Embed Size (px)
Citation preview

REVUE DE MICROPALI~ONTOLOGIE Vol. 40, n ° 4, d6cembre 1997, pp. 313-329
CAN DETAILED SAMPLING AND TAPHONOMICAL ANALYSIS OF FORAMINIFERAL ASSEMBLAGES OFFER NEW DATA
FOR PALEOECOLOGICAL INTERPRETATIONS?
UN [~CHANTILLONNAGE DETAILLE ET UNE ANALYSE TAPHONOMIQUE D'ASSOCIATIONS DE FORAMINIFE, RES
PEUV.ENT-ILS OFFRIR DE NOUVELLES DONNEES POUR LES INTERPRETATIONS PALEOECOLOGIQUES ?
by Katar~na HOLCOVA*
ABSTRACT. - - Distinctions between the composition of foraminiferal assemblages in lithologically different superimposed beds were established in the Lower and Middle Miocene samples from the South Slovak Basin included in the Central Paratethys. These distinctions may be caused by different depositionaÁ mechanisms of the individual beds.
Foraminiferal assemblages from the beds were analysed quantitatively. A detailed taphonomical study of the oryctocenoses preceded paleoecological interpretations. Reworked, transported (in suspended load and bed load) and indigenous foraminiferal tests were recognized in assemblages. The distribution of these tests in beds was recorded. Three types of distribution were recognized: (1) only suspension-transported foraminifers occuring in both coarser and finer beds, (2) indigenous and reworked foraminiferal tests occuring with different ratios in the beds, (3) tests transported within bed load occuring in coarser beds, in finer beds they may be accompanied by reworked or indigenous tests.
The presented analysis of foraminiferal oryctocenoses from different beds supphes data on depositional environments and source areas of the sedimentary particles bearing foraminiferal tests, as well as on their transport mechanism. Analyses showed that sampling of the same lithotype in whole sections (e.g. the finest sediments) may give fragmentary or even misleading data (better preserved indigenous assemblages may occur in coarser sediments).
RI~SUME. -- Des diff6rences de composition entre les associations de foraminif~res issues de couches superpos6es de lithologie diff6rente ont 6t6 6tablies pour des 6chantillons du Miocb~ne inf6rieur et moyen de la Parat6thys Centrale (Juhoslovensk~ panva). Ces 6carts peuvent ~tre dus ~ des diff6rences dans les m6canismes de d6p6t des couches individuelles.
Les associations de foraminif~res des diff6rentes couches ont 6t6 analys6es quantitativement. Une 6tude taphonomique d6taill6e de l'o~yctoc6nose est suivie d'interpr6tations pal6o6cologiques. Des tests de foraminif~res remani6s, transport6s (en suspension ou en traction) et autochtones sont reconnus dans les associations. La distribution de ces tests dans les couches a 6t6 not6e. Trois types de distribution ont 6t6 reconnus : (1) les foraminif~res transport6s en suspension sont senls pr6sents, (2) les tests de foraminif~res autochtones et remani6s sont pr6sents dans des proportions variables, (3) les tests transport6s par traction sont pr6seuts dans les couches les plus grossi~res; dans les plus fines, ils peuvent ~tre accompagn6s de tests remani6s ou autochtones.
Ces analyses d'oryctoc6noses de foraminif~res apportent des informations sur les milieux de d6p6t, sur l'origine des particnles s6dimentaires accompagnant les foraminifbres et sur les m6canismes de transport. L'6chantillonnage d 'un seul et m~me facies dans une coupe complete (p. ex. les s6diments les plus fins) peut n 'apporter que des indications fragmentaires ou erron6es (par exemple, les associations les mieux pr6serv6es peuvent se trouver dans les s6diments les plus grossiers).
Key-words : Foraminifera - Taphonomy - Depositional environment - Miocene - Central Paratethys - Palaeoenviromnent.
Mots-cl6s : Foraminif~res - Taphonomie - Milieu de d6p6t - Miocene - Parat6thys Centrale - Pal6oenvironnement.
* Department of Paleontology, Charles University, Albertov 6, CZ-128 43, Czech Republic.

314 PALEOECOLOGICAL ANALYSIS OF FORAMINIFERAL ASSEMBLAGES
I N T R O D U C T I O N
Foraminiferal tests included in sediments provide two kinds of information about the paleoenviron- men t : (1) foraminifers need specific life habitats during their life, (2) tests of dead foraminifers are t ransformed into sedimentary particles and, as such, they can be transported and .transformed.
The starting point of this work is a paleoecolo- gical study of foraminiferal assemblages obtained from millimeter- to centimeter-thick lithologically different beds in clastic sediments (siltstone and sandstone). Paleoecological interpretations are based on a taphonomical analysis of the assemblages. It is supposed that the compared beds were deposited in a similar paleoenvironment by different deposi- tional mechanisms. Therefore, differences in species composition, size distribution and preservation of foraminiferal tests in beds were caused by these depositional mechanisms. This work aims to show the influence of test t ransport on the composition of foraminiferal assemblages, and then on paleoen- vironmental interpretations based on this composi- tion.
In the study of clastic sediments, environmental interpretation based mainly on the grain-size dis- tribution of sedimentary particles provides an ex- planation of the t ransport mechanism (e.g. Folk and Ward, 1957 ; Friedman, 1979 ; Middleton, 1976 ; McCave, 1 9 7 8 ; McLaren, 1 9 8 2 ; Visher, 1 9 6 9 ; Singer and Anderson, 1 9 8 4 ; etc.). But interpreta- tions of grain-size distribution are equivocal in many cases. McLaren and Bowles (1985) gave two reasons for the disagreement in interpretations : the process of erosion and deposition of particles involves many variables, which cannot be measured very well; the grain-size distribution changes continuously fi'om source to final deposits by multiple t ransport or other sedimentary processes (winnowing, selective deposition, etc.), which cannot be easily recognized.
Foraminiferal tests differ from other sedimentary particles by certain characteristics. Therefore, se- dimentological models are not fully applicable on foraminiferal assemblages. The size distribution of t ransported foraminiferal tests can be different from the distribution of clasts. It can be caused by :
(1) difference in the size distribution of source deposit ( log-normal: Udden, 1914) and "source" (= indigenous) foraminiferal assemblages (polymo- dal, flat distribution; shape of distribution curve depends on species composition);
(2) t ransport and resedimentation of foraminife- ral tests, that usually do not show as many recur-
rences as those of sedimentary particles. Therefore, foraminiferal tests document mainly the last rewor- ked episodes of sedimentary particles;
(3) predisposition to t ransport under certain phy- sical conditions, that depends on the ratio between volume and weight of transported particles. Diver- sity of morphotypes of foraminiferal tests is mar- kedly larger than tile morphological diversity of clasts. Therefore, an accumulation of some forami- niferal morphotypes can provide detailed informa- tion about t ransport mechanisms.
In spite of this, the size range of size-sorted tests has provided a distinction between the suspended- load and bed-load t ransport (Murray, 1965 ; Loose, 1 9 7 0 ; Culver and Banner, 1 9 7 8 ; Murray et al., 1 9 8 2 ; Wang and Murray, 1983).
The interpretation of the t ransport mechanism of foraminiferal tests can be made more easily than that of inorganic sedimentary particles. The follo- wing characters can be utilized (Murray, 1 9 9 1 ) : absence of juveniles or adults, breakage and abra- sion of foraminiferal test, different modes of test preservation. A discrepancy between ecological re- quirements of species in the assemblages may corres- pond to different life habitats of source and depo- sition areas. Differences in stratigraphical range of some species in the assemblages may indicate a succession of reworking features.
Therefore, t ransported and resedimented tests are able to provide more information about the sediment story than a single grain-size distribution. On thc other hand, the paleoecological interpreta- tion of a test t ransport requires the experience of a sedimentologist.
M A T E R I A L
The analysed material comes from the Miocene of the South Slovak Basin (included in the Central Paratethys) (Fig. 1). Two core-samples and two sections were studied from the "Rzehakia (Onco- phora) Beds" and 10 core-samples were selected from the Lower Badenian interval.
"Rzehak ia (Oncophora) B e d s "
The "Rzehakia (Oncophora) Beds" represent an important Upper Ottnangian (= Lower Miocene, equivalent of the middle par t of the Burdigalian : Papp et al., 1973) Paratethys horizon bearing a characteristic endemic molluscan fauna. Generally,

