Embed Size (px)
Citation preview

Phyrochemurry. 1976. Vol. 15, pp. 189%18%. Pngamon Rar. Rinted m England
COMPOSITION LIPIDIQUE DES RACINES DE DIVERSES ESPECES CULTIVEES
MICHEL ROSSIGNOL* Centre de Recherches Agronomiques du Midi C.E.P.E L.Emberger et R.C.P. no246 du C.N.R.S.
(Recked received 7 Jute 1976)
Key Word Index-Angiospermae; Gymnospermae; roots; phospholipids; fatty acids.
Abstract-fatty acids containing 10 to 22 carbons have been found in the whole root of 20 plant species; Cl6 and Cl8 acids are the more abundant ones. All common phospholipids are present; the most frequent are phosphati- dylcholine and phosphatidylethanolamine. A relationship is suggested between the complexity of the lipid composi- tion and phylogeny. Species which exhibit very poor tolerance to high-lime soils seem to contain more saturated fatty acids than species which do not become chlorotic under the same conditions.
INTRODUCHON
Alors que de trks nombreuses don&es ont ktk publikes sur les lipides des tissus photosynthktiques, les lipides racinaires n’ont fait l’objet que de peu d’ktudes; leur nature, en particulier en ce qui conceme les phospholi- pides et les acides gras, est peu connue. A c&C de quel- ques don&es g&&ales [1,2], on peut rappeler que Lepage [3], sur les racines de Br~ssicu napus, et Roughan et EIatt [4]. sur celles de PastiMcQ sotioa, indiquent que les phospholipides majeurs de ces racines sont la phos- phatidylcholine et la phosphatidykthanolamine. Bartho- lomew et Mace[5] notent une prksence importante de phosphatidylinositol dans les racines de Phuseolus limensis. Par contre Keenan et al. [6] observent des teneurs importantes en acide phosphatidique dans les racines d’Auena satil;Q. Keenan et al. et Holman et Nichols [n, sur les racks de diverses Orchid&, ont trouvk que l’acide linoleique est quantitativement le plus important. Par contre Lepage indique que l’acide linok- nique est majoritaire dans les racks de Brassica napus.
De nombreux travaux ont post& une intervention des lipides dans l’adaptation des racks au milieu. Fergu- son [8], puis Kuiper et al. [9] ont montrC que la teneur en phosphatidylcholine des racines de diverses Gra- minks dkpend de la composition de la solution nutritive. Oursel et ol. [lo], puis Lamant et Heller [ 1 l] ont observe une relation entre la capacitk de fixation passive du cal- cium par les racks de deux Lkgumineuses et leur teneur en phospholipides acids. On a kgalement souvent note des diffkrences de composition en phospholipides et en acides gras entre les racks de variktks d’une m&me esp&ce diffkremment sensibles au froid [12,13].
Dans le cadre d’un programme de recherches sur la mkcanismes &adaptation aux sols calcaires, nous nous sommes attach& & l’ttude de la composition en phospho- lipides et en acides gras des racks de diverses espks
* Adresse actuelle: Laboratoire d’Ecologie de la Nutrition Minbale du Centre L. Emberger, BP 5051.34033 Montpellier- Cedex, France.
cultivkes, annuelles ou pkrennes, choisies en fonction de leur caractkre calcicole ou calcifuge.
RESULTATS ET DISCUSSION
Ces rksultats ont ktk obterius sur des racks entitres et correspondent aux valeurs moyennes de 5 k 10 rkp-kti- tions.
