orcinol spray These are densitometric scans of autoradiograms in trans- parency mode (lanes B and D) and thin-layer chromatograms in reflection mode (lanes A and C ) HPTLC system CHC13/CH30H/ HzO (60/35/8)
Fig. 9. High-performance thin-layer chromatography of 14C-labeled (acetylated) glycosphingolipid obtained by incubating purified mem- branes (peak 3 from the sucrose gradient) with LcOse4Cer (lane C ) and H-1 fucolipid (lane D ) acceptors. These are densitometric scans of orcinol/H2SO4-sprayed plates for references (reflection mode) (lanes A and B), and fluorograms (lanes C and D) in transparency mode, HPTLC system CHCI3/CH30H, 95/5
isolated from SW1116 cells and located by monoclonal anti- body binding (Fig.6) [15, 231. Some of the radioactive prod- ucts (20%) observed by autoradiography and fluorography may include endogenously [ ''C]fucosylated material which has been deducted from each radioactivity determination automatically by the programmed counter (see above). The slight differences in migration between the cellular Leb fucolipid and the 14C-labeled enzyme product (Fig. 6) may correspond to differences in fatty acyl composition between acceptor H-1 fucolipid (isolated from human meconium) and cellular SWlll6, H-1 fucolipid (whose fatty acyl elongation pathways are at least partially dependent on cell culture media).
The radioactive glycolipid(s) obtained from LcOse4Cer fucosylation were only partially degraded by fucosidase from beef epididymis: 40% of the starting material remained as lipid-soluble glycolipid after 24 h incubation under condi- tions which also partially degraded 14C-labeled products obtained from using semi-purified fractions from human meconium [23]. Enzymatic degradation of the 14C-labeled products of LcOse4Cer and H-1 fucosylation was monitored by HPTLCiin the solvent system CHCl3/CH30H/Hz0 (60/ 35/8) (not shown). In our hands the 14C-labeled products of LcOse4Cer fucosylation were more susceptible to fucosidase digestion than those of H-1 fucosylation (i.e. Leb glycosphingolipid, 22% degraded) due perhaps to the pres- ence of [14C]H-1 glycosphingolipid in the former. The main degradation products migrated behind Leb glycosphingolipid and in the region of [3H]fucose reference standard on HPTLC.

216
Total acid degradation and paper chromatography showed that essentially all of the radioactivity in these l4C-labe1ed cell-free products could be accounted for by [ ''C]fucose.
DISCUSSION
Alterations of fucolipid antigens in relation to oncogenesis has been the object of increasingly intense study by bio- chemists, immunochemists and cancer researchers since the detection and isolation of unusual structures, or quantita- tively, the unusual presence of blood group antigens in some tumor cells and tissues [5 - 71.
Conclusions from studies on the biosynthesis of complex fucolipids are dependent on the purity of acceptor substrates. Advances in the characterization of the fucosyl transferases were limited by the availability of these acceptors until the advent of synthetic oligosaccharides [31] and pure glycolipid acceptors [27, 281 from human sources. We have chosen to work with intact glycolipids because they are the actual sub- strates in situ; oligosaccharides and complex glycolipids differ predictably in a sufficient number of physical and metabolic properties (compartmentalization, lateral mobility and other membrane-dependent structure/function relationships) to make such data interesting and useful.
The human intestinal tumor cells, SW1116, are interesting because when cultured in totally synthetic medium, they are immunohistochemically positive for Lea, Leb and sialylated Lea antigens [7, 8, 15, 321. In addition, purified glycolipid extracts of these cells present the same antigenicities on HPTLC immunobinding assays [15, 23, 331.
Whole cells in culture incorporate [3HJfucose into fucolipid fractions [15] (Fig.6). Some of this material co- migrates with standard Lewisa and Lewisb fucolipids on HPTLC but we do not exclude the presence of fucolipids in SW1116 cells with type-2 chains.
We conclude from our results that cell-free extracts of SW1116 cells are able to support the fucosylation of purified H-I glycolipid (IV2Fuca-LcOse4Cer) to Leb glycolipid [II141V2(Fuca)2LcOse4Cer].
These cells have been shown to contain a Leb-positive lipid-soluble fucolipid whose structure has been reported [8]. We conclude that it is this compound which comigrated on HPTLC immunobinding assays with the 14C-labeled enzymatic product, which in turn migrated behind three authentic five-sugar glycolipids, and with authentic Leb glycolipid from human meconium as native and acetylated structures (Figs 6 and 7).