HOLCOVA 315
r~
y
• LOWER BADENIAN
FIG. l . - Locations of the studied cores and exposure.
Localisation des carottes et affleurement ~tudi&
the "Rzehakia Beds" were deposited in shallow brackish seas or large brachyhaline to oligohahne lagoons (Kvaliashvili, 1 9 6 2 ; ~tyrok)~, 1968) . In the southern par t of the Paratethys, these brackish basins were periodically connected with the open Lower Miocene sea. In the majority of the Parate- thys basins, the Beds represent a regressive phase of the overlying Lower Miocene (in tile Central Paratethys Eggenburgian - Lower Ottnangian) cycle of seadevel changes.
In the Pannonian Basin (the studied area - the South Slovak Basin - represents the northern par t of the Pannonian Basin), the "Oncophora (Rzeha- kia) Beds" seem to transgressively overlie the conti- nental Ottnangian sedimentation. Therefore, the Beds are proposed by some authors to be a trans- gressive phase of younger Karpat ian cycle of sea- level changes (e.g. Vass, 1 9 9 5 ; Hhmor, 1985) .
The lowstand character of the beds, very similar lithotypes in the whole Paratethys region and a significant influence of storm activity (tempestites, foraminiferal tests transported by storm waves) enable one to propose an isochroneity of the "Rzehakia (Oncophora) Beds" (in agreement with the opinion of Kovh~ et al., 1989). The relative transgressive or regressive position of the Beds may be caused
by tectonic control of sea-level changes (Holcovh- ~utovsk~, 1996) .
The "'Rzehakia (Oncophora) Beds" in the South Slovak Basin (Vass et al., 1979 , 1992) are repre- sented by massive or laminated fine-grained sand- stone. The massive sandstone is intensively biotur- bated. The laminated sandstone contains laminae or thin beds of claystone, sihstone or coarse-grained sandstone. Samples issued from these well-indivi- duahzed siltstone and sandstone were chosen for analysis (Fig. 2 ; P1. 1, fig. 1-3).
An abundant occurrence of foraminiferal tests is a distinctive feature of the South-Slovak "'Rzehakia (Oncophora) Beds". Small, thin-walled, and well- preserved tests prevailed in the analysed samples. The abundance of these tests is high, foraminiferal numbers (= number of foraminiferal tests in 1 g of sediment) reach values of 800. Benthonic forami- niferal assemblages are dominated by outer shelf to bathyal species, e.g. Globoc~a~sidulina, Cassidulina, Cibicidoides, Trifarina, Bolivina~ Uvigerina, Episto- maria, Caucasina, Spiroloxostoma, etc. In some horizons, inner shelf assemblages dominated by Am- monia ex gr. parkinsonia-tepida occur. Planktonic foraminifers are represented by Globigerina ex gr. praebulloides BLOW, Cassigerinella boudecensis POK. and small microperforate pentacamerate spe- cies (Tenuitella plexus). In some horizons, large and well-preserved tests occur among the small thin-wal- led tests. The large tests included hyposaline spe- cies : Haynesina depressula (WALKER and JACOB), Monspeliensina vulpesi GLAGON and LYS and Am- monia parkinsonia-tepida group. Stenohaline, epi- phytic species Elphidium div. sp., Lobatula lobatula (WALKER and JACOB), Hanzawaia boueana (d'OR- BIGNY), Asterigerinata planorbis (d'ORBIGNY) w e r e found very rarely.
Two significant cyclical changes of foraminiferal oryctocenoses were recognized for the South-Slovak "Rzehakia (Oncophora) Beds" using: (1) changes in the ratio of indigenous, suspension-transported, bedload-transported and reworked tests, and (2) changes in the abundance of indigenous tests. Using these characteristics, two cycles of sea-level changes of the 5th order may be interpreted for the South-Slovak "Rzehakia (Oncophora) Beds" (Uoleovh, 1996b) (Fig. 2).
T h e E a r l y B a d e n i a n
The Early Badenian sediments (equivalent of the Langhian) in the South Slovak Basin characterize, in a transgressive position, the Early Badenian sediments in the whole Paratethys region (e.g. R6gl

316 PALEOECOLOGICAL ANALYSIS OF FORAMINIFERAL ASSEMBLAGES
Z
~D
N -83
N-96 N-65
~ 2
~ 4 ~ s ~ 6
96 ~ 12
117'" ~ 13
MK=I TM
MK-2
- - - 20 -- 21
'~A 15
FIG. 2. - Lithological sections of the s tudied cores N-48 , N - 6 5 , N-96 , N - 8 3 , N-95 and exposures f rom sandpi t Mal~ Krti~ (MK 1, MK 2). Explana t ions : 1 - c laystone, 2 - sands tone , 3 - conglomera te , 4 - siltstone, 5 - clayey sands tone , 6 - tuffaceous sands tone , 7 - g r a d e d bedding, 8 - l amina ted sands tone , 9 - sandstone with undula t ing lamina t ion , 10 - cross-bedding, 11 - siltstone with sandstone laminae , 12 - b recc ia with volcanic mater ia l , 13 - tuffite wi th shell de t r i tus , 14 - sands tone lenses accumula t ed in t roughs of wave r ipples , 15 - b io tu rba t ions , 1 6 - 1 8 - pa l eoenv i ronmen ta l in te rpre ta t ions . ( l 6 - lacus t r ine sediments , 17 - h t to ra l , hyposal ine env i ronmen t , 18 - l i t toral , s tenohal ine env i ronmen t ) , 19 - h ia tus , 20 - b o u n d a r y be tween cycles of sea-level changes, 21 - cor re la t ion
of h ighs tand t racts . Samples a re f r a m e d and the i r depths a r e given. F o r detailed lithology of samples see Fig. 3 -5 .
Coupes l i thologiques des carot tes dtudi~es N ~ 8 , N - 6 5 , N - 9 6 , N 8 3 , N - 9 5 et des a f f l euremen t s de la sabli~re Ma ly Kr t i s ( M K 1 , M K 2 ) . L@eudes : 1 - argilol i thes, 2 - gr$s , 3 - conglom~rats , 4 - s i l ts tone, 5 - gr$s argi leux, 6 - gr~s tufJhcd, 7 - g r a n o c l a s s e m e n t , 8 - gr$s lamin~, 9 - gr~s ~t l amina t i on ondul~e, 10 - d6p~ts entrecrois~s, 11 - s i l ts tone & lamines gr~seuses, 12 - br$che ~t mater ie l volcanique , 13 - tu f f i te ~t ddbris coquill iers, 14 - lentilles gr6seuses accumul~es dans les c r e u x de rides de houle, 15 - b io turbat ions , 1 6 - 1 8 - i n t e rpre ta t ion pa l~oenv i ronnemen ta l e ( i 6 - s~diments lacustres , 1 7 - e n v i r o n n e m e n t l i t toral hypoha l in , 18 - e n v i r o n n e m e n t l i t toral s t~nohal in) , 19 - h ia tus , 2 0 - l imites en tre cycles de var ia t ions d u n i veau mar in , 21 - corre la t ion des corteges de h a u t n iveau
mar in . Les ~chanti l lons sont localisds, et l eur p r o f o n d e u r indiqu~e. Voir f i g . 3-5 p o u r la litologie d~taill~e des ~chanti l lons.
and Steininger, 1983) . The sedimentation in South Slovakia was significantly influenced by volcanic activity. The studied boreholes are situated in the marginal par t of the Early Badenian sea. Tuffaceous sandstones and sihstones with common bioturbation dominated by domichnia overlie the basal coarse volcaniclastics (Fig. 2 ; P1. 1, fig. 4-7). Well-indivi- dualized laminae to thin beds of siltstone and sand- stone characterize the whole sections, what allowed more detailed sampling of the N-83 borehole.
The benthonic foraminiferal assemblages in bo- reholes N-95 and N-83 are dominated by epiphytic, inner shelf, stenohaline species : Pararotalia stellata (REUSS), Lobatula lobatula (WALKER and JACOB), Asterigerina planorbis (d'ORBIGNY), Glabratella pa- telliformis (d'OBBIGNY), elphidiids and miliolids. The rare occurrence of Praeorbulina and Orbulina allows biostratigraphical interpretation. The species composition of the assemblages as well as the per- centage of species in assemblages are very similar

HOLCOVA 317
in the whole sections. In the nearshore area (N-48 borehole), hyposaline assemblages dominated by the Ammonia parkinsonia-tepida group and Haynesina depressula (WALKER and JACOB) occur very rarely in the middle par t of the sections.
Cychcal changes in these very similar, upper neritic foraminiferal assemblages are not very pro- minent. Cyclical changes in the foraminiferal abun- dance, part ly in the epifauna/infauna ratio and in the similarity, were used for the interpretation of cycles of sea-level changes. Three cycles, probably of the 4 th order, may be distinguished (Holcov~, 1996b) .
M E T H O D S
Well-individualized, superimposed thin beds, re- presented by coarse and fine sediment (sihstone/fine sandstone, fine sandstone/coarse sandstone, sand- stone/coarse conglomerate) were washed separately using a 63 ~m (240-mesh) sieve. Weight percentage of the sand fraction (> 63 ~tm) was entered for every sample. For comparison, a par t of the whole sample containing both lithotypes was washed.
S t a n d a r d a n d q u a n t i t a t i v e analys is o f f o r a m i n i f e r a l a s s e m b l a g e s
Analysis of foraminiferal assemblages includes :
(1) determination of species occurring in assem- blages (list of species for boreholes N-83 and N-95 was published in Holeov~ et al., 1996) ;
(2) quantitative analysis of assemblages. The fol- lowing four quantitative data, which can be used in paleoenvironmental interpretation, were given for all analysed beds (Fig. 6 below) :
- - foraminifera1 number of Scott (= number of foraminiferal tests in 1 g dry weight of sedi- ment) ;
- - P/B-ratio (= percentage of planktonic forami- nifcrs in foraminiferal assemblage calculated for 300-500 specimens);
- - diversity of benthonic foraminiferal assem- blages calculated for 300 specimens using Simpson's formula :
N ( N - l ) D -
Ni (N~-I)
where D is diversity, N is the number of counted specimens in the assemblage and Ni is the number of specimens of the i-th species.
- - similarity of foraminiferal assemblages from superimposed beds calculated for 300 speci- mens using Jaccard ' s formula.
C S -
C + U
where S is similarity, C is the number of species occurring in both compared samples and U is the number of species occurring only in the first sample + number of species occurring only in the second sample.
T a p h o n o m i c a l analysis o f f o r a m i n i f e r a l o r y e t o e e n o s i s
For recognizing the influence of t ransport on foraminiferal tests, the following criteria were stu- died :
(1) Size distribution o f tests : from every sampled bed, 100 rounded, 50 elongate benthonic forms and 50 planktonic foraminiferal tests were chosen for the analysis from > 63 ~m (240-mesh) sieve fraction. The largest diameter of tests for rounded forms and planktonic foraminifers and the length of tests for elongate forms were measured using the VIA video measuring system. Size distributions of tests (separately for rounded forms, elongate forms and planktonic foraminifers) were summarized in histograms.
For recognizing the t ransported and autochto- nous assemblages, size sorting was deduced from histograms : narrow, unimodal histograms characte- rize size-sorted tests, polymodal histograms with great range of test size are typical of unsorted tests.
To determine the mode of t ransport , a range of test sizes were studied among transported tests. Small tests (rounded forms < 200 ~tm, elongate forms < 300 ~m) indicate a suspended-load trans- port . Adults of a majority of species absent in the oryctocenoses were t ransported within a suspended load.
Large well-sorted tests (> 300 ~m) have been t ransported within a bed load (Murray, 1 9 6 5 ; Loose, 1 9 7 0 ; Culver and Banner, 1 9 7 8 ; Murray et al., 1 9 8 2 ; Wang and Murray, 1983). Juveniles are absent in oryctocenoses t ransported within a bed load.
These results are also applicable to fossil assem- blages (Holcov~, 1996a).
(2) Preservation o f tests determined by SEM examination : well-preserved and corroded/abraded tests were recognized. Significant diagenetic changes of the tests' walls were not observed.

