Embed Size (px)
Citation preview

B u l l . SOC. Chim. Belg. vo1.85/n012/1976
CONTRIBWION DES CALCULS QUANTIQUES A L'ELUCIDATION
DE LA STRUCTURE EN SOLUTION DE L'ACIDE RIBONUCLEIQUE DE TRANSFERT
PAR LES TECHNIQUES DE LA RMN
B. PULLMAN
I n s t i t u t de Biologie Physico-Chimique Laboratoire de Biochimie Theorique
associe au C.N.R.S. 13, rue P. e t M. Curie - 75005 Par is -
RESUME - Les spectres RMN de haute resolution des acides ribonucleiques de t r a n s f e r t (tARN) pre-
sentent dans l 'eau une s e r i e de resonances comprises en t re -11 e t -15 p.p.m. qui sont a t t r i - buIs aux protons U-N H e t G-N H impliqUes dans l e s l i a i sons hydrogene en t re l e s paires de bases. Les positions3des r6sofiances sont les resul tantes des valeurs "intrins8ques" caracte- r is t iques des paires A-U e t G-C isolees e t de leur perturbation sous l ' e f f e t d'empilement due essentiellement aux courants n de bases. Les calculs quantiques de ces e f f e t s on t per- mis l 'ass ignat ion des resonances observees aux pa i res de bases specifiques e t on t a i n s i con- tribu6 d'une fason decisive I 1'6lucidation de l a s t ruc ture secondaire e t t e r t i a i r e des tAPN en solution.
I. INTRODUCTION
Les raisons qui m'incitent I presenter dans ce &moire ce fragment p a r t i c u l i e r de ma con-
ference pleniere au 7.5 Collcque de Chimistes Quanticiens d'Expression Latine sont doubles.
D'une par t , j e voudrais i n c i t e r l e s chimistes quanticiens, un peu t rop centres peut-&re s u r
l e s problemes des spectrosmpies u l t rav io le t te e t v i s ib le , I etendre leur champ d'action. En
par t icu l ie r , l a resonance magnetique nucleaire (RMN) qui e s t aujourd'hui un des o u t i l s prin-
cipaux de l a chimie e t biochimie s t ruc tura les , me p a r a i t n'avoir a t t i r e que relativement peu
d ' in te r6 t de l a par t des chimistes theoriciens. Deuxiemement, a ceux qui s ' in te r rogent e t
nous interrogent sur l a s ign i f ica t ion pratique des calculs de l a chimie quantique, j e vou-
d r a i s indiquer sur un exemple que j e c ro is assez spectaculaire l e r61e dec is i f que peuvent
prendre de t e l s calculs dans l a solution d'un probleme experimental mmplexe, de grande actua-
lit&. Cet exemple j e l e prendrai dans l e domaine de l 'e lucidat ion de l a s t ruc ture en solut ion
des acides ribonucleiques de t ransfer t (W).
11. LE PROBLEME
L'elucidation de la s t ruc ture secondaire e t t e r t i a i r e des acides ribonucleiques de trans-
f e r t , par t ic ipants a c t i f s essent ie l s dans les processus de synthese de proteines , e s t un des
probl.5mes centraux de l a biologie mol6culaire actuel le . De tres nombreux r e s u l t a t s exp6ri-
mentaux ont d te obtenus durant ces dernieres annees par des etudes aux rayons X en ce qui
- 925 -
concerne l a s t ructure c r i s t a l l i n e de ces composes ( ' ) . Tuutefois, l ' i n t e r l t pr incipal de l a
biologie moleculaire concerne p l u t d t l a s t ruc ture de ces comps4s en .solution et durant ces
dernieres annees de t res nombreux travaux ont 6th consacres d ce probl&re, l a technique prin-
cipale u t i l i s e e &ant l a resonance nucleaire magnetique de haute resolut ion (voir par exemple
2-14).
+ on
t C
I Bmr de 7 I'aminoacidc
p 0. . I 1
I 1 0..
I : 0. .
0 . .
!.l
Bms 1 1 du hU ,-u,t'; B ~ s du T I C
Y'R'A , 0. . -..A
i, r-m-R-Y, O-O-O-O-G, R G.G.A r-.. ' T, Ur.6 : b-o-?-G 'o-~-e-o-C'~ I . . . . .