The above results and those from epididymis fucosidase and total acid degradation are consistent with the presence of a non-terminal 1 -4-['4C]fucosyl moiety in the ',C-labeled glycosphingolipid products of the incubations.
Furthermore, we conclude that the SW1116 cell-free ex- tracts fucosylate LcOse,Cer to Lea fucolipid (II14Fuca- LcOse4Cer), and to H-1 fucolipid (IVZFuca-LcOse4Cer) which is formed in smaller amounts (10-20%) and at a lower rate by the crude membrane extract and the purified membranes. The monofucosylated products were also identified chromatographically (with reference compounds) in three solvent systems both as acetylated and native structures and by fucosidase and acid degradations. The presence of the LcOse4Cer core structure in the 14C-labeled product was also indicated by isolation of double-labeled glycosphingolipid fractions after incubation of [3H]LcOse4Cer preparations with GDP-['4C]fucose in the presence of purified membranes (Fig. 5). The Lea fucolipid
was not a good substrate for further ['4C]fucosylation under the conditions of our assay; the molar excess (> 100-fold) of taurocholate over glycolipid substrates assured an effective dissolution of glycolipids in the incubation media.
In a previous study we showed that semi-purified glycolipid samples from human meconium contained good acceptors for fucosylation [23]. The fraction termed, 'tetragIycosy1'-containing, behaved like LcOse,Cer by prece- dent and partial characterization. Longer incubation times favored the biosynthesis of slower migrating (HPTLC) glycolipid, probably Leb.
The results in Table 1 and Figs 1 and 2 show that we are comparing maximal rates of fucosyl 1 44-transferase activities under the conditions of incubation.
We were unable to reproduce the differences in pH maxima of LcOse4Cer and H-1 acceptor activities in the purified mem- branes (Fig. 3). But a shift in pH maxima is not unexpected considering that the membranes have shed relatively 90 - 95% of their protein, and probably a substantial amount of lipid, during manipulation and purification. These pH maxima of 4.8 - 5.3 are significantly lower than that reported by Prieels et al. [ l l ] for the a1 +3/al +Cfucosyl transferase from human milk (pH optima 7.0 - 7.8).
Despite a 12 - 19-fold purification, the detergent curves showed similar differences and dependencies for LcOse,Cer and H-1 acceptor activities in crude membranes and purified membranes (peak 3); it remains to be seen whether these data constitute evidence for a difference in the microenvironment (sub-compartmentalization) of the fucosyl transferases. How- ever, the 4.6-fold difference in apparent K,,, (Fig. 4) observed at optimal or near-optimal conditions (Table 1, Figs 2 and 3) are a strong indication that not only is the 1 +Cfucosyl moiety recognized (Lewis" fucolipid was inactive) but also the 1 +2- fucosyl moiety of the exogenous acceptors. This could take the form of compartmentalization resulting in a different sub- cellular microenvironment, two different enzymes for each of the acceptors LcOse4Cer and H-1 fucolipid or the same enzyme recognizing the two different acceptors. These appar- ent K,,, values are much lower (by lo3) than those reported by Prieels et al. [ l l ] who used synthetic oligosaccharides and purified a1 +3/al+4-fucosyl transferase from human milk, although they also found a higher apparent K, (threefold) for lactotetraose than for the H-1 fucopentaose [ll]. Even given differences in assay procedures, taken together these results betoken a role for the non-glycosidic moieties of the glycosphingolipid molecules, and point to a need for studies with intact natural acceptors.
A major portion of these fucosyl transferases (52 - 53%) from SWll l6 cells appears to be associated with a high- density membrane fraction which is richest in Na+/K+- ATPase, has a high level of NADH - cytochrome-c reductase, and was followed in density by the mitochrondrial fraction as marked by the succinate dehydrogenase activity. The relative- ly high specific activity of UDP-GalNAc transferase (mucin acceptor) suggests that the light membrane fraction (peak 1) may be richest in enzymes of the Golgi apparatus. Taken together the data from Table 2 also suggest to us that a portion of these fucosyl transferases may be associated not only with microsomal cell fractions. It is tempting to speculate that this association may be at the level of trans Golgi and possibly the plasma membrane [35]. As with all density centrifugation techniques, our results carry the caveat of incidental cross- contamination between subcellular fractions.