318 PALEOECOLOG1CAL ANALYSIS OF F O R A M I N I F E R A L ASSEMBLAGES
(3) P a l e o e c o l o g i c a l i n t e r p r e t a t i o n o f s p e c i e s in assemblages were based mainly on data of Pfleger (1965) , Boltovskoy and Wright (1976) , Mur ray ( 1 9 7 3 , 1991) , Reiss and Hott inger (1984) . Diffe- rent paleoecological requirements of species in an assemblage indicate the occurrence of t r anspor ted tests.
(4) R e w o r k e d s p e c i e s can be distinguished by comparison of biostrat igraphical ranges of species in assemblages (mainly Cicha e t a l . , 1 9 8 3 ; RSgl, 1986) .
R E S U L T S
T y p e s o f d i f f e r e n t l y p o s t m o r t e m - m o d i f i e d f o r a m i n i f e r a l o r y c t o c e n o s e s
Based on the taphonomical analysis of foramini- feral oryctocenoses, six types of oryctocenoses were distinguished in the analysed material using previous resuhs (Holcov~, 1996a ) (Tab. 1 ) :
(1) Foraminiferal tests in oryctoccnoses are size- unsorted, well-preserved and exhibit different onto- genetic stages. Ecological requirements and strat igraphical ranges of species are identical. These parameters character ize indigenous or weakly t rans- por ted tests without any influence of t ranspor t on their composition and preservation.
(2) The oryctoeenosis is composed of size-sorted, small ( rounded tests < 200 ~m, elongate ones < 300 ~m), well-preserved and thin-walled tests, with juveniles being dominant . Outer shelf to ba- thyal species occur in assemblages. This type of oryctoccnoses was formed by t ranspor t in suspen- sion.
(3) Tests t r anspor ted in bedload are size-sorted and essentially > 200 ~m. They are abraded , bro- ken and dominated by robust forms. Lit toral species occur in this type of oryctocenoses.
(4) Two types of test preservat ion were observed in this oryctoccnosis : some tests are unsorted, well- preserved and belong to outer to inner shelf species ; the others are size-sorted, large, broken, abraded, and belong to euryhal ine and littoral species. The strat igraphical ranges of all species in assemblages are identical. The oryctocenosis contains indigenous or weakly t ranspor ted elements mixed with tests t ranspor ted in bed load f rom a coastal area to a more central pa r t of the basin.
TAB. 1. - Types of differently postmortem modified foraminil~ral oryctocenosis. Abbreviations: corr. = corroded, us = unsorted, sl = size-sorted, large tests (larger than 200 ~m), ss = size-sorted, small tests (< 200 ~m), hs = hyposaline environment, is = inner shelf, stenohaline, os = outer shelf, ba = bathyal, LM = Lower Miocene, LB = Lower Badenian, ident. = identical for all species in sample, u.b. = in underlying beds. For detailed explanations
see text. Divers types d'associations ~bssiles de forautinigQres d i f ~ r a n t p a r les modif icat ions pos tmor t em subies. Abr~viat ions : corr. = cor- rode, us = non classd, sl = grands tests classes selon leur taille (sup~rieure & 2 0 0 ~tm), ss = pet i ts tests classes selon leur taiUe ( inf~rieure h 200 ~m), hs = env i ronnemen t hyposal~, is = p la- te forme interne, s tdnohaline, os = p la t e forme ex terne , ba = ba thyva l , LM = Miocene inf~rieur, LB = B a d d n i e n inf$rieur, ident . = c o m p a r a b l e p o u r toutes les esp~ces de l '6chant i l lon , u .b . = d a n s les n i v e a u x souss jacen t s . Voir le tex te p o n r les
l@endes ddtaill~.es.
== ~ ~ ~ o z~NN'E~ • ~ ~ O~ ~ ~ ~ ~ ~ 0 ~ " ~ "~ Interpretation
_ ~ ~ of postmortem ~- ~o ~_ ~ '~ > ~ ~.° ~= ~ changes
- . . I ~ , I . . . . ,,l~or, i j . v l . d h,li, lo, lba LM ILB I x x x x ident, ident, indigenous or weakly transported
- I 2 x x x rare x x x suspension-transported
3 x x are x x x ident, bedload-transported
x x x x x x indigenous mixed 4 ident, with bedload transported
x x x x x indigenous with
5 x x x x x x suspension-t ra nsport ed x x x rare x x x and then reworked
suspension-transported i 6 x x x rar~ x I x x u .b and then reworked _
(5) The size distl~budon of tests in the orycto- cenosis is polymodal with a great peak at the value of about 165 ~m. All the tests are well-preserved. Small ones are thin-walled, issued f rom outer shelf to ba thyal environments and often represented by juveniles only. Index species of the Lower Miocene were observed among them. Species with size-un- sorted tests are issued f rom inner shelf environments with index species of the Lower Badenian. Both juvenile and adult specimens occur in assemblages. The oryctocenosis is composed of indigenous or little t ranspor ted , shallow marine, Lower Badenian as- semblages with reworked Lower Miocene suspension- t ranspor ted tests.
(6) The oryctocenosis consisting of only Lower Miocene suspension- t ranspor ted tests reworked to Lower Badenian sediments is character ized by size- sorted, small, well-preserved or cor roded , thin-wal- led tests belonging mainly to juveniles. Outer shelf and ba thyal species occur in this oryctocenosis. Only the Lower Miocene index species were found, while in the underlying and overlying samples Lower Badenian index species occur.

H O L C OVA 3 1 9
TAB. 2. -- Types of distribution of foraminiferal oryctocenosis in superimposed thi~l beds characterized by different lithologies, lx - 1st postmortem transport of tests, 2x = 2nd postmortem transport of tests, in = indigenous tests, st = suspension-trans- ported tests, bt = bedload-transported tests, rw = reworked tests, LB = Lower Badenian, LM = Lower Miocene. 1 - transport of foraminiferal tests in suspension, 2 - resedimentation of forami- niferal tests, 3 - mixing of foraminiferal assemblages by biotur- bation, 4 - transport of foraminiferal tests in bedload, 5 - hyposaline environment, 6 - inner shelf, stenohaline, 7 - outer shelf, 8 - posssible misinterpretation of paleoenvirorm)ent, 9 - sandstone, 10 - siltstone, l l - elaystone, 12 - undulating lami- nation, 13 - graded bedding. For detailed explanations see text.
Types de distribution des associations fossiles de foraminif~res dans des lits minces superposes, caract~ris~s par des lithologies diff~rentes, lx = premier transport post-mortem des tests, 2x = 2 ~ transport post-mortem des tests, in = tests autochtones, st = tests transport~s en suspension, bt = tests transport~s par trac- tion, rw = tests remanids, LB = Bad~nien infdrieur, LM = Miocene inf~rieur, 1 - transport en suspension des tests de foraminiJ~res, 2 - res~dimentation des tests, 3 - rr~lange des associations de foraminij~res par bioturbation, 4 - transport des tests par traction, 5 - environnement hyposal$, 6 - plateforme interne, st~nohaline, 7 - p l a t e f o r m e externe, 8 - risques d'erreur dans l~nterpr~tation du pal~oenvironnement, 9 - gr$s, 10 - siltstone, 11 - argilolithe, 12 - lamination ondul~e, l 3 - granoclassetwsnt.
Voir le texte pour les l@endes d~taill~es.
D i s t r i b u t i o n o f t y p e s o f o r y c t o c e n o s e s i n b e d c o u p l e s
F o r a m i n i f e r a l o ryc tocenos i s f r o m pa i r s of well i n d i v i d u a l i z e d th in beds of f ine ( d o m i n a t e d by s ih- s tone) a n d c o a r s e ( d o m i n a t e d by s ands tone ) sedi- ments w e r e t a p h o n o m i c a l y a n a l y s e d a n d c lass i f ied in to six types of o ryc tocenoses f r o m Tab . 1. T h e d i s t r i b u t i o n o f o ryc tocenoses in ana ly sed samples def ines the fo l lowing types a n d sub types (Tab . 2) :
T y p e A : O n l y s u s p e n s i o n - t r a n s p o r t e d
f o r a m i n i f e r s o c c u r r i n g i n t h e s e d i m e n t
S u b t y p e A a : S u s p e n s i o n - t r a n s p o r t e d f o r a m i n i f e r s (o ryc tocenos i s 2) o c c u r in b o t h c o a r s e r a n d f ine r beds (Fig. 3) .
F o r c o m p a r i s o n wi th o t h e r a n a l y s e d samples , the exc lus ive o c c u r r e n c e of s u s p e n s i o n - t r a n s p o r t e d fo- r a m i n i f e r s is n o t e d usua l ly in t he f ines t s ed imen t s (Fig. 6) , w h a t is in a g r e e m e n t wi th t he o b s e r v a t i o n of W a n g a n d M u r r a y ( 1 9 8 3 ) . Changes in g r a in size, e x p r e s s e d by p e r c e n t a g e o f s and in s ed imen t , h a v e no i n f l u e n c e on the r a n g e of test size. T h e specif ic c o m p o s i t i o n o f assemblages is s imi l a r ( G l o b o c a s s i -
d u l i n a div. sp. , C a s s i d u l i n a l a e v i g a t a , I s l a n d i e U a
sp., B o l i v i n a a n t i q u a , C a u c a s i n a s c h i s c h k i n s k a y a e
a n d C i b i c i d o i d e s p s e u d o u n g e r i a n u s d o m i n a t e ) , w h a t can be c a u s e d by some m o r p h o l o g i c a l c h a r a c t e r i s t i c s of tests s u c h as th ickness of tes t wal l , size, a n d
C"-
] i B ~ iii!~i~ <
°11
~ ~ X X X ~
1 × ×
_:_7
N t ype of o r n a m e n t a t i o n p r e d i s p o s i n g t h e m to t r a n s - p o r t in suspens ion . T h e r e f o r e , the i ndex of s imila- r i t y f o r the assemblages is h igh ( 6 8 - 9 4 %) (Fig. 6) . Th i s t y p e of f o r a m i n i f e r a l a s semblage t r a n s p o r t e d by suspens ion a n d r e c o g n i z e d in all a n a l y s e d beds occu r s on ly in the " R z e h a k i a ( O n c o p h o r a ) B e d s " .
P a l e o e n v i r o n m e n t a l i n t e r p r e t a t i o n : T h e o u t e r she l f to u p p e r b a t h y a l s t e n o h a l i n e p a l e o e n v i r o n m e n t c a n be c o n s i d e r e d as the s o u r c e a rea . T r a n s p o r t o f