.... I I 0.. .. 0 . 0 variable 7.7 .-...) Bmr
*\.,. b u r de
I'anticodon Bmr de
I' anticadon
B M S variable
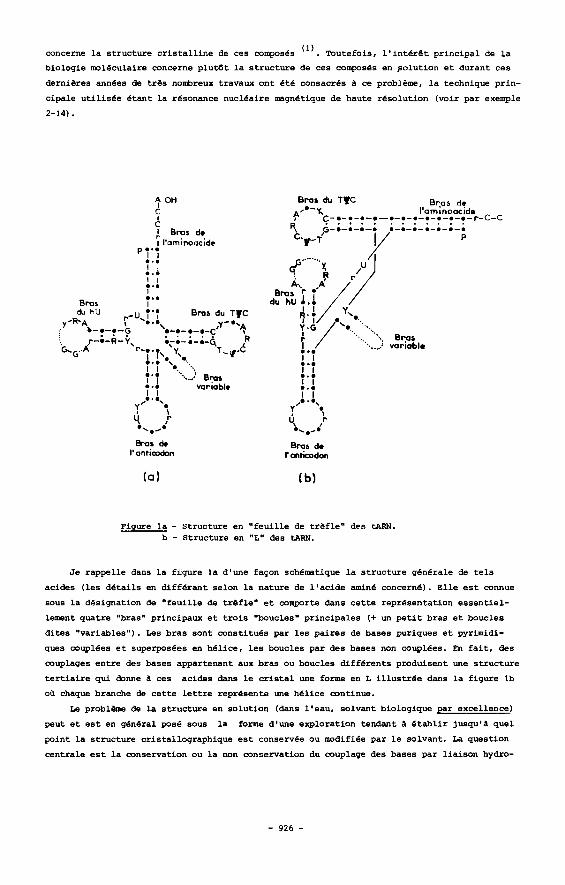
Figure la - Structure en " f e u i l l e de t r & f l e " dQS mFX. b - Structure en "L" des tARN.
J e rappelle dans l a f igure l a d'une facon schematique l a s t ruc ture gBn6rale de t e l s
acides (les d e t a i l s en d i f fe ran t selon l a nature de l ' ac ide amin6 concern€). E l le e s t mnnue
sous l a designation de " feu i l le de t r s f l e " e t comprte dans c e t t e representation essent ie l -
lement quatre "bras" pr inc ipaw e t t r o i s "boucles" pr incipales (+ un p e t i t bras e t boucles
d i t e s "variables"). Les bras sont m n s t i t u e s par l e s pa i res de bases puriques e t pyrimidi-
ques couplees e t superposees en hel ice , l e s boucles par des bases non couplees. En f a i t , des
couplages en t re des bases appartenant aux bras 00 boucles d i f fe ren ts produisent une s t ruc ture
t e r t i a i r e qui &me d ces acides dans le c r i s t a l me forme en L i l l u s t r e e dans l a f igure lb
oil chaque branche de c e t t e l e t t r e represente une hel ice continue.
Le p r o b l h e de l a s t ruc ture en solut ion (dans l 'eau, solvant biologique par excellence)
forme d'une exploration tendant d Otabl i r jusqu'd quel peut e t e s t en general p s 6 sous
p i n t l a s t ructure cristallographique e s t conservee ou modifiee par l e solvant. La question
centrale e s t l a conservation ou l a non conservation du muplage des bases par l i a i son hydro-
l a
- 926 -
gene e t ensui te c e l l e de leur superposition en hel ices , ces caracter is t iquen determinant l e s
elements essent ie l s de l a s t ructure secondaire e t t e r t i a i r e des tARN.
L'out i l pr incipal pour une t e l l e determination e s t la RMN de haute resolution, sur tout
depuis que Kearns, Shulman e t
majorite des resonances protoniques, observees dans l e s spectres des tRRN dans l a region en t re
-11.5 e t -15 p.p.m. sont dues aux protons des paires A-U e t G-C fa i san t p a r t i e des cycles des
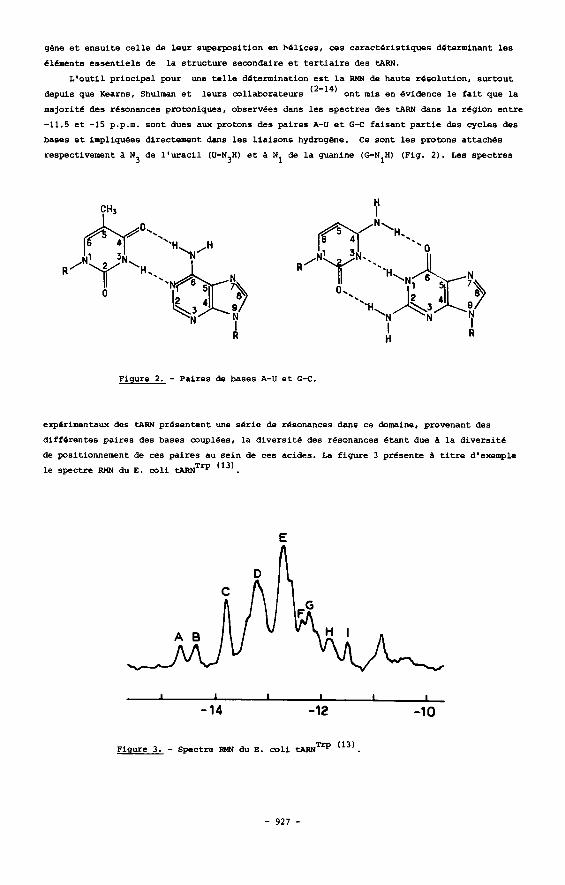
bases e t impliquees directercent dans l e s l i a i sons hydrogPne. Ce sont l e s protons at taches
respectivement a N3 de l ' u r a c i l (U-N,H) e t 1 N1 de l a guanine ( G N I H ) (Fig. 2 ) . Les spectres
leurs collaborateurs (2-14) on t m i s en evidence l e f a i t que l a
Figure 2. - Paires de bases A-U e t G-C.