Our results support the notion that in intact SW1116 cells the direct precursor of the Leb fucolipid is the H-3 fucolipid

21 7
which, in turn, is synthesized by the 1 -+2-fucosylation of LcOse4Cer. The first fucosylation competes for the LcOse4Cer pool with other biosynthetic pathways such as for Lea fucolipid and sialylated LcOse4Cer (IV3NeuNAcLcOse4Cer), the probable precursor of the 19/9 ganglioside [34].
The co-presence of Lea and Leb found in these cells was not observed in a survey of histologically normal human intestinal tissues but is characteristically found in meconia of ‘secretor’ and Lewis-positive individuals [28]. The Lea fucolipid in SW1116 cells and fetal intestine may theoretically arise from sialidase action on the 19/9 ganglioside. Alter- natively, Lea and Leb are synthesized because of a ‘primitive’ or tissue-specific expression of fucosyl transferases in these cells permitting the co-synthesis of these two fucolipids. Our data do not support the hypothesis of ‘aberrant’ fucosylation of Lea to Leb antigen [13] as a characteristic of oncogenesis. These results with human colorectal carcinoma cells (SW1116) do not allow us to extrapolate directly to Lewis antigenic expression in human erythrocytes or human milk globules.
I t has been suggested that the over-all conformation imposed by the presence of the fucosyl moiety may help ex- plain an increased affinity for an antibody (type XIV pneumococcal antipolysaccharide) of the lacto-N-fucopen- taose compared to the lacto-N-tetraose [36]. Theoretically, steric considerations may help to explain the differences be- tween the lacto-N-tetraosyl and lacto-N-fucopentaosyl acceptor activities reported here, and previously in other systems [ l l , 311. Furthermore, the presence of the lipophilic ceramide in the glycosphingolipid acceptors could restrict the number of allowed conformations or at least restrict the mo- lecular motion of the lacto-N-tetraosyl and fucopentaosyl (H-1) groups in comparison to the free oligosaccharides, in the presence of cell-free membrane preparations and detergent. This could then result in a higher accessibility or affinity of the oligosaccharide moieties of the purified glyco- sphingolipid acceptors (observed as lower apparent K,,, val- ues) for their membrane-bound fucosyl transferases.
Further purification of these enzymatic activities is in pro- gress and may provide further insights into their membrane localization, molecular diversity and mechanism of fucosyla- tion.
We thank the Centre National de la Recherche Scientifique (C.N.R.S 160104) and the Association for Cancer Research (n.415- 85) for financial support (V.A.L). We thank Alain Jolif for assistance with the sucrose gradient purifications and marker enzyme determinations in partial fulfillment of the D.E.A. degree. We thank Dr Jan Thurin (currently of the Wistar Institute, Philadelphia) for help in the LcOse4Cer purification. The preparation of glycolipid references and purified glycolipid acceptors was supported by grant 3967 from the Swedish Medical Research Council. We thank Dr M. Maunoury and Dr R. Lowy (Biostatistics Unit, CNRS, IRSC) for the statistical analyses, and for the use of the IBM (pc) computer. Special thanks to Drs Pierre Burtin, Charles Frayssinet Icard- Liepkans, Lars Svennerholm and George Cernaks for interesting discussions and helpful advice. Thanks to Mrs Dominique Chardaire for typing the manuscript.
REFERENCES 1 . Koscielak, J. (1963) Biochim. Biophys. Acta 78, 313-328. 2. Yamakawa, T., Irie, R. & Iwanaga, N. (1960) J. Biochem. (Tokyo)
48,490.
3. Hakomori, S. I. & Jeanloz, T. (1963) J. Biol. Chem. 239, 225- 228.
4. Hakomori, S. I. & Young, W. W. (1983) in Handbook of lipid research 3, Sphingolipid biochemistry (Hakomori, S. I. & Kanfer, J., eds) pp. 404-429, Plenum Press, New York.
5. Hakomori, S. I. (1984) Annu. Rev. Immunol. 2, 103-126. 6. Solter, D. & Knowles, B. B. (1978) Proc. Nut1 Acad. Sci. USA 75,
7. Blaszcyk, M., Pak, K. Y., Herlyn, M., Sears, H. F. & Steplewski, Z. (1985) Proc. Natl Acad. Sci. USA 82, 3552-3556.
8. Brockhaus, M., Magnani, J. L., Balsczyk, M., Steplewski, Z.. Koprowski, H. Karlsson, K. A., Larson, G. & Ginsburg, V. (1981) J . Biol. Chem. 256, 13223-13225.
9. McKibbin, J. M., Spencer, W. A,, Smith, E. L., Mansson, J. E., Karlsson, K. A., Samuelsson, B. E., Li, Y. T. & Li, S. L. (1982) J . Biol. Chem. 257, 755 -760.