Ao
Ab
MK
1
o',
" -o
"oo
•
"o'o
, "
,o
,,
°,
.o
~
•
r.e~
.'k/.e
m..v
~.k
3¢..m
~.V.
r.t.
V..~
k.V.
mL"
k,r~
el
:-"'
"'(i
'"'"
'"'"
'""'
"*'"
"'.'
.'.'
-:
: :
! ;
....
.
::::
::::
::::
::::
::::
::::
::::
::::
::::
::::
::::
::::
::::
::::
::::
iiiiiiiiii
i i i i;i;',;i
iiiiii,i,i
il;i!i;i i;
i ;i :1
BE
NTH
ON
IC F
OR
AM
INIF
ER
A-
RO
UN
DE
D F
OR
MS
IO
t3
o ~
038
~27
' 0,
36
' 'o
,=5'
~ O
N/C
FO
RA
MIN
IFE
RA
1o
MK
14
..
..
..
..
..
..
..
..
13
0~0~
0,
18
c~27
(~
3s
~5
O
,S4
MK
2 -;-~:Tf
f:-':':':'
:':"- :~
'"7"":"
o.
N-96
/305
sandstone
~.-
-
~~
_~
ii
il
B
EN
THO
NIC
FO
RA
MIN
IFE
RA
- R
OU
ND
ED
FO
RM
S
. -7
i.
~~..
....
-..;
.;..
..:.
;.
I ac
e o,1
8 a~
o.
ze
a.~
0,~
~65/
120
san
dst
on
e
/
i~48
/95
~d.~
'one
::::::::::
::::::::::
::::::::::
::::::::::
:::::~ O~o
.
..
.
:':'
:':'
":':
""""
" .
..
..
..
c~aystone
,o :
/
~i:i::+:i:::::~":-':':"~
" ~
BE
NTH
ON
IC F
OR
AM
INIF
ER
A-
ELO
NG
ATE
FO
RM
S
°o.c~
O,
lS
o.:-.
, o.
~ k.
e ~
.~
SIZ
E-D
ISTR
IBU
TIO
N O
F T
ES
TS
FR
OM
SA
MP
LES
MK
11 -
MK
14
o ~
BE
NTH
ON
IC F
OR
AM
INIF
ER
A-
RO
UN
DE
D F
OR
MS
1o
PLA
NK
TON
IC F
OR
AM
INIF
ER
A
..
..
..
.
21
BE
NTH
ON
IC F
OR
AM
INIF
ER
A-
ELO
NG
ATE
FO
RM
S
t,~
24
..
..
..
21
so •
SIZ
E-D
ISTR
IBU
TIO
N O
F TE
STS
~
FRO
M S
AM
PLE
S M
K21
- M
K 2
4
o
BE
NTH
ON
IC F
OR
AM
INIF
ER
A-
ELO
NG
ATE
FO
RM
S
PLA
NK
TON
IC F
OR
AM
INIF
ER
A
0,0~
0,
18
0,27
0,
36
0,4~
0,
54
1 j
~o
~ N
-96/
305
m -
silt
ston
e
a a
~
E:~
N
-65/
120
silts
tone
[~
N-4
8/95
m -
silt
ston
e
o 0,
0~
0,18
0,
27
0,36
0,
45
0,54
0,
0~
0,18
0,
27
0,36
0.
45
0,54
6 7
SIZ
E-D
ISTR
IBU
TIO
N O
F TE
STS
FR
OM
WH
OLE
SA
MP
LES
o
N-6
5/12
0 M
o,oe
o,te
o,
z~
o,:~
o,
,~
o,~
vert
ical
c0(
is-n
umbe
r of
spe
cim
ens
horiz
onto
l c0
(Is -
test
stz
e (u
rn)
©
©
©
©
©
k=m
;z
CI3
FIG
. 3
. -
Siz
e d
istr
ibu
tio
n
of b
enth
on
ic
rou
nd
ed,
ben
tho
nic
el
on
gat
e an
d p
lan
kto
nic
fo
ram
inif
eral
tes
ts i
n sa
mp
les
N-9
6/3
05
m
, N
-65
/12
0
m,
N-4
8/9
5
m
and
se
ctio
ns
MK
1
and
M
K 2
:
typ
e A
w
ith
su
spen
sio
n-t
ran
spo
rted
te
sts.
E
xp
lan
atio
ns
of l
ith
olo
gic
al s
chem
es :
1
- si
hst
on
e,
2 -
san
dst
on
e,
3 -
clay
sto
ne,
4
- sa
nd
sto
ne
wit
h p
lan
t an
d
shel
l d
etri
tus,
5
- sa
nd
sto
ne
wit
h l
um
ach
elle
bed
s, 6
-
7 -
dis
tin
ct l
amin
ated
str
uct
ure
, 8
- in
dis
tin
ct l
amin
ated
str
uct
ure
. L
eng
ht
of s
cale
bar
:
2 cm
. S
ub
typ
es A
a an
d A
b
are
def
ined
in
tex
t.
Dis
trib
uti
on
d
e ta
ille
de
s te
sts
de f
ora
min
iJ~
res
ben
thiq
ues
a
rro
nd
is,
ben
thiq
ues
al
long
~s
et p
lan
cto
niq
ues
, d
an
s le
s $c
hant
illo
ns
N-9
6/3
05
m
, N
-65
/12
0
m,
N-4
8/9
5
met
d
an
s le
s co
up
es
MK
1
et
MK
2
: ty
pe
A,
?t t
ests
tr
an
spo
rt6
s en
su
spen
sio
n.
L~
gen
des
de
s fi
gu
rds
lith
olog
ique
s :
I -
silt
ston
e,
2 -
grbs
, 3
- ar
gilo
lith
e,
4 -
grb
s h
d~br
is
de
pla
nte
s et
de
co
quil
les,
5
- gr
$s ?
t ba
ncs
de l
um
ach
elle
, 6
-7
- st
ruct
ure
la
min
ae
dis
tin
cte,
8
- st
ruct
ure
la
min
ae
ind
isti
nct
e.
Lo
ng
ueu
r de
la
ba
rre
d'~
chel
le
: 2
cm.
Les
so
us-
typ
es
Aa
et
Ab
so
nt
d~fi
nis
du
ns
le t
exte
.

HOLCOVA 321
benthonic foraminifers in suspension is caused by storm waves. Absence of indigenous foraminifers does not allow one to interpret a paleoenvironment in the depositional area. A hfe habitat unsuitable for the survival of foraminifers (? extremely hypo- saline) is supposed.
Comparison with foraminiferal assemblages from the whole sample (including fine and coarse beds) : oryctocenoses in both beds are very similar, there- fore, no significant differences between results ob- tained from separate beds and the whole sample were observed.
Subtype Ab : Reworked, suspension-transported foraminiferal tests (oryctocenosis 6) occur in both coarse and fine beds (Fig. 3).
This oryctocenosis was found in one Lower Ba- denian sample only. Tests are larger in coarser beds than in finer beds, differing from only suspension- transported foraminifers. Size sorting of tests pro- bably results from their reworking. Reworked fo- raminifers as well as other sedimentary particles are derived from the underlying Lower Miocene sediments, because index species of the Lower Mio- cene occur here. Species composition of assemblages from separate beds and the whole sample is similar. Oryctocenoses differ in a wider range of test size for the whole sample.
Type B : Indigenous or weakly transported species accompanied by reworked foraminiferal tests
Subtype Ba : Indigenous foraminifers (oryctoce- nosis 1) occur in coarse beds (sandstone), indigenous and reworked ones (oryctocenosis 5) in fine beds (siltstone or fine sandstone) (Fig. 4).
This type of distribution was observed in the Lower Badenian sediments. Indigenous assemblages contain foraminifers issued from a littoral, steno- haline environment: Asterigerinata planorbis, Dis- corbis discoides, Lobatula lobatula, Eponides um- bonatus, Pararotalia stellata, Elphidium div.sp., Ammonia ex gr. beccarii, Quinqueloculina div.sp., etc. Suspension-transported assemblages contain Lo- wer Miocene index species (Uvigerina graciliformis, Uvigerina bononiensis primiformis, Uvigerina par- keri breviformis). The species composition looks like that found in the "Rzehakia (Oncophora) Beds". The similarity index of the suspension-transported asemblages occurring in the Lower Badenian sedi- ment and assemblages from the "Rzehakia (Onco- phora) Beds" (sections MK 1 and MK 2) varies from 76 to 95 %. Therefore, the first transport of tests in suspension may be considered as occurring in the "'Rzehakia beds time interval".
During the Lower Badenian, small foraminifers were reworked from the "Rzehakia (Oncophora) Beds" and deposited probably from suspension. The absence of small, thin-walled suspension-transported foraminifers in the coarsest beds is probably related to a high energ~¢ environment, unsuitable for sedi- mentation of these small tests.
Indigenous foraminifers are 3 to 30 times more abundant in the coarse than in the fine beds. This fact may signify a lower sediment supply during the deposition of coarse beds.
Different species in assemblages from finer and coarser beds with different paleoecological require- ments and stratigraphical ranges lead to low values (45-78 %) of the similarity index (Fig. 6). Also the ranges of the diversity index and P/B-ratio for samples are wide (Fig. 6).
Comparison with foraminiferal assemblages from the whole sample (including fine and coarse beds) : (1) If foraminiferal abundance in a coarse bed is higher than the abundance in a fine bed, the size-distribution of tests from the whole sample may look like a polymodal one, and size-sorting of tests in suspension may be "hidden". In that case, as- semblages may be erroneously interpreted as deep- water with transported shallow-water species and a rare occurrence of the Lower Miocene index species (sample N-83/255 m). (2) For sample with similar foraminiferal abundance in fine and coarse beds, indigenous and suspension-transported tests are re- cognized also in whole sample (sample N-83/235 m). (3) If foraminiferal abundance in a coarse bed is lower than in a fine bed, indigenous foraminifers occur very rarely and only the peak for suspen- sion-transported foraminifers is visible on the size distribution curve (sample N-83/225 m). In some cases, indigenous foraminifers may not be found. This causes a misinterpretation of the paleoenviron- ment in the depositional area of suspension-trans- ported foraminifers.
Subtype Bb: Indigenous or weakly transported and reworked foraminiferal tests (oryctocenosis 5) are present in both finer beds and coarser fillings of bioturbation (Fig. 4).
This type of distribution was found only in the Lower Badenian sediments among samples with Ba type of test distribution. It is supposed that fora- miniferal assemblages distributed in these sediments were originally type Ba. Passage from type Ba to type Bb distribution could have been caused by bioturbation. The indexes of similarity are high (67, 77 %).
As expected, assemblages obtained from separate beds and the whole sample are similar.