experimentaux des tARN presentent une s e r i e de resonances dans ce domaine, provenant des
differentes paires des bases couplees, l a d ivers i te des resonances &ant due a l a d ivers i te
de positionnement de ces pa i res au se in de ces acides. La f igure 3 presente 1 t i t r e d'exemple
l e spectre RMN du E. coli ~ A R N ~ ~ ( 1 3 ) .
E
I 1 I 1 1 I
- 14 -12 -1 0
Figure 3. - Spectre RMN du E. coli tARNTrp (13)
- 927 -

L'emplacement observe de resonances e s t l a resul tante de deux e f f e t s contraires . D'une
par t , l a formation meme d'une l ia i son hydrogene entre l e s bases s'accompagne d'un diplace-
merit des risonances U-N H e t G-NIH vers les champs bas par rapport d l eur posi t ion dans l e s
molecules separees (15124). L'origine de c e t e f f e t e s t due sur tout d un e f f e t de champ(25).
D'autre par t , l a superposition ver t ica le des h-ses ou de pai res de bases produl t un deplace-
ment de ces mbes resonances vers l e s champs hauts (voir par exemple 14 e t 26,27) par suite
de l ' e f f e t d ' icran dP
(29). La grandeur de ces derniers deplacements depend naturellement de l 'o r ien ta t ion mutuelle
des bases ou paires de bases impliquies. L a f igure 4 presente schematiquement l ' a l l u r e e t
l'ampleur du phenomene.
principalement (bien que pas uniquement) ('*) au courant II des bases
-11.5
Risononces dons les bases libres
G - N , H Z-10.5
s -11.0 T-N3H \
diplacement vem le champ bible lam de
Rironances dans les de bases e m p i l k
A- T - 14.5 % -14.6
Figure 4. - Origine des resonances dans le domaine -11 d -15 p.p.m. du
U-N-,H e t G-N1H.
Le p r o b l h e avec lequel sont confront& l e s experimentateurs ayant obtenu un spectre
RMN d'un tARN dans l a region concernee e s t de pouvoir effectuer une a t t r ibu t ion des resonan-
ces observees I des paires de bases specifiques. En e f f e t , seule une t e l l e a t t r ibu t ion per-
met t ra i t de rendre compte de la s t ruc ture secondaire e t t e r t i a i r e des U R N en solut ion e t
en par t icu l ie r de comparer c e t t e s t ruc ture avec c e l l e des a s acides dans l e c r i s t a l .
111. LA CONTRIBUTION DE LA THEORIE A LA SOLUI'ION DU PROBLSME.
L'experience seule ne p a r a i t pas pouvair ou tou t au moins n'a pas reussi jusqu ' ic i B
risoudre ce p r o b l h . En revanche, l a th6orie a pu apporter dans ce domaine une CUntribUtiOn
decisive qui a f a i t que ce problame a pu &re t r a i t € avec succes pratiquement des q u ' i l a i t 6
- 928 -

pose. Aujourd'hui l a s t ruc ture en solut ion d'un grand nombre des tARN e s t dans ses grandes
l ignes Gterminee, l e s & r i t e s en revenant a l a contribution decisive de l a theorie pour
l ' in te rpre ta t ion des spectres RMN de ces acides.