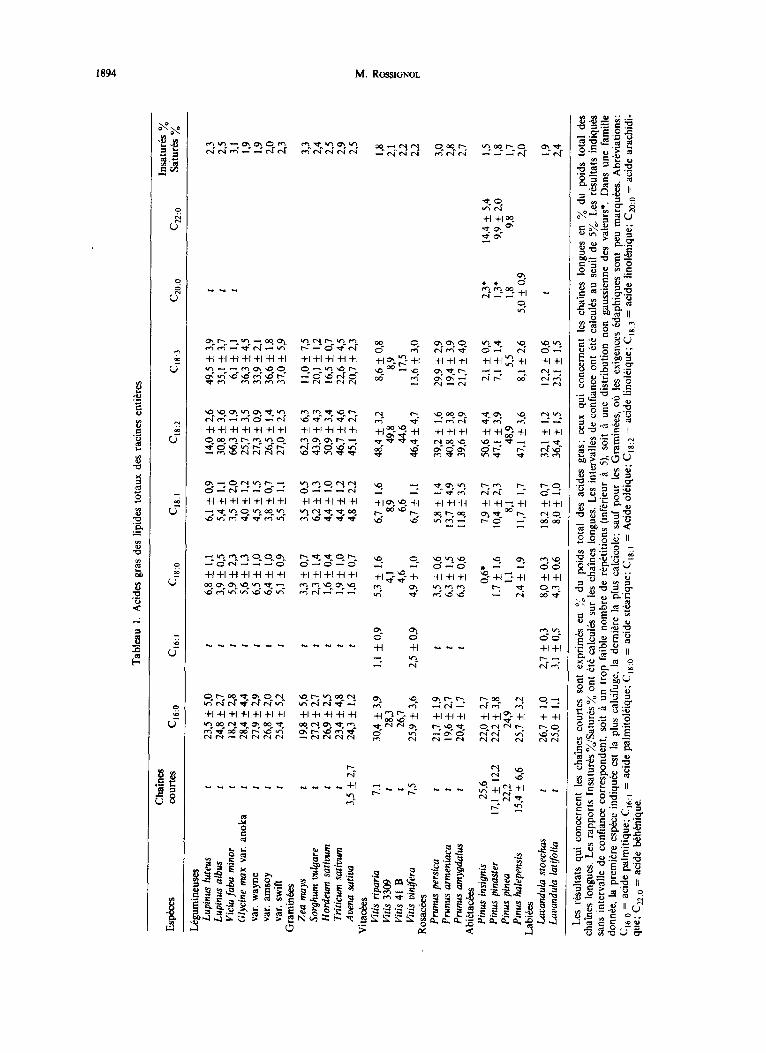
Acides gras. Les acides gras racinaires des espkces ktudikes sont en gtnbal B chaines longues (16 et 18 atomes de carbone), sauf chez les Abi&ackes qui contien- nent des proportions importantes d’acides gras a cha^mes courtes (10 ?I 14 atomes de carbone) ou ti chaines trb longues (20 ou 22 atomes de carbone) (tableau 1). Dans toutes les families il y a davantage d’acides gras insaturb que d’acides gras saturb (1,5 g 3 fois plus). Les acides gras saturb sont constituks essentiellement d’acides pal- mitique et, dans une moindre mesure, stkarique; les acides gras insaturts, d’acides linokique et linoknique; les monoinsaturks &ant peu reprkntb.
L’acide linolkique est le plus souvent majoritaire, sauf chez les Lkgumineuses oh l’acide linolknique est en gtntral le plus important. Mais il ne s’agit probablement pas +ne caractkristique de cette famille; en effect Vicia J&Q manifeste des tendances inverses, tout comme Medi- CQgO satica [ 133.
Les Abiktackes prksentent des compositions plus varites que les autres familles; nous avons trouvk des rtsultats similaires chez une Mimosa& (ACQC~I mdis- sima). Kaimal et Lakshminarayama [14] ont kgalement notC des tendances analogues cha une Malvacke (Ceiba pentandra) et une Sterculiacke (Stercukafoetida). &‘I peut remarquer que ces familles, apparues ZI 1’Qe secondaire, sont peu Cvolukes d’un point de vue phylogknique; la autres familles que nous avons &udiks sont apparues plus rkcemment, B I’tre tertiaire. On constate que la com- position en acides gras de leurs racks est qualitative- ment trb homogtie, puisqu’elles ne comportent presque exclusivement que des acides gras g 16 et 18 carbones. Nous n’avons pas ktudik un nombre suffisamment impor- tant de families pour qu’il soit possible de tirer des con-
1893
E P
Tab
leau
1.