Ba Bb
N-83/225
BENTHONIC FORAMINIFERA
BENTHONIC FORAMINIFERA
PLANKTONIC FORAMINIFERA
sandstone
ROUNDED
FORMS
ELONGATE FORMS
4o
=o
3°1
~t I
I~
~
N-83
/225
sa
ndst
one
o ..
..
..
..
o
'°0
/ sillstone
o,o9
0,
18
0,27
0,
36
0,45
0,
54
0,09
0,
18
0,27
0,
35
0,45
0,
54
0,09
0,
18
0,27
0,
38
0,45
0,
54
)
N-8
3/23
5 [:2
2] N
-83/
235
silts
tone
: ~ /~
20
lO
3
°~
40
] ~
20
80
1 °0
~
~ N
-83/
235
sand
ston
e
--
'-
-~
si
ltsto
ne
..
..
..
..
..
..
o~
8 o~
00
,09
0,18
0.
27
0,36
0,
45
0,54
0,
09
0.18
0,
27
0,36
0,
45
0,54
N-8
3/25
5
l/
no planktonic
fora
min
ifera
/
..
..
..
..
..
..
.
0 0~' ...............
silt
sto
ne
0,
09
o j8
0,
2~
0,38
0,
45
0,54
0,
09
o,~e
0,
27
0,35
0,
45
0,54
1Oo
~ .
..
.
/ Io°~ ~ ~
....
.. /
silts
fone
o.
o9
0.18
0,
27
0.36
0.
45
0,54
o.
og
0,18
0,
27
0.36
0,
45
0,54
~
(~
0,18
0.
27
0,36
0,
45
0.54
BE
NTH
ON
IC R
OU
ND
ED
FO
RM
S -
WH
OLE
SA
MP
LE (C
OA
RS
E +
FIN
E B
EDS)
0,1~1
N-8
3/22
5 M
"' 8
3/2
35
M
0,0g
0,
18
0,27
0,
36
0,45
0,
54
0,ca
o,18
o,21
o,~
o,,~
o,
~ I
Be
N'8
3/27
8 co
ng
lorn
e ro
l- e
san
dst
on
e
~ N
-95/
225
silts
tone
..
..
..
..
o
0,G
g 0,
18
0,27
0,
36
0,45
0,
54
0,O
g 0,
18
0,27
0,
36
0,46
0,
54
0,09
0,
18
0,27
0,
36
0,45
0,
54
30
30
20
1 10
..
..
..
..
..
..
..
.
0
0,0g
0,
18
0,27
0,
30
0,45
0,
54
o,o9
0,
18
0,27
0,
36
0,~5
0,
54 ~4
[Z
B
N-8
3/27
8 m
silt
ston
e 40
~
N-8
3/27
8 m
san
dsto
ne
30
20
lO
iP
/ o
......
0,09
0,
18
0,27
0,
36
0,45
0,
54
o,oe
o,
18
o,27
o,
a8
o,45
o,
~
ver'd
cal a
xis
- num
ber o
f spe
cim
ens
horiz
onta
l axi
s - t
est s
ize
(urn
)
FIG
. 4
. -
Siz
e d
istr
ibu
tio
n o
f b
enth
on
ic r
ou
nd
ed,
ben
tho
nic
elo
ng
ate
and
pla
nk
ton
ic f
ora
min
ifer
al t
ests
in
sam
ple
s N
-83
/22
5
m,
N-8
3/2
35
m
, N
-83
/24
5
m,
N-8
3/2
55
m
, N
-83
/27
8
m,
N-9
5/2
25
m
: t
yp
e B
wit
h i
nd
igen
ou
s o
r w
eak
ly t
ran
spo
rted
an
d r
ewo
rked
fo
ram
inif
eral
tes
ts.
Fo
r ex
pla
nat
ion
s of
lit
ho
log
ical
sch
emes
see
Fig
. 3
. L
eng
ht
of s
cale
bar
: 2
cm
. S
ub
typ
es B
a,
Bb
and
Bc
are
def
ined
in
tex
t.
Dis
trib
utio
n de
tai
lle
des
test
s de
fo
ram
ini~
res
bent
hiqu
es a
rron
dis,
be
nthi
ques
al
long
& e
t pl
anct
oniq
ues,
da
ns
les
~ch
anti
llon
s N
-83
/22
5
m,
N-8
3/2
35
m
, N
-83
/24
5
m,
N-8
3/2
55
m
, N
-83
/27
8
met
N
-95
/22
5
m :
typ
e B
, ~
test
s de
fo
ram
ini~
res
auto
chto
nes
ou f
aibl
emen
t tr
ansp
ort~
s ou
rem
ani~
s, v
oir
la F
igur
e 3
po
ur
les
l~ge
ndes
des
fig
ures
lit
holo
giqu
es.
Lon
gueu
r de
la
barr
e d'
~che
lle
: 2
cm.
Les
sou
s-ty
pes
Ba,
Bb
et B
c so
nt d
~fin
is d
ans
le t
exte
.
~9
©
©
C
mm
©
©