Le probl&oe theorique au depart e s t double. 11 s ' a g i t en premier l i e u de determiner ce
que s e r a i t l a position de l a resonance de U-N H e t G-N H dans des paires des bases complemen-
t a i r e s AU e t GC i so lees , non soumises a l ' e f f e t d'empilement (resonance " intr insMue") . I1
n 'ex is te pas de renseignemmts experimentaux a ce s u j e t , l e s bases l i b r e s ayant dans l ' eau
une tendance de s ' assoc ier par empilement v e r t i c a l p lu td t que par l i a i sons hydrogene (voi r
par exemple 30, 3 1 ) . Une f o i s ce renseignement de depart obtenu, il s ' a g i t de determiner
jusqu'a quel point ce deplacement de resonance vers l e champ fa ib le par l a formation des
l ia isons hydrogene e s t cornpens6 par un deplacement vers l e champ haut sous l ' e f f e t de l'em-
pilement de ces paires dans l e s par t ies hel icoidales de l ' A R N , &ant entendu que ce second
e f f e t e s t essentiellement vari'able d'une pa i re a l ' a u t r e en fonction de l a nature e t de l ' a r -
rangement mutuel des bases e t pa i res des bases superposCes.
questions. En ce qui concerne l a premiere nous venons d'evaluer treS recemment (32) l e s va-
leurs du deplacement vers l e champ bas des reronances de U-N,H e t G-NIH sous l ' e f f e t de l a
formation des paires de bases A-U e t G-C, fournissant a i n s i l ' information directe sur l a va-
leur des resonances intrins&ques de ces protons. En ce qui concerne l a d e u x i h e , Giessner-
Pre t t re e t Pullman ont
bases des acides nucleiques l e s deplacements chimiques devant se produire sous l ' e f f e t du
courant n d e ces bases pour des protons des molecules voisines s e trouvant dans d i f fe ren tes
regions d'un plan s i t u e a 3 . 4 A au-dessus (ou au-dessous) du plan de l a base pr i se en consi-
deration ( l a distance de
l e s acides nucleiques). Etant donne l'importance his tor ique de ces courbes, nous l e s repro-
duisons i c i dans l a f igure 5 pour l e s quatre bases pr incipales , tout en s ignalant que ces travaux ont €te aujourd'hui etendus a toutes l e s dis tances ver t ica les jusqu'a 7 ( 3 3 ) e t
qu'ont 6 t e egalement Bvaluees l e s contributions des facteurs complementaires t e l s que l ' e f f e t
d'anisotropie atomique ou "locale" (28) .
3 1
Aujourd'hui une reponse directe peut &re fournie par l a theorie quantique a ces deux
BvaluC d8s 1970 (") des courbes d ' isoecran indiquant pour chacune des
3 . 4 A e s t l a distance moyenne habi tuel le d'empilement de bases dans
Toutefois, il peut 6 t r e interessant de s ignaler qu'au moment oii l e s experimentateurs se
SOnt trOuv8s confront& pour l a premiere f o i s avec l e probleme de l ' i n t e r p r e t a t i o n de spectres
RMN des tARN (vers 1973) seuls 6 ta ien t connus d'eux nos r e s u l t a t s concernant l e s courbes
d'isoecran pour l e deplacement chimique dues a l'empilement des bases. I1 leur manquait donc
l e renseignement primaire mncernant l e s resonances "intrinseques" de U-N3H e t G-N H dans l e s
paires des bases non perturbees. (En r e a l i t 4 ce renseignement e x i s t a i t dans une Note aux
Comptes Rendus publiee par nous en 1970 (25) mais qui e s t passee inaperpue des experimenta-
teUrS). En fa i san t appel a des experiences ingenieuses sur des modeles oligonucleotidiques,
des experimentateurs ont reuss i a determiner indirectement c e t t e donnee manquante en s e ser-
vant pour ce la de nos donn6es theoriques sur l e s courbes d'isoecran. Le pr incipe du procede
e s t l e suivant (voi r par exemple 2 3 ) . Considerons un deoxyoligonucleotide (11-13) d-ADTDGD
1
C A T qui en solut ion dans l ' eau forme une double P P
_ _ _ hel ice auto-complBmentaire :
A T G C A T .P:P:P:P:P: . . . . . . . . . . . . . . . . . . . . . . . . T A C G T A P P P P P
- 929 -

I 1 i H
Figure 5. - Courbes d'isodeplacement chimique pour des protons s e trouvant B 3.4 A au-dessus (ou au-dessous) du plan des bases (29) .