Aci
des
gras
de
s lip
ides
to
taux
de
s ra
cine
s en
tiere
s
Cha
ines
C
oUrt
es
cl6:
O
‘I,:,
Cl,:
0 C
18.1
‘I,
:2
%3
c2o:
o %
:O
Insa
turb
“/
_
.._A
Si
tu&
s %
L&
umin
euse
s Lu
pinu
s lu
teus
t
Lupi
nus
albu
s t
Vici
a fa
bu
min
or
r G
lyci
ne
mux
va
r. an
oka
t va
r. W
ayne
va
r. am
soy
var.
swif
t G
ram
in&
s Z
ea
may
s So
rghu
m
m&a
re
Hor
detm
r sa
tivum
T
ritic
um
sntiv
wn
Aven
a sa
tiva
Vita
&es
V
itis
ripa
ria
Viti
s 33
09
Viti
s 41
B
Vi
tis
ving
era
Ros
a&s
Prun
es
pers
ica
Prm
u4.s
arm
enia
ca
I+m
nIs
anry
gdal
us
Abi
ita&
s Pi
nus
insi
gnis
Pi
nus
pina
ster
Pi
nu
s pi
nea
Pinu
s ha
lepe
nsis
La
b&
s
Lac
an
du
la
stoe
cha
Luva
ndul
a la
tijol
ia
23,5
+
5.0
t 6.
8 _+
I,1
6.
1 f
0,9
14.0
f
2,6
49.5
f
39
24.8
f
27
t 3,
9 *
0.5
5.4
f 1.
1 30
,8
).
3.6
35,1
+
_ 3,
7
182
f 2.
8 t
579
f 2.
3 3.
5 f
2,0
66,3
It
1.
9 6.
1 k
1J
28,4
*
4,4
t 5.
6 f
1.3
4,0
* I,
2 25
.7
+ 3,
5 36
,3
_+ 4
,5
2739
f
29
t 6.
5 f
I,0
4,5
f 1.
5 27
.3
f 0,
9 33
,9
_+ 2
.1
26,8
k
2,0
t 6.
4 f
I,0
3,8
f 0,
7 26
.5
f 1.
4 36
.6
f 1,
8
2594
f
5J
t 5,
l f
0.9
5.5
f I,
1 27
.0
f 2.
5 37
,0
f 59
t t t
3,s
; 2.
7
19.8
+
5,6
27L
+I 2
7 26
.9
f 2.
5 23
,4
$- 4
,8
24,3
f
I,2
3.3
* 0.
7 3.
5 f
0,s
62,3
+
6.3
II,0
+
7J
2,3
_+ 1
,4
6.2
It
193
43.9
+
4,3
20.1
+
12
I,6
+ 0,
4 4,
4 f
1.0
50.9
f
3,4
16,5
f
47
1.9
f 1,
O
4.4
It
12
46.7
+
4.6
22.6
f
4.5
1,6
f 0.
7 4,
8 +
2,2
45,1
+
2,7
20,7
)
2,3
7.1 1 7:5
30.4
f
3.9
28J
26.7
25
.9
f 3.
6
t 1 t t t
),I
f 0.
9
2.5
f 0,
9
t t t
5.3
f 1.
6 6.
7 k
1.6
48.4
f
32
8.6
f 0.
8 4,
l 87
9 49
.8
8.9
4,6
6,6
44.6
17
,5
4.9
* I,
0 6,
7 f
1,1
46.4
It
4.7
13
.6 +
3.
0
t 21
.7
f 1.
9 t
19,6
f
2.7
t 20
,4
k I,7
3.5
+ 0.