HOLC OVA 323
Subtype Bc: Indigenous foraminifers (oryctoce- nosis 1) occur in fine beds (fine sandstone), indi- genous and reworked ones (oryctocenosis 5) in coarse beds (fine conglomerate) (Fig. 4).
This type occurs only in one sample f rom the base of the Lower Badenian sedimentary cycles, and can reflect specific conditions in the beginning of transgression (rapid sedimentation). Species compo- sitions of foraminiferal assemblages belong to the Ba type, but their distribution in the laminae of sediments is reversed : reworked, suspension-trans- ported foraminiferal tests are present in the coarse bed, while indigenous foraminifers are present in the fine bed.
The size distribution curve for the whole sample does not correspond to the described type of size sorting of tests.
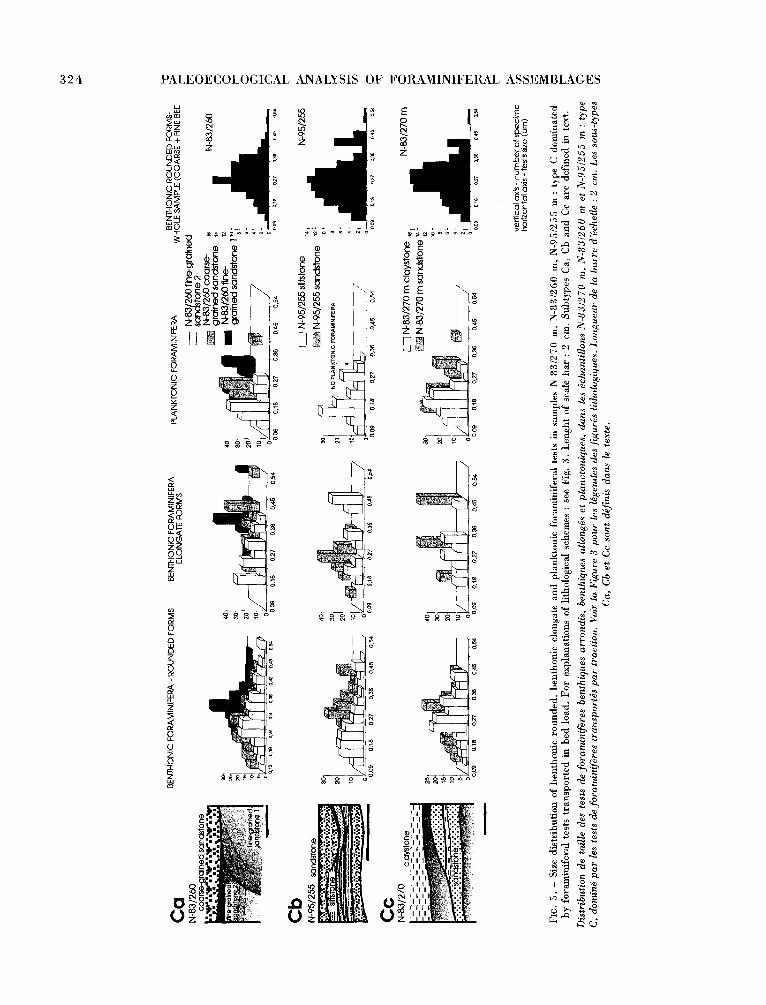
Type C: Tests transported in bed load prevailing in samples (Fig. 5)
The tests t ransported in bed load are always present in coarse beds but may also occur in fine beds. The sediments including these assemblages are the coarsest of all analysed samples (Fig. 6). Species compositions of indigenous and bedload-transported assemblages are similar (index of similarity varies between 74-88 %) with prevailing littoral, stenoha- line species : Asterigerinata planorbis, Discorbis dis- coides, Lobatula lobatula, Eponides umbonatus, Pa- rarotalia steUata, Elphidium div. sp., Ammonia ex gr. beccarii, Quinqueloculina div. sp., etc. This similarity shows that foraminiferal tests were trans- ported in bed load only along a short distance in the same type of paleoenvironment. Tests t ranspor- ted in bed load are often abraded and rarely broken. All samples with this type of distribution come from the Lower Badenian unit.
The following subtypes of type C were distinguis- hed (Fig. 5). All subtypes are represented only by one sample :
Subtype Ca : Tests t ransported in bed load (oryc- tocenosis 3) are present in coarse bed (coarse sandstone), while indigenous foraminifers or weakly t ransported ones (oryctocenosis 1) occur in fine laminae (fine sandstone).
Subtype Cb : Tests t ransported in bed load (oryc- tocenosis 3) are found in coarse beds (sandstone), while indigenous Lower Badenian foraminifers and reworked suspension-transported Lower Miocene fo- raminifers (oryctocenosis 5) occur in fine beds (silts- tone). Lower values of the similarity index are due to the occurrence of different reworked species.
In the whole sample, mixing of corroded shal- low-water species and well-preserved deep-water species may lead to an interpretation of a deep-wa- ter environment with bedload-transported shallow- water species.
Subtype Cc : Tests t ransported in bedload (oryc- tocenosis 3) are observed in coarse beds (coarse sandstone), while indigenous and bedload-transpor- ted foraminifers (oryctocenosis 4) occur in fine beds (fine sandstone).
Assemblages from the whole sample of types Ca and Cb contain indigenous and bedload-transported assemblages, and, generally, the paleoenvironmental interpretation of assemblages f rom the whole sam- ples and from the separate beds is similar. Analysis of separate beds gives complementary information about accumulation of bedload-transported tests in coarse beds (probably lenses accumulated in troughs of wave ripples).
P o s i t i o n o f t ypes A-C in cyc le o f s ea - l eve l c h a n g e s
Cycles of sea-level changes were distinguished in analysed sections from the cyclical changes in fora- miniferal assemblages (Holcov~, 1996b). The pre- sence of types A, B and C distributions of foraminiferal tests in beds was summarized for these cycles. The presented results are based on a small number of samples from only a shallow-marine environment. The set analysed is too small to draw significant conclusions, but it can provide some useful information :
Lower Badenian samples (Fig. 7) The Lower Badenian transgressive layer is cha-
racterized by two distinctive subtypes of oryctoce- nosis distr ibution: (1) in the nearshore area only reworked tests occur in both beds (type Ab), (2) in the more central par t of the basin, small, sus- pension-transported and reworked tests occur in coarse beds (type Bc). I t may be caused by a rapid sedimentation of coarse beds which enabled the preservation of small tests. These subtypes are ab- sent from other parts of sea-level change cycles.
Tests t ransported in bed load without reworked species (types Ca, Cc) are typical of the middle and upper parts of transgressive systems tracts.
Reworked species, deposited probably in finer beds from suspension (type Ba) appear in highstand systems tracts and characterize also lowstand sys- tems tracts. In highstand systems tracts, the species reworked into coarser beds may be accompanied by tests t ransported in bed load (type Cb). Type Ba with indigenous or weakly t ransported tests in
Ccl
N
-83
/26
0
Cb
1%
95/2
55
sand
ston
e
one
• ,,.'.'
.,,.,';
,.,.,';
.
Cc
N-8
3/2
70
cl
ovst
one
BE
NTH
ON
IC F
OR
AM
INIF
ER
A -
RO
UN
DE
D F
OR
MS
B
EN
THO
NIC
FO
F4AM
INIF
ERA
ELO
NG
ATE
FO
RM
S
..
..
.
. 0
~-
,
, ,
, ,
, ~
012
018
O24
03
O~
04~
O~
O54
0,09
0,
18
O,27
0,
36
0,45
0,
54
30
40
~
20
0 /
, ,
..
..
..
~
e 0
0,09
0,
18
0,27
0,
36
0,45
0,
54
0,09
5,~8
0,27
0,
35
0AS
0,54
0.09
0.
18
0,27
0,
36
O,4.5
0,
54
0,0g
0,
15
0,27
0,
88
0,45
0,
54
PLA
NK
TON
IC F
OR
AM
INIF
ER
A
WH
OL
N-8
3/26
0 fi
ne
-gra
ine
d
~a
nd
sto
ne
2
N-8
3/26
0 co
ars
e-
~ T
gra
ine
d s
ands
tone
40
, ~
-~
I
N-8
3/2
60
fin
e-
0,09
0,
18
0,27
0,
36
0,45
Q
,54
o,0~
BE
NTH
ON
IC R
OU
ND
ED
FO
RM
S-
WH
OLE
SA
MP
LE (C
OA
RS
E +
FIN
E B
EE
1'4-
83/2
60
0,18
0,2
7 0,3
s o,4
s o,~
,
N-9
5/25
5 sU
tsto
ne
,,
30
~ ~
N-9
5/25
5 sa
nd
sto
ne
ii
21
!~
~I
NI
FE
R
~N
O
PLANKT
ONIC
FORA
MIN
IFERA
q;I t
0,09
0,
18
0,27
0,
36
0,45
0,
54
O,OS
N-9
5/25
5
N-8
3/2
70
m c
lays
ton
e
~o
~ ~
1%83
/270
m s
an
dst
on
e ~
0,09
0,
15
0,27
0,
36
0r45
0,
54
O,OB
NL8
3/27
0 m
0,18
0,27
Q,36
0,4
5 0,5
4
veR
ical
axi
s -
num
ber
of s
peei
me
horiz
onta
l axi
s - t
ests
siz
e (u
m)
FIG
. 5.
-
Siz
e d
istr
ibu
tio
n o
f b
enth
on
ic r
ou
nd
ed,
ben
tho
nic
elo
ngat
e an
d p
lan
kto
nic
fo
ram
inif
eral
tes
ts i
n sa
mpl
es N
-83
/27
0
m,
N-8
3/2
60
m
, N
-95
/25
5
m :
typ
e C
do
min
ated
b
y f
ora
min
ifer
al t
ests
tra
nsp
ort
ed
in b
ed l
oad.
F
or
exp
lan
atio
ns
of l
itho
logi
cal
sche
mes
: s
ee F
ig.
3.
Len
gh
t of
sca
le b
ar :
2
cm.
Su
bty
pes
Ca,
C
b an
d
Cc
are
defi
ned
in t
ext.
Dis
trib
utio
n de
tai
lle
des
test
s de
fo
ram
inif
~re
s be
nthi
ques
a
rro
nd
is,
bent
hiqu
es
allo
ng6s
et
pla
ucto
niqu
es,
clan
s le
s ~
chan
till
ons
N-8
3/2
70
m
, N
-83
/26
0
m e
t N
-95
/25
5
m :
typ
e C
, do
min
d p
ar
les
test
s de
fo
ram
ini~
res
tran
spor
t$s
pa
r tr
acti
on.
Voi
r la
Fig
ure
3 p
ou
r le
s l~
gend
es
des
fig
ur$
s li
thol
ogiq
ues.
L
on
gu
eur
de l
a ba
rre
d'dc
hell
e :
2 cm
. L
es s
ous-
type
s C
a, C
b et
Cc
sont
d~f
inis
d
an
s le
tex
te.
©
©
©
> .5 5/3
C
©
C/2
L~
HOLC OVA 325
8O
z 70 O 60
(N b=, ~o
.o
~ 20
0 0
45
40 LU ~. 35
~ 3o ~, 25 _. 2O
n,- , 10
II)
0
PERCENTAGE OF S A N D F R A C T I O N IN SEDIMENT
o o
o
=• D
a •
i i i i i i
10 20 30 40 50 60
%SAND FRACTION - SAMPLE 1
PERCENTAGE OF PLANKTONIC FORAMINIFERA
o o
10 20 30 4 0
P/B-RATIO - SAMPLE 1
50
• A a + A b
B a + B c
Bb
C
ii A a + A b
Ba + Bc
• B b
C
14 t~ u.= 12
: E l 0 < o~ 8
~" 4
-- 2 a 0
DIVERSITY OF BENTHONIC FORAMINIFERA
o
o o
o
• n o
5 10
DIVERSITY - SAMPLE 1
FORAMINIFERAL NUMBER (BENTHONIC FORAMINIFERA)
300 - uJ
o :~ 250 -
z r~ 200 - .,JLU
,,,=$ 150- . , , < • o
~o loo o ° ° m~--.m m
< 50 • ~ ~ n- O • •
0 50 100 150 200
FORAMINIFERAL NUMBER - SAMPLE 1
250
• A a + A b
Ba + Bb
Bb
C
i
15
• A a + A b
" Ba+Bc
Bb
C
SIMILARITY OF FORAMINIFERAL ASSEMBLAGES
• 8ot c 70 +
~ 40 t
: E 1 0 ~
o~ 0
I $ o
II 8 o BC
o c
Ba
Aa
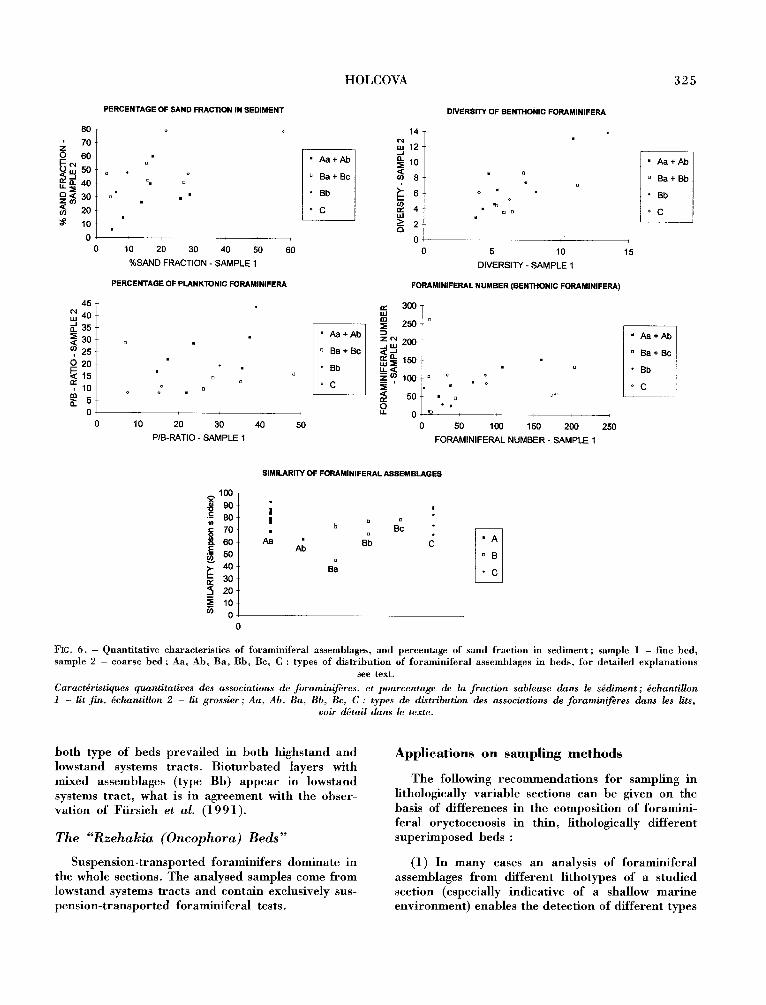
FIG. 6. - Quan t i t a t i ve charac te r i s t i cs of fo ramin i fe ra l assemblages , and percen tage of s and f rac t ion in s e d i m e n t ; sample 1 - fine bed, sample 2 - coa r se bed ; Aa, Ab, Ba , Bb, Bc, C : types of d i s t r i b u t i o n of f o r a m i n i f e r a l a s semblages in beds , for d e t a i l e d e x p l a n a t i o n s
see text.
Caract~ristiques quantitatives des associations de foraminif~res, et pourcentage de la f rac t ion sableuse dans le s~diment ; ~chantillon 1 - lit rift, ~chantillon 2 - lit grossier ; Aa, Ab, Ba, Bb, Bc, C : types de distribution des associations de foraminif~res dans les lits,
voir d~tail dans le texte.
both type of beds prevailed in both highstand and lowstand systems tracts. Bioturbated layers with mixed assemblages (type Bb) appear in lowstand systems tract, what is in agreement with the obser- vation of Fiirsich et al. (1991).
The "Rzehakia (Oncophora) Beds"
Suspension-transported foraminifers dominate in the whole sections. The analysed samples come from lowstand systems tracts and contain exclusively sus- pension-transported foraminiferal tests.
Applications on sampling methods
The following recommendations for sampling in lithologically variable sections can be given on the basis of differences in the composition of foramini- feral oryctocenosis in thin, lithologically different superimposed beds :
(1) In many cases an analysis of foraminiferal assemblages from different lithotypes of a studied section (especially indicative of a shallow marine environment) enables the detection of different types