En spectroscopie RMN c e t t e he l ice presente dans l e champ bas t r o i s resonances B -13.35, -13.8
-12.7 P.P.m. correspondant de toute evidence aux t r o i s
une
lors de l ' i l eva t ion de
indiqud. en commenpant par c e l l e a -13.35 p.p.m e t terminant par c e l l e a -12 p.p.m, indiquant
par 1.3 l a dissolut ion progressive des pai res de bases.
de pa i res de bases pr isentes :
pai re A-T terminale, une pa i re A-T in te r ieure e t une pa i re G-C centrale . Par a i l l e u r s ,
l a temperature ces resonances disparaissent ("fondent") dans l 'o rdre
Les informations thermodynamiques que l 'on possede s u r l e s s t a b i l i t e s re la t ives des
paires de bases (voir par exemple 34, 35) permettent d'affirmer sans grand risque d 'erreur
que dans l e cas qui nous interesse, l a plus durable des resonances doi t appartenir B l a p a i r e
G C , que l a resonance de pers is tance intermediaire doit appartenir d l a pa i re A-T i n t i r i e u r e
e t que l a resonance qui d i spara i t l a premiere doit carac tdr i ser l a pa i re A-T terminale. Les
frequences a i n s i assignees resu l tan t evidemment de l a combinaison des e f f e t s dus B l a forma-
t ion des l ia i sons hydrogene e t de ceux dus a l'empilement. Neanmoins, l 'ass ignat ion &ant
f a i t e , si l 'on adopte pour l a double hel ice m e s t ructure determinee, disons c e l l e du B-ADN,
dont l e s caracter is t iques g i o d t r i q u e s sont connues, on peut a l o r s evaluer, grace a nos cour-
bes d'isoecran, quel le s e r a i t pour l e s d i f fe ren ts types de pai res de bases presentes l a a m p -
- 930 -

sante du &placement chimique
nous l ' indiquons dans
vers l e champ haut due 1 l'empilement. Ainsi par exemple come
l a p a r t i e superieure de l a f igure 6 ce deplacement 6valu6 theoriquement
Ditermination de friquences <'intrin&ques')
DGplacement vers le champ Risonance Paire de Risonance ,, effct de eintrin&que*
Observe' I ' empilement de bases bases
G - C -12.7 - 0.9 -13.6
A-T int. -14.6
Cas de A - T terminal
A-T term. -13>\ r%zce -
divergence d w -13.9 a %-aying* - 14.6
Figure 6. - Wtermination semi-empirique des resonances intr inssques.
s e r a i t de 0.9 p.p.m. pour l a pa i re G-C e t 0.8 p.p.m. pour l a paire A-T. A p a r t i r de c e t t e
donnee on peut a lors prevoir, par soustract ion, quel le s e r a i t l a valeur de resonance " in t r in-
s&que" des paires isolees ( f i g . 6 ) : e l l e s e r a i t de13 .6 p.p.m. pour l a pa i re G-C e t de -14.6
p.p.m. pour la paire A-T. Bien que d i f fe ren ts auteurs u t i l i s a n t d i f fe ren ts exemples trouvent
des valeurs legsrement d i f fe ren tes , e l l e s sont toutes dans l'ensemble t res proches des valeurs
prdcitees.
Le cas de l a paire A-T terminale e s t special e t montre l e s precautions q u ' i l convient
de prendre dans ce genre de t rava i l . En e f f e t , l e deplacement vers l e champ haut, dCi a l'empi-
lement, &value pour c e t t e pa i re , e s t de 0.07 p.p.m.. Combine A l a valeur intrinsSque de l a
resonance de l a paire A-T f ixee plus haut a -14.6 p.p.m., il conduit 1 prevoir pour c e t t e
paire une resonance s i tuee vers -13.9 p-p-rn. (Fig. 6 ) . Or l a resonance exHrimentale e s t a -13.35 p.p.m.. La divergence n'infirme en f a i t nullement l a th lor ie ; e l l e e s t due au phenomene
de "fraying" de l a paire t e m i n a l e qui consis te essentiellement en son decouplage p a r t i e l .
Q u o i q u ' i l en s o i t , les resonances "intrinssques" des paires A-T (ou A-U, e l l e s ne SOnt
pas supposees Ctre tres differentes) f ixees , e t comme nous l 'avons vu f ix6es en bonne p a r t i e
grace a l a contribution de l a theorie , il e s t a lors a i s e , t o u t au moins en principe, de deter-
miner l e s resonances previs ibles pour l e s d i f fe ren tes pa i res de bases d'un tARN, en calcu-
- 931 -
l a n t pour chaque paire l e deplacement prevu vers l e champ haut sous l ' e f f e t du courant II des
paires de bases ou des bases adjacentes e t en u t i l i s a n t pour cela l e s murbes de l a f igure 5 ,
I condition naturellement d'avoir adOpt6, pour l ' e d i f i c e une g 6 o d t r i e appmpriee. La g6om6-
t r i e adoptee e s t en general c e l l e de l ' h e l i c e A ou A'-ARN.