10. Prohaska, R., Shenkel-Brunner, H. & Tuppy, H. (1978) Eur. J . Biochem. 84, 161 - 166.
11. Prieels, J. P., Monnom, D., Dolmans, M., Beyer, T. A. & Hill, R. L. (1981) J . Biol. Chem. 256, 10456-10463.
12. Campbell, C. & Stanley, P. (1984) J . Biol. Chem. 259, 11 208- 11 214.
13. Hakomori, S. I. (1985) Cancer Res. 45,2405-2414. 14. Beyer, T. A., Sadler, J. E., Rearick, J. I., Daulson, J. C. & Hill,
R. A. (1981) Adv. Enzymol. 23, 116. 15. Liepkans, V. A., Herrero-Zabaleta, M. E., Fondaneche, M. C. &
Burtin, P. (1985) Cancer Res. 45, 2255-2263. 16. Schacter, H., Jabbal, I., Hudgin, R. L., Pinteric, L., McGuire, E.
J. & Roseman. S. (1970) J . Biol. Chem. 245. 1090-1100.
5565 - 5569.
17.
18. 19.
20.
21.
22.
23.
24. 25.
26.
27.
28.
29. 30. 31.
32.
33.
34. 35. 36.
Knipe, D . M., Baltimore, D. & Lodish, H.’ F. (1977) Am. J .
Bradford, M. M. ( 1 976) Anal. Biochem. 72,248 - 254. Solomonson, L. P., Liepkans, V. A. & Spector, A. A. (1976)
Biochemistry 15, 892- 897. Solomonson, L. P. & Vennesland, B. (1977) Biochim. Biophys.
Acta 267, 544-551. Fitzgerald, D. K., McKenzie, J. & Ebner, K. E. (1971) Biochim.
Biophys. Acta 235,425-428. Coste, H., Martel, M. B., Assar, G. & Got, R. (1985) Biochim.
Biophys. Acta 814, 1 - 7. Liepkans, V. A,, Razafimahaleo, B., Gouyette, A,, Herrero-
Zabaleta, M. E. & Burtin, P. (1985) Biochem. Biophys. Res. Cornmun. 131, 736-742.
Virol. 21, 1128-1139.
Saito, T. & Hakomori, S. I. (1971) J . Lipid Res. 12,257-259. Leskawa, K. C., Dasgupta, S., Chien, J. L. & Hogan, E. L. (1984)
Magnani, J. L., Brockhaus, M., Smith, D. F. & Ginsburg, V.
Karlsson, K. A. & Larson, G. (1979) J . Biol. Chem. 254,931 1 -
Karlsson, K. A. & Larson, G. (1981) J . Biol. Chem. 256, 3512-
Spiro, R. G. & Spiro, M. J. (1965) J . Biol. Chem. 240,997 - 1000. Carlsen, R. B. & Pierce, J. G. (1972) J . Biol. Chem. 247, 23 -32. Johnson, P. H. & Watkins, W. M. (1965) in Glycoconjugates,
proceedings of the 8th international symposium (Davidson, E. A., Williams, J. L. & Ferrante, N. M., eds) pp. 222-223, Praeger Publishers, New York.
Magnani, J. L., Brockhaus, M. Zopf, D., Steplewski, Z., Koprowski, A. & Ginsburg, V. (1982) J . Biol. Chem. 257,
Liepkans, V. A. & Burtin, P. (1985) in Glycoconjugate,proceedings ofthe 8th international symposium (Davidson, E. H., Williams, J. C. & Ferrante, N. M., eds) p. 622, Praeger Publishers, New York.
Hanson, G. L. &Zopf, D. (1985)J. Biol. Chem. 260,9388-9392. Farquhar, M. G. (1985) Annu. Rev. Cell Biol. 1,447-488. Biswas, M. & Rao, V. S. R. (1980) Biopolymers 19, 1555-1566.
Anal. Biochem. 140, 172- 177.
(1982) Methods Enzymol. 83, 235-241.
9316.
3524.
14365 - 14369.
















![Équations Différentielles Stochastiques Rétrogrades[PP92] , Backward stochastic differential equations and quasilinear parabolic partial differential equations, Stochastic partial](https://img.pdfslide.fr/doc/110x75/5f3f690470d8062e9676eb02/quations-diirentielles-stochastiques-r-pp92-backward-stochastic-diierential.jpg)