326 P A L E O E C O L O G I C A L ANALYSIS OF F O R A M I N I F E R A L ASSEMBLAGES
LOWER PART OF TRANSGRESSIVE SYSTEMS TRACTS
, ~ N-48/95 rn N-83/278 m
Aa
MIDDLE AND UPPER PART OF 1RANSGRESSIVE SYSTEMS TRACTS
\ N-83/260 m N-83/270 m
HIGH STAND SYSTEMS TRACTS
N-83/235 m Aa N-83/255 m N-95/255 m
LOW STAND SYSTEMS TRACTS
Aa N-83/245m
N-83/225m N-95/225m
Bb
1
- - " ~ 7 " a ' ~ 8 * 9 ~"'.-,,,10
FIG. 7. - Model of distribution of types A, B, C (for detailed explanation see text) in the Lower Badenian cycles of sea-level changes, l - sandstone, 2 - siltstone, 3 - claystone, 4 - graded bedding, 5 - siltstone with undulating lamination and sandstone lens accumulated in a trough of wave ripples, 6 - coarse filling of ichnofossil in laminated siltstone, 7 - resedimentation of foraminiferal tests, 8 - bedload transport of forami~fiferal tests, 9 - occurrence of indigenous foraminiferal tests, 10 - mixing of
foraminiferal assemblages by bioturbation. ModUle de d i s t r ibu t ion des types A , B, C (vo ir d~tail dans le tex te) dans les cycles de var ia t ion d u n iveau m a r i n au Bad~n ien inf~rieur. 1 - gr~s, 2 - s i l ts tone, 3 - argilol i the, 4 - granoc las - s emen t , 5 - s ihs tone & lamina t ion ondul~e et lentille gr~seuse accumul~e dans u n c reux de r ide de houle, 6 - rempl issage gross ier d 'une t race foss i le dans un si l ts tone lamin~, 7 - res~- d imen ta t i on de tests de f o r a m i n i ~ r e s , 8 - t ranspor t p a r t rac t ion de tests de f o r a m i n i f ~ r e s , 9 - p r d s e n c e de tests de f o r n m i n i ~ r e s a u t o c h t o n e s , 10 - m ~ l a n g e d 'a s soc ia t ions de f o r a m i n i f ~ r e s p a r
b io turba t ion .
of assemblages on the basis of the degree of post- mor t em changes. The presence of t r a n s p o r t e d and r eworked tests p rov ides add i t iona l da t a abou t the bas in h i s to ry (e.g. su spens ion - t r anspo r t ed assem- blages - type A - give in fo rmat ion about the cen t ra l p a r t of bas in and its d y n a m i c s ; tests t r a n s p o r t e d in bed load - type C - about marg ina l p a r t ; r eworked species - types A b and B - abou t the
occu r rence of o lder sediments and condi t ions sui~ tab le for the i r r ework ing and r e sed imen ta t ion of par t ic les) .
(2) The finest beds are genera l ly cons ide red as the more f avourab le for micropa leonto log ica l ana- lyses and are often sampled preferent ia l ly . Our s tudy shows tha t assemblages wi thout ev ident post- mor t em changes can occur in the coarses t beds while r e w o r k e d tests a re found in the finest beds ( type Ba).
(3) Strongly b i o t u r b a t e d layers conta in mixed fo ramin i fe ra l assemblages ( type Bb) , so a de ta i l ed sampl ing of these layers p roduces mis leading da ta .
CONCLUSIONS
Var iab ih ty in the composi t ion of fo ramin i f e r a l assemblages (more exact ly oryctocenoses) f rom dif- fe ren t hthologies were s tudied in 12 core samples and 2 ou tc rops f rom the Ot tnangian (Middle Bur - digal ian) and Lower B a d e n i a n (Langhian) sediments of the South Slovak Basin inc luded in the Cen t ra l P a r a t e t h y s .
Based on size sor t ing of tests , the i r p r e s e r v a t i o n and the presence /absence of species with d i f ferent paleoecological r equ i remen t s and s t r a t i g raph ica l ranges in assemblages, six types of fo ramin i f e r a l oryctocenosis were d is t inguished (Tab. 1), each of them conta in ing indigenous , su spens ion - t r anspo r t ed tests , b e d l o a d - t r a n s p o r t e d and /o r r e w o r k e d tests. These types of oryctocenoses may be d i s t r i bu t ed in couples of supe r imposed l i thological ly d i f fe ren t beds . The following cases may occur :
(1) Only suspens ion - t r anspo r t ed fo ramin i fe ra l tests occur in the bo th beds. The tests be long to ner i t ic and u p p e r b a t h y a l species, and cha rac t e r i ze the source a rea by ma te r i a l t r a n s p o r t e d in suspen- sion by s to rm waves. A coasta l , hyposa l ine envi ron- men t unsu i tab le for the surv iva l of fo ramin i fe r s is p r o p o s e d for the depos i t iona l a rea . These oryc to- cenoses may occur in the whole cycle of sea-level change.
(2) Sed imen t s wi th p r e v a i l i n g u n s o r t e d , well- p r e s e r v e d , inner-shelf , indigenous or weak ly t r ans - p o r t e d tests may conta in r eworked , su spens ion - t r anspb r t ed , outer -shel f to b a t h y a l fo ra - minifers . The l a t t e r or iginate f rom the under ly ing sediments , fully e r o d e d in the ana lysed area . Re- worked tests occur in the t ransgress ive layer , while