Tableau I. - Attribution des resonances dans l e spectre RMN
de E. c o l i tRNATrP
Posi t ion observee Resonance Intensi te Position
prevue Paire
A 0.8 14.68 14.8 A14- (S4)U8
D
E
B 0.8 14.40 13.8 A412
A 4 3 1
13.4 -53
1.0 13.82 14.2
1.0 13.80 13.8 '-'52 1.0 13.42
1.0 13.30 13.30 A-U1
1.0 13.24 13.25 G-C30 1.0 13.20 13.15
1.0 13.12 13.15
1.0 13.06 12.75
1.0 12.83 12.80 G-CZ
Dc6 1.0 12.76 12.70
1.0 12.74 12.65
1.0 12.69 12.65 G-C4
C
=5 t e r t i a i r e
G-C2 7
GC3
1.0 12.67 12.65 G-C49
12.55 GC7
1.0 12.53 12.50 G-ClO F 1.0 12.36 12.40 G-c5 1
G-c2 8
1.0 12.58
1.0 12.23 12.20
1.0 12.11 12.20 G-c29 G
H 1.0 11.82 - u protege (prob. U33)
I 13 1.0 11.49 11.50 G-C
Le pro&d€ s 'avere extrCmement eff icace. I1 conduit I une ident i f ica t ion de l 'apparte-
nance de chaque resonance e t grosso mod0 a permis d ' e t a b l i r que l a s t ruc ture des tARN en solu-
t ion est tr& sensiblement identique I c e l l e observee dans l e s cr is taux correspondants, bien
que cer ta ines differences s ign i f ica t ives paraissent s e manifester Bgalement. A i n s i , par exem-
ple , a lors que l e s cr is taux de cer ta ins tRNA paraissent comporter des paires de bases non usuelles, en par t icu l ie r G-U, couplds par l i a i sons hydrogene, ce couplage ne p a r a i t pas se
conserver en solution (voir par exemple 12) .
I (13). 11 concerne l e ~ R N A ~ ~ dont l a s t ruc ture e s t indiquee dans l a f igure 7. On peut cons- t a t e r que dans l'ensemble l 'accord en t re l e s previsions theoriques e t l e s observations expe-
rimentales e s t excel lent , l ' e c a r t entre l e s deux 6 tan t en general in fer ieur 0.1 p.p.m. qui
Un exemple de l a cornparaison de l a theorie e t de l 'exp6rience e s t donne dans l e tableau
- 932 -

A I C
C
G
I 1 C * G I I
G * C I I
SOG C a
Figure 7. - wTT.
represente l a precision meme des mesures. Dans l a pratique tou t n ' e s t pas auss i simple naturel-
lement que dans ma presentation schematique e t l ' a t t r i b u t l o n de cer ta ines resonances necessi te
une discussion plus serree ( c ' e s t l e cas par exemple de l!inversion des a t t r ibu t ions pour l e s
paires A-ulZ e t A-U31 dans 1s Tableau I ) . Toutefois, ces d e t a i l s techniques e t autres , quelque
s o i t leur importance, sont en dehors du but de ce t a r t i c l e . Les lecteurs interesses p u r r o n t
en trouver l a presentation dans l e s memoires originaux.
Le procede d e c r i t i c i e t qui f a i t si largement appel i l a contribution de l a chimie
quantique represente actuellement l e myen essent ie l pour l a determination de l a s t ruc ture en
solution des tARN. A ce jour il a ete u t i l i s e dans de nombreux laboratoires , pour plusieurs
tARN di f fe ren ts (2-15).
En plus de sa s ign i f ica t ion d i rec te pour ce probl&ue, l a contribution de l a theorie a des
repercussions indirectes pour tou t un ensemble de p r o b l a e s qu 'eclaire l ' a t t r i b u t i o n des reso-
nances RMN a des
t ion suivre dans l e s d e t a i l s l e changement de s t ructure moleculaire d'un tARN sous e f f e t de
l a chaleur, l a d i spar i t ion progressive des resonances avec 1'Blevation de l a temperature pou-
vant &re r e l i e e a l a dis1oca:ion des pa i res determinees (par exemple 9.24) . De mbe, l ' e f f e t
de cations
mes de cwrdinat ion avec des s i t e s bien d6terminLs (par exemple l o ) . Pareillement, l'observa-
t ion des modifications produites dans l e spectre par interact ion avec des agents d ' interca-
lement, t e l par exemple l e bromure d'ethidium (36'37), permet de loca l i se r avec precision l e s
paires de bases specifiques des tARN. Ainsi on peut grace a c e t t e a t t r ibu-
par exemple Mg++ sur l e spectre RMN des tARN peut a i n s i Ctre interpret6 en t e r -
- 933 -

sites de l ' in te rac t ion . O n a pu par e x e m p l e determiner de cette f a p o n glle l e site d ' interca-
lement de cette substance qu i est un i m p o r t a n t agent antiviral e t ant ibacter ien, dans tRANPhe
se situe au niveau des paires AU6 e t AU7 (36).