6 5.
8 f
1.4
39.2
rt
I,
6 29
,9
f 2,
9 3.
0 6.
3 +
I,5
13.7
+
4,9
40.8
k
3,8
19.4
+
3.9
2,8
6.3
+ 0,
6 11
.8 f
3,
5 39
,6
+ 2,
9 21
.7
-f
4.0
2.7
25.6
22
.0
f 2,
7 0,
6*
79
f 2-
7 50
.6
f 4,
4 17
.1 f
12
.2
22.2
f
3,8
1,7
_+ 1
.6
IO,4
+
2,3
47.1
f
3.9
22,2
24
,9
1.1
891
15,4
f
6.6
48,9
25
.7
+ 3,
2 2.
4 k
1.9
11.7
f
1.7
47.1
f
3.6
2.1
+- 0
,s
2.3+
7.
1 &
1.4
1,
3*
5.5
1.8
8,l
+ 2,
6 5,
O +
0.
9
1 26
.7
f 1.
0 2.
7 f
0,3
8,O
f
0.3
182
+ 0.
7 32
1 +
1,2
12.2
f
0,6
t 25
.0
+ 1,
l 3.
1 *
0.5
4.3
k 0.
6 8,
O f
I,
0 36
,4
_+ 1
.5
23.1
f
1.5
t
23
25
3,l
1.9
1.9
20
23 2:
25
29
25
1.8
21
22
22
14.4
*
5.4
1,5
99
It
2,0
I,8
938
;zl
LC
S r&
hats
qu
i co
nccr
nent
le
s ch
ainc
s co
urte
s so
nt
expr
imh
en
:‘,
du
poid
s to
tal
des
acid
es
gras
; ce
ux
qui
conc
erne
nt
les
chah
es
long
ues
en
“/,
du
poid
s to
tal
des
cha*
mes
1O
ngW
L
es
rapp
orts
ha
turb
~JSa
turk
sYo
ont
ete
calc
uli
sur
1e-s
chai
nes
long
ues.
L
es
inte
rval
les
de
cont
ianc
e on
t C
te c
alcu
lCs
au
seui
l de
5%
. L
es
rt!s
ulta
ts
indi
ques
sa
ns
inte
rval
le
de
conf
ianc
e co
rres
pond
ent.
soit
a un
tr
op
faib
le
nom
bre
de
repe
titio
ns
(inf
erie
ur
a 5)
. so
it a
unc
dist
ribu
tion
non
gaus
sien
ne
des
vale
urs+
. D
ans
une
fam
ille
donn
ee.
la
Prcd
re
qxh
indi
quee
cs
t la
pl
us
calc
ifug
c.
la
dcm
iere
la
pl
us
calc
icol
e;
sauf
po
ur
les
Gra
min
&es
, ou
le
s ex
igen
ces
Cda
phiq
ues
sont
pe
u m
arqu
ees
Abr
evia
tions
: C
r6.e
=
acid
e pa
lmiti
que;
C
u.,
= ac
ide
palm
itole
ique
; C
,sIo
=
acid
e st
eari
quc;
C
,,:,
= A
cide
ol
eiqu
e;
C,,:
, =
acid
e lin
olei
que;
C
qu
e;
C,,,
=
acid
e E
heni
que.