HOLC OVA 327
in highstand and lowstand systems tracts they ap- pear in finer beds probably deposited from suspen- sion. Bioturbations occurring in lowstand systems tracts cause mixing of reworked and indigenous assemblages.
Size-distributions of tests from some whole sam- ples (including coarse and fine beds) are polymodal without a peak for suspension-transported forami- nifers. In that case, a deep-water paleoenvironment with transported shallow-water species and a rare occurrence of the reworked Lower Miocene forami- nifers may be erroneously interpreted.
(3) The coarsest beds (in some cases lenses ac- cumulated in troughs of wave ripples) contain lit- toral stenohaline foraminifers sorted by bed-load transport. Unsorted, indigenous or weakly-transpor- ted tests prevail in the finest beds. Sediments were deposited in a coastal marine stenohaline environ- ment affected by wave action. They were observed in the middle and upper parts of transgressive systems tracts, and, together with reworked outer- shelf to bathyal foraminffers, also in highstand systems tracts.
An analysis of assemblages from the whole sample may cause a misinterpretation of a deep-water en- vironment with an occurrence of bedload-transpor- ted shallow-water species and a rare occurrence of the Lower Miocene index species.
(4) Only reworked tests and similar assemblages in both beds were observed at the base of a trans- gressive systems tract.
Thus, detailed sampling of all lithotypes and a taphonomical analysis of assemblages is able to provide new paleoenvironmental information. Sam- pling of a unique lithotype (e.g. claystone) in sec- tions may yield incomplete data.
ACKNOWLEDGMENTS
The author thanks reviewers for their comments and suggestions.
This study is a part of research project No. 201/95 supported by Charles University, Prague.
BIBLIOGRAPHY
BOLTOVSKOY E. and WRIGtlT R. ( 1 9 7 6 ) : Recent Foraminifera. Junk , The Hague, p. 1-515.
CICHA I., ZAPLETALOVA I,, MOLCiKOVA V. and BRZOBOHAT'~ R. (1983) : Stratigraphical range of Eggenburgian-Badenian Fo- ranlinifera in West Carpathian Basins. Knih. Zem. Plyn Nafta, Hodonin, vol. 4, p. 99-144 .
CULVER S.J. and BANNER F.F. (1978) : Foraminifera1 assemblages as Flandrian paleoenviroumental indicators. Palaeogeogr., Palaeoclimatol., Palaeoecol., Amsterdam, vol. 24 , p. 53-72.
~TYROK't P. (1968) : The correlation of Rzehakia (Oncophora) series in Eurasia. Palaeogeogr., Palaeoclimatol., Palaeoecol., Amsterdam, vol. 4, p. 2 5 7 - 2 7 0 .
FOLK R.L. and WARD W.C. (1957) : Brazos River Bar : a study in the significance of grain size-parameters. Journ. Sediment. Petrology, Tulsa, vol. 27 , p. 3-26.
FRIEDMAN G.M. (1979) : Differences in size distribution of po- pulations of particles among sands of various origins. Sedi- mentology, Amsterdam, vol. 26 , p. 3-32.
Ft~Rs~cn F.F., OSCHMANN W. and JA1THY A.N. ( 1 9 9 1 ) : Faunal response to transgressive-regressive cycles : example from the Jurassic of Western India. Palaeogeogr., Palaeoclimatol., Palaeoecol., Amsterdam, vol. 85 , p. 149-159 .
HAMOR G. (1985) : Geology of the N6grad-Cserhat area. Geolo- giea Hungarica, Budapest , wil. 22 , p. 1-307.
HOLCOVJ~ K. (1996a) : Determination of t ransport of foramini- feral tests in the fossil record (the South Slovakia Basin, Middle Miocene). Neues Jb. Geol. Paleont., Mh., Sttutgart, vol. 4, p. 193-217 .
HOLCOV$ K. ( 1996b ) : Cykhck6~zmeny foraminiferov)~ch spo- lo~enstiev : nov6 hdaje pre sekven~n6 stratigrafin ? Sedimen- tdrni geologie v ~'esk~ republice, 96 , Praha , p. 15.
]:IOLCOVA-SUTOVSKA K. ( 1 9 9 6 ) : Foraminiferal assemblages: in- dicator of paleoenvironmental evolution of marine basins and eustatic changes (Kiscellian-Karpatian of the South Slovakia and Danube basins). Geologica Carpathica, Bratislava, vol. 47 (2), p. 119-130 .
HoLcovX K., DOIAKOVX N., VASS D., ZAGOR~EK K. and ZELENKA J. ( 1 9 9 6 ) : Foraminifera, Bryozoa, Ostracoda and palyno- morphs like indicators of marine environment in the Lower Badenian of Strh~re-Tren~ graben (South Slovakia Basin). Mineralia Slovaca, Bratislava, vol. 28 , p. 99-119 .
KovX~ M., C[CRA I., KRYSTEK I., SLACZKA A., STRANUK Z., OSZCZYPKO N. and VASS D. (1989) : Palinspastic maps of the Western Carpathians Neogene. Geological Survey (Praha), p. 1-35.
KVALIASHVILI G.A. ( 1 9 6 2 ) : Onkoforovyj (rzehakiovyj) gorizont Eurazii. Akadem. Nauk Gruzinskoj SSR, Tbilisi, p. 1-222.
LOOSE T.L. (1970) : Turhulent t ransport of benthonic foramini- fera. Contt: Cushman Found. Foram. Res., Ithaca N.Y., vol. 21 , p. 164-166 .
McCAVE I.N. ( 1 9 7 8 ) : Grain size t rends and t ranspor t along beaches : example from Eastern England. Marine Geology, Amsterdam, vol. 28 , p. 43 -51 .
McLAREN P. (1982 ) : Hydraulic control of grain-size distributions in a macrotidal estuary - discussion. Sedimentology, Amster- dam, vol. 29 , p. 4 3 7 - 4 3 9 .
McLAREN P. and BOWLES D. ( 1 9 8 5 ) : The effects of sediment t ransport on grain-size distributions. Journ. Sediment. Pe- trology, Tulsa, vol. 55(4), p. 457=470.
MIDDLETON G.V. (1976) : Hydraulic interpretation of sand dis- tribution. Journ. Geology, Chicago, vol. 84 , p. 405 -426 .
MURRAY J.W. (1965) : Significance of benthic foraminiferids in plankton samples. Journ. Paleont., Tulsa, vol. 39 , p. 56- 157.
MURRAY J.W. (1973) : Distribution and ecology of living benthic foraminiferids. Heinemann educ. books, London, p. 1-274.
MURRAY J.W. ( 1 9 9 1 ) : Ecology and paleoecology of benthic Foraminifera. Longman Scientific and Technical, London, p. 1-397.

328 PALEOECOLOGICAL ANALYSIS OF FORAMINIFERAL ASSEMBLAGES
MURRAY J.W., STURROCK S. and WESTON J. (1982) : Suspended load transport of foraminiferal tests in a tide- and wave-swept sea. Journ. Foram. Res., Washington, vol. 12 (1), p. 51-65.
PAPP A., BOGL F. et SENE~ J. (1973) : Chronostratigraphie und Neostratotypen 3. Ottnangien. Vydav. Slov. Akad. Vied, Bra- tislava, p. 1-841.
PHLEGER F.B. (1965) : Ecology and distribution of Recent Fo- raminiferds. John Hopkins Press, Baltimore, p. 1-289.
REISS Z. and HOTTINGER Z. ( 1 9 8 4 ) : The Gulf of Aqaba - ecological micropaleontology. Springer Verlag, Berlin-Heidel- berg-New York-Tokyo, p. 1-354.
ROGL F. (1986) : Late Oligocene and Miocene Planktic Forami- nifera of the Central Parate thys . In Plankton stratigraphy, edited by Bolli H.M., Saunders J.B. and Perch-Nielsen K. Cambridge Univ. Press, p. 3 1 5 - 3 2 8 .
ROGL F. and STEININGER F.F. (1983) : Vom Zerfall der Tethys zu Mediterran und Paratethys. - Die neogene Pal~ogeographie and Palinspastik des z i rkummedi terranen Raumes. Ann. Na- turhist. Mus., Wien, vol. 85/A, p. 135-164 .
SINGER J.K. et ANDERSON J.B. (1984) : Use of total grain-size distributions to define bed erosion and t ranspor t for poorly
sorted sediment undergoing simulated bioturbation. Marine Geology, Amsterdam, vol. 57, p. 335 -359 .
UDDEN J.A. (1914) : Mechanical composition of clastic sediments. Geol. Soc. Amer. Bull., Tulsa, vol. 25 , p. 655 -744 .
VASS D. (1995) : Global sea level changes reflected on Nor thern margin of the Hungar ian Paleogene the Fiakovo and Novoh- rad (Nograd) Lower Miocene Basin (South Slovakia). Mine- ralia slovaca, Bratislava, vol. 27 , p. 193-206 (in slovak).
VASS D., KONE~N~" V. and SEFARA J. ( 1 9 7 9 ) : Geology of the Ipel'sk~ kothna Basin and file Krupinsk~ planina Upland. Geol. Ust. D. ~tftra, Bratislava, p. 1-227 (in slovak).
VASS D. (ed.) ( 1992 ) : Explanations to geological map of the Lu~eneck~ kotlina Basin and the Cerovh vrchovina Upland 1 : 5 0 0 0 0 . Geol. (/st. D. ~tftra, Bratislava, p. 1-196 (in slovak).
VISHER G.S. ( 1 9 6 9 ) : Grain size distributions and depositional processes. Journ. Sediment. Petrology, Tulsa, vol. 39 , p. 1047 -1106 .
WANG P. and MURRAY J.W. (1983) : The use of Foraminifera as indicators of tidal effects in estuarine deposits. Mar. Micropaleont., Amsterdam, vol. 51 , p. 2 3 9 - 2 5 0 .
PLATE 1
Studied outcrops and core samples (sd = sandstone, st = siltstone).
1. Section MK 1, samples 1-4, Mal~ Krti~ sand pit, "Rzehakia (Oncophora) Beds". Massive sandstone (1) at the base, laminated sandstone with slump structure (2) above. Lami- nated sandstone (3) with thin beds of shell detritus (4) in the upper par t of the section (tempestites).
2. Section MK 2, samples 1 4 , Mal)~ Krti~ sand pit, "Rzehakia (Oncophora) Beds". Massive sandstone (1) overlain by la- minated sandstone (2) with pebbles of massive sandstone (3). Cross bedding (4) is observed in the upper part .
3. Core N-96, 305 m, "Rzehakia (Oncophora) Beds". Thin bed of sihstone (st) in massive sandstone (sd). Coin diame- ter : 2 cm.
4. Core N-83, 235 m, Lower Badeuian. Sihstone with undu- lating lamination (st), a sandstone lens (sd) is accumulated in a trough of a wave ripple. Lenght of scale bar : 2 cm.
5. Core N - 8 3 , 2 7 8 m, Lower Badenian. Fine-grained sandstone with undulat ing lamination (st), conglomerate (sd) on ero- sional surface. Bioturbations occur, fillings of bioturbations were not sampled.
6. Core N - 9 5 , 2 5 5 m, Lower Badenian. Laminae to thin beds of siltstone and fine-grained sandstone (st) overlain by coarse-grained sandstone (sd).
7. Core N-95, 225 m, Lower Badenian. Coarse filling of ichnofossil (sd) in laminated siltstone (st).
Echantillons $tudi~s d'affleurements et de carottes (sd = gr~s, st = siltstone).
1. Coupe MK 1, ~chantillons 1-4, Mal~ Krti~ sand pit , ~ Bancs it Rzehakia (Oncophora) ~,. Gr$s massi f h la base (1), gr~s laminas it slump au-dessus (2). Gr$s lamin~ (3) h miuces lits de d~bris coquilliers (4) it la partie sup~rieure de la coupe (tempestites).
2. Coupe MK 2, ~chantilloas 1-4, Mal~ Krti~, ~, Bancs it Rzehakia (Oncophora) ,,. Gr$s massi f (1) surmont6 par des gr$s laminas (2) it galets de gr$s massi f (3). Des d~p6ts entrecrois~s (4) s'observent it la partie sup~rieure.
3. Carotte N-96, 305 m, ~ Bancs it Rzehakia (Oncophora) ,,. Mince lit de siltstone (st) dans du gr$s massi f (sd). Diam$tre de la piece : 2 cm.
4. Carotte N-83, 235 m, Bad~nien inf~rieur. Sihstone it la- mination ondul~e (st), une lentille gr~seuse (sd) est accu- mul~e dans un creux de ride de houle. Longueur de la barre d'$chelle : 2 cm.
5. Carotte N-83, 278 m, Bad~nien inf~rieur. Gr$s f i n it la- mination ondul~e (st), conglom~rat (sd) sur une surface d'~rosion. Presence de bioturbations, dont les remplissages n'out pas ~t~ ~chantillonn~s.
6. Carotte N-95, 255 m, Bad$nien iuf~rieur. Lamines ou minces lits de siltstone et gr$s f ins (st) surmont~s de gr$s grossiers (sd).
7. Carotte N - 9 5 , 2 2 5 m, Bad~nien inf6rieur. Remplissage gros- sier d'une trace fossile (sd) darts un siltstone lamin~ (st).

HOLCOVA. PLANCHE
REVUE DE MICROPALI~ONTOLOGIE, VOL. 40, N ° 4





