L e domaine de l a RMN des acides ribonucleiques de transfert n ' e s t pas le seul dans le-
quel nos courbes d'isoecran des bases puriques e t pyrimidiques ont trouv6 un usage extrsmement u t i l e . I1 e n est de meme pour d 'autres p r o b l e m e s analogues comme par e x e m p l e l ' in tercalement
de l'actinomycine e t d 'autres agents dans les helices doubles (38-43), la c o n f o r m a t i o n des
c o e n z y m e A (53) etc. oligo- e t ply-nucl6ot ides ( p a r exemple 44-50) , des c o e n z y m e s d'oxydo-reduction (51-52) r du
BIBLIOGRAPHIE
1)
2 ) 3) 4 ) 5)
6)
7) 8 )
9) 10) 11) 12)
13) 14)
15) 16) 17) 18) 19) 20) 21) 2 2 ) 23) 241 25) 26) 27) 2 8 ) 29) 30)
31)
32)
33)
34) 35)
36) 371 38) 39) 40) 411 42 )
P o u r une revue voir S.H. Kim, P r o g r e s s i n N u c l e i c A c i d R e s e a r c h and Molecular B i o l o g y , 17, 181 (1976). D.R. Kearns, D.J. Patel e t R.G. Shulman, N a t u r e , 229, 338 (1971). R.G. Shulman, C.W. H i l b e r s , D.R. K e a r n s , B.R. R e i d e t Y.A. Wong, J. Mol. B i o l . 3, 57(1973). R.G. Shulman, C.W. H i l b e r s , Y.P. Wong, K.L. Wong, D.R. L i g h t f o o t , B.R. R e i d e t D.R. K e a r n s , Proc. N a t l . A c a d . Sci . USA, 70, 2042 (1973). D.R. K e a r n s , D.R. L i g h t f o o t , K.L. Wong, Y.P. Wong, B.K. R e i d , R.G. S h u l m a n e t L. C a r y , Ann. N.Y. A c a d . Sci . , 222, 324 (1973). K.L. Won9 e t K.R. Kearns, B i o p o l y m e r s , 13, 371 (1974). L.S. &an, P.O.P. TS'O, F. van der H a a r , M. Spr inzl e t F. C r a m e r , B i o c h e m i s t r y , 14, 3278 (1975). K.L. wong, Y.P. wong e t D.R. K e a r n s , B i o p o l y m e r s , 14, 749 (1975). B.F. Bordorf, D.E. K e a r n s , H. E r i n e t s . H . C h a n g , B i o p o l y m e r s , Is, 325 (1976). G.T. R o b i l l a r d , C.E. Tarr , F. Vosman e t H.J.C. B e r e n d s e n , N a t u r e , 262, 363 (1976). G.T. R o b i l l a r d , C.W. H i l b e r s , B.R. Reid, J. G a n g l o f f , G. D i r h e i m e r e t R.G. S h u l m a n , 810- chemistry, Is, 1883 (1976). C.R. Jones, D.R. K e a r n s e t K.H. Muench, J. M o l . B i o l . , 10), 747 (1976). Pour une revue voir D.R. K e a r n s , Progress i n N u c l e i c A c i d R e s e a r c h and Molecular B i o l o g y , 18. 91 (1976). R.R. Shoup, H.T. Miles e t E.D. Becker, B i o c h e m . B i o p h y s . Res. Comm. 2, 194 (1966). L. K a t z e t S . P e n m a n , J. Mol . B i o l . Is, 220 (1966). L. K a t z , J. M o l . B i o l . 44, 279 (1969). R.A. Newmark e t C.R. C a n t o r , J. Am. Chem. SOC. s, 5010 (1968). S.M. Wang e t C. L i n , J. Amer . Chem. SOC. 90, 5069 (1968). K.H. Schei t , Angew Chem. Intern. E d . 