,8
.3 =
ac
ide
iinol
eniq
ue;
C,,:
, =
acid
e ar
achi
di-
Lipides racinaires
Tableau 2. Phospholipides des racines entieres
1895,
l%p&XS PI PC PG PE + PS DPG PA
Legumineuses iapims 1zJwUS L.upinuli atbus Vicia @a minor GI,vcine W.Y var. anoka
var. swift Graminkes
Zea muys Sorghum vulgare Hordeurn sativum Triticum sativum
Vita&s Yitis ripariu Vitis vinifera
Rosa&es Prunus pmsica Pwws armeniaca Prunus amygdalus
Ablacks Pinus insignis Pinus pinaster Pinus pinea Pinus h&per&s
Labiks Lavandula lati$olia
8,s * 2,8 35.3 1 0 6,s + 2,l 31.3 f 8,4 7.7 f: 5P lo,4 * 4,l 891 f 3,3 352 f 32 677 f 2.3 36,2 f 7,4 591 f 1.8 %O + 42 7,7 * 1,7 43.4 f 42 65 f 20 33,7 f 3,s 4,9 f 1,4 336 f 1+2 7.8 f 3.6 43,l + 42 7,o f 2,4 33.1 f 2.4 5,l f 197 39 f 197 578 f 26 46,4 f 16 4,9 f 1.8 3399 f 2+6 32 f LO 3,o f 1.2
;1* 21 It 199
51.2 f 3,l 5,7 f 3,l 3621 f 390 3.5 + 98 3,3 f 1,6 46.8 f 5,l 140 f 0.9 30;2fZO 5,2 f 29 50 f 20 50,6 f 26 5,2 f 1-l 36,s f 1,3 4,7 f 1,3 2,4 f 1.6 46,9 f 4*0 694 f 22 33,2 + 5,3 5.6 f Z4 53 4 1,5
2$* 1,4*
42,l f 4,4 47.6 It 4,2
990 f 3,6 4,9*
358 f 7,O 34,7 f 8,7
4.5 f 136 5.2 f 1.2
5.9 f 4,l 5,o k 2,7
1,3* 46,6 f 1,7 8,7 + 59 36,3 + 3,l 23 It 48 42 f 0,7 1.1 f 1,o 41,6 f 1,9 8,8 f 3,9 360 f 2,6 55 f 1,7 7.0 * 1,5
3,1* 38.2 + 1,6 9,O f 3,6 33,5 f 2,7 60 + 2.6 141 f: 2,6
13,s 14.0
29.5 13,s 39,9 6,O 332 143 34,7 102
24.0 24,2
zl$
z 911 9,l
335 f 3,s
11,l 7J 4,6
12,6 82
11.0
52,l * 49 590 f 21 35,5 rt 53 3,3 4 2,9
Les rksuhats sent exprimks en % du phosphore lipidique total. Les intervalles de confiance ont et& calcuf&s au seuil de 5%. Les rksultats indiquks sans intervalle de confiance correspondent soit & un trop faible nombre de repetitions (inferieur a 5) soit B une ~st~bution non gaussienne des valeurs+. Darts une famille donnke, la premike espt?ce indiquk at la plus calcifuge, la demiere la plus calcicole; sauf pour lea Gramintks oti les exigences tdaphiques sont peu marquees. Abreviations: PI = phosphatidylinositol; PC = phosphatidylcholine; PG = phosphatidylglyckol; PE = phosphatidylethanolamine; PS = pbosphatidyl~rine; DPG = diph~phatidylgl~rol; PA = acide phosphatidique.
elusions d~~tives de ces remarques. Gn peut cependant avancet l’hypothtke que I’holution se serait traduite au niveau des acides gras racinaires par une plus grande homog~~it~.
Enfin on observe que le pourcentage d’acides gras saturks semble 6tre plus tlevt chez les espkes calcifuges que chez lea espkes calcicoles (tableau 1 rapport Insa- t&s (%)/Saturb (“A).