5, 180 (1967). D.M. C r o t h e r s , C.W. H i l b e r s e t R.G. Shulman, Proc. N a t l . A c a d . Sci. USA, E, 2899 (1973). D. P a t e l e t A.E. T o n e l l i , Proc. N a t l . A c a d . Sci. USA, 2, 1945 (1974). D. P a t e l e t A.E. Tonel l i , B i o p o l y m e r S 13, 1943 (1974). N.R. Kallenbach, W.E. D a n i e l e t M.A. K a m i n k e r , B i o c h e m i s t r y , Is, 1218 (1976). C. G i e s s n e r - P r e t t r e e t B. P u l l m a n , C o m p t e R e n d . A c a d . Sci. Paris, 270, 866 (1970). A.D. cross e t D.M. C r o t h e r s , B i o c h e m i s t r y , lo, 4015 (1971). L.S. Kan, P.N. Borer e t P.O.P. Ts 'o , B i o c h e m i s t r y , 14, 4864 (1975). C. G i e s s n e r - P r e t t r e e t B. Pullman, B i o c h e m . B i o p h y s . Res. Comm. '0, 578 (1976). C. G i e s s n e r - P r e t t r e e t B. Pullman, J. Theoret. B i o l . 2, 81 (1970). P.O.P. TS'O dans Molecular A s s o c i a t i o n s i n B i o l o g y (B. P u l l m a n E d . ) A c a d e m i c Press, New Y o r k , 1968, p. 39. P.O.P. Ts 'o dans Fine Structure of P r o t e i n s and N u c l e i c A c i d s (G.D. Fasman e t S.N. T i m a s h o f f E d s . ) Marcel D e k k e r , New Y o r k 1970, p. 49. C. G i e s s n e r - P r e t t r e , 8. Pullman e t J. cai l le t , N u c l e i c A c i d Research, (1976). sous presse. c. G i e s s n e r - P r e t t r e , B. P u l l m a n , P.N. Borer, L.S. R a n g e t P.O.P. Ts'o, B i O p o l y m e r s , sous presse. A. Pullman e t B. P u l l m a n , Adv. i n Q u a n t u m Chem. 5, 267 (1968). 8 . P u l l m a n e t A. Pullman, Progress i n N u c l e i c A c i d R e s e a r c h and M o l e c u l a r B i o l o g y , 9, 327 (1969). C.R. Jones e t D.R. Kearns, B i o c h e m i s t r y , 14, 2660 (1975). T.R. K r u g h , F.N. W i t t l i n e t S.P. C r a m e r , B i o p o l y m e r s , 14, 197 (1975). N.S. A n g e r m a n , T.A. V i c t o r , C.L. B e l l e t S.S. D a n y l u k , B i o c h e m i s t r y , 2, 2402 (1972). T.R. K r u g h e t J . W . N e e l y , B i o c h e m i s t r y , 11, 4418 (1973). D. P a t e l , B i o c h e m i s t r y , 2. 1476 (1974). D. Patel , B i o c h e m i s t r y , 11, 2388 (1974). D. Patel, B i o p o l y m e r s , Is, 533 (1976).
D.R. L i g h t f o o t , K.L. Wong, D.R. K e a r n s e t B.R. R e i d , 3. M o l . B i o l . 2, 71 (1973).
- 934 -

43) J. B a r b e t , B.P. R o g u e s , S. C o m b r i s s o n e t J . B . Le P e w , B i o c h e m i s t r y , Is, 2642 (1976). 44) P.O.P. Ts'o, N.S. Kondo, M.P. S c h w e i z e r e t D.P. H a l l i s , B i o c h e m i s t r y , 8, 997 (1969) . 45) N.S. K o n d o , A.M. H o l m e s , L.M. Stempel e t P.O.P. Ts'o, B i o c h e m i s t r y , 9, 3479 (1970) . 46) L.S. Kan, J.L. B a r r e t t and P.O.P. Ts'o, B i o p o l y m e r s , 12, 2409 (1973) . 47) P.N. B o r e r , L.S. K a n , P.O.P. TS'O B i o c h e m i s t r y , 14, 4847 (1975) . 48) L.S. Kan, P.N. Borer e t P.O.P. Ts'o, B i o c h e m i s t r y , 14, 4864 (1975) . 49) D.B. D a v i e s e t S.S. D a n y l u k , B i o c h e m i s t r y , 14, 543 (1975) . 50) N.S. K o n d o e t S.S. D a n y l u k , B i o c h e m i s t r y , z, 756 (1975) . 51) R.H. Sarma e t R.J. Mynott, J. Am. Chem. SOC. 95, 7470 (1973) . 52) M. Kainosho e t Y. K y o g o k u , B i o c h e m i s t r y , 2, 741 ( 1 9 7 2 ) . 53) Ch. H. Lee e t R.H. S a m , J. Am. Chem. Soc. 97, 1225 (1975) .
- 935 -





