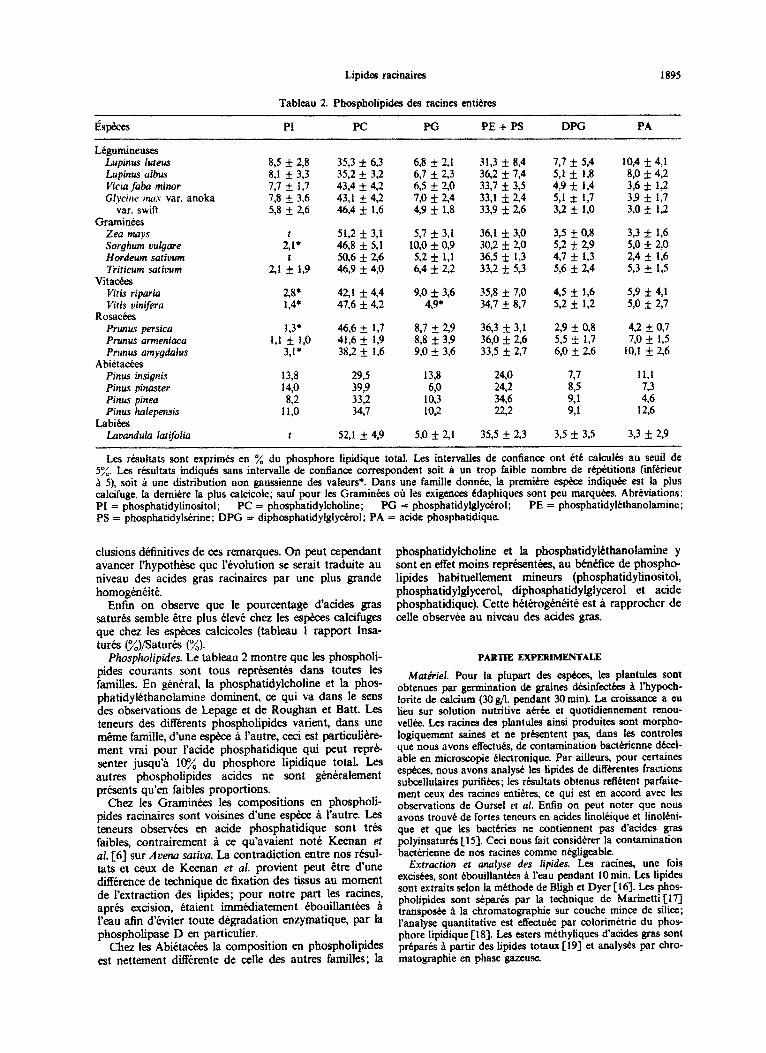
~~osp~~~p~es. Le tableau 2 montre que les phosphoti- pides courants sont tous reprksentks dans toutes les familles. En general, la phosphatidylcholine et la phos- phatidylethanolamine dominent, ce qui va dans le sens des observations de Lepage et de Roughan et Batt. Les teneurs des differents phospholipides varient, dans une m&me famille, dune espkce a I’autre, ceci est particuhtre- ment vrai pour I’acide phosphatidique qui peut reprk- senter jusqu’a lq% du phosphore lipidique total. Les autres phospho~pides acides ne sont gkniralement presents qu’en faibles proportions.
Chez les Graminkes lea compositions en phospholi- pides racinaires sont voisines dune espke a i’autre. Les teneurs observkes en acide phosphatidique sont tr6.s faibles, contrairement a ce qu’avaient not4 Keenan et al. /6] sur Arena sutiuu. La contradiction entre nos r&4- tats et ceux de Keenan et al. provient peut &re dune ditTerence de technique de fixation des tissus au moment de l’extraction des lipides; pour notre part les racks, aprb excision, &tient immkdiatement tbouillantkes a l’eau afin d%viter toute d~~dation enzymatique, par Ia phospholipase D en particulier.
Chez les Abietaaks la composition en phospholipides est nettement di!Terente de celle des autres families; la
phosphatidylcho~ne et la phosphatidyl~~oI~ine y sont en effet moins rep&sent&es, au b&&ice de phospho- lipides habituellernent mineurs (phosphatidylinositol, phosphatidyl~y~ero~, ~phosphatidylgly~rol et acide phosphatidique). Cette heterogknkite est ii rapprocher de celle obserwk au niveau des atides gras.
PARTIE EXPERIMENTALE
Mattiki. Pour la plupart des espkes, les plantules soot obtenues par germination de graines dbinfectks a lhypoch- lorite de calcium (30 g/l. pendant 30 mitt). La croissance a eu lieu sur solution nutritive a&&e et quotidi~n~~t renou- velhk. Les racks des plant&s ainsi produites sont morpho- logiquement saines et ne prksentent pas, dans les controles que nous avons effect& de contamination bacterienne dkcel- able en microscopic &ctronique. Par aiRems, pour certaines espkces, nous avons analyse les lipides de difftrentes fractions subcellulaires purifiks; les r+suhats obtenus reflQent pa&k-- ment ceux des racines entitres, ce qui est en accotd avec les observations de Oursel et al. Entin on peut noter que nous avons trouve de fortes teneurs en acides linolkique et linolkni- que et que les bactkies ne contiennent pas d’acides gras polyinsatur&s [ 151. Ceci nous fait considerer la contamination bacterienne de nos racines comme negligeable.
Extraction et analyse des lipides. Lea racks, une fois exciskes. sont tbouillantkes a l’eau pendant 10 min. Les lipides sont exiraits selon la methode de Bhgb et Dyer [16]. Les phos- pholipides sont s&parks par la technique de Marinetti [17j transoosee a la chromatographie sur couche mince de silk; l’anaiyse qu~ti~tive est &ectuC par colorimetrie du phos- phore lipidique [18]. Les esters methyliques d’acides gras sont prepares a park des lipides totaux [19] et analyses par chro- matographie en phase gazeuse.

1896 M. R~SIGNOL
1.
2.
3. 4.
5. 6.
7.
8. 9.
10.
REFERENCES
Hit&cock, C. et Nichols. B. W. (1971) Plant Lipid Bia- chemistry, p. 387. Academic Press. New York. Galliard. T. (1973) in Form and Function of Phasphalipids, (Ansell. G. B.. Danson. R. M. C. et Hawthorne. J. N. eds). ~ pp. 253 -288. B B A Library, Amsterdam. Lepage. M. (1967) Lipids 2. 244. Roughan, P. G. et Batt, R. D. (1969) Phyrachemistry 8.
363. Bartholomew, L. et Mace, K. D. (1972) Cytobios 5, 241. Keenan. T. W.. Leonard. R. T. et Hodges, T. K. (1973) Cyrobios 7. 103. Holman, R. T. et Nichols, P. C. (1972) Phyfochemistry Il. 333. Ferguson, W. S. (1966) Con. J. Plant. Sci. 46. 639. Kuiper, P. J. C., Kahr, M., Stuiver, C. E. E. et Kylin, A. (1974) Physiol. Plantarum 7, 59. Oursel, A., Lamant, A., Salsac, L. et Mazliak, P. (1973) f’hyrochauistry 12, 1865.
11. 12.
13.
14.
15.
16.
17. 18.
19.
Lamant, A. et Heller, R. (1975) Physial. tig. 13. 685. Gerloff, E. D.. Richardson, T. et Stahmann, M. A. (1966) Plant Physiol. 41, 1280. Grenier, G., Mazliak, P., Tremolitres, A. et Willemot. C. (1973) Physial. Veg. 11, 253. Kaimal, T. N. B. et Lakshminarayama, G. (1970) Phyta- chemistry 9. 2225. Erwin, J. A. (1973) in Lipids and Biomembranes of Eukarya- tic Microorganisms, (Erwin, J. A. ed.), pp.42-143. Aca- demic Press. New York. Bligh, E. G. et Dyer. W. J. (1959) Can. J. Biachem. Physiol. 37. 911. Marinetti, G. V. (1965) J. Lipid Res. 6. 315. Shibuya, I., Honda, H. et Marco, B. (1%7) Agr. Biol. Ghan. 31. 1 II. Metcalfe, L. D., Schmitz, A. A. et Pelka, J. R. (1966) Anal. Chem. 38, 514.





























