Embed Size (px)
Citation preview

ment 382 (2007) 22–29www.elsevier.com/locate/scitotenv
Science of the Total Environ
Detection of genotoxicity in the marine environment: A preliminaryfeasibility study using primary mussel tissue culture
Michel Cornet ⁎
UMR 5805 EPOC "Environnements et Paléoenvironnements Océaniques", Université Bordeaux 1, CNRS, Avenue des Facultés,33405 Talence Cedex, France
Received 13 June 2006; received in revised form 16 March 2007; accepted 22 March 2007Available online 4 May 2007
Abstract
The purpose of this study was to evaluate the feasibility and potential usefulness of primary cultures of somatic tissues fromadult mussel by means of sister chromatid exchange induction (SCE). This research is an initial pilot study carried out with musselmantle tissue using seawater artificially contaminated with cadmium and polluted seawater from the port of Arcachon. With regardto cadmium concentration, mean SCE numbers showed a progressive increase from 1.07±0.18 per diploid cell in controls (i.e.cultures without contaminant) to 2.91±0.42 per diploid cell for the highest concentration, 10–4M. With regard to the mediumprepared with seawater from the port of Arcachon, the mean SCE number reached a value of 5.85±0.85 per diploid cell. Theanalysis of SCEs induced by cadmium showed DNA responses even at the lowest concentration (i.e. 10–7 M). The studydemonstrates the feasibility of the sister chromatid exchange (SCE) approach based upon primary mussel tissue culture, for thegenotoxicity testing of contaminated seawater. Highlights from this procedure are (1) the presence of an active cell proliferation, (2)the use whole-water samples, (3) the possibility of culturing without serum, (4) the absence of cell dissociation before culturing and(5) a cellular proliferation which can be obtained in cultures carried out in a medium containing seawater whose salinity is comprisebetween 28 and 35‰.© 2007 Elsevier B.V. All rights reserved.
Keywords: Primary culture; Genotoxicity; SCE; Cadmium; Polluted seawater; Mytilus galloprovincialis
1. Introduction
Each year, a large number of chemicals are conti-nually entering aquatic environments and can bioaccu-mulate in the living organisms. A significant proportionof these inputs, mainly of anthropogenic origin, have thepotential to induce DNA and/or cell damage or interferewith the processes involved in cell division (Livingstoneet al., 2000). Xenobiotic compounds may affect the
⁎ Tel.: +33 5 40 00 87 34; fax: +33 5 40 00 87 01.E-mail address: [email protected].
0048-9697/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.scitotenv.2007.03.033
genetic material of exposed organisms by either causingdirect alterations in genes and gene expression orselective effects on gene frequencies (Pesch et al., 1981;Depledge, 1998).
Genetic ecotoxicology examines the effects ofxenobiotic compounds on the structure and function ofDNA (Theodorakis, 2001). During the past decades,cytotoxicity tests with in vitro cultured cell lines ofvertebrates (mammals and fish, Freshney, 2001) havelargely replaced in vivo acute toxicity bioassays(Birmelin et al., 1999; Ferro and Doyle, 2001). Anincreasing number of studies are now concerned with

23M. Cornet / Science of the Total Environment 382 (2007) 22–29
cells of marine invertebrates maintained in vitro,particularly bivalve molluscs, for toxicological investi-gations (e.g. Burgeot et al., 1995; Zaroogian andAnderson, 1995; Birmelin et al., 1998; Mitchelmore etal., 1998; Domart-Coulon et al., 2000; Le Pennec and LePennec, 2001, 2003).
A method of mussel tissue culture with active cellproliferation was recently developped in our laboratory(Cornet,1993, 1995, 2000) to obtain metaphase-chro-mosomes intended for ecological and phylogeneticapplications. We hypothesized the method could alsobe used as a genotoxicity bioassay by taking advantageof the high quantity of seawater (70%) incorporated inthe culture medium: if tissues are grown in a mediumcontaining seawater, it is likely the water quality willinfluence the quality of the cells produced duringcultivation. Especially, if seawater contains genotoxiccompounds, DNA damages could appear and bedetected in the proliferating cells. Therefore, it wouldbe possible to assess seawater genotocicity by compar-ison of DNA integrity in tissues grown in two differentmedia, one prepared with the seawater to be tested andthe other prepared with a non-contaminated referenceseawater.
Accordingly, the main objective of the present workwas to evaluate the potential of primary mussel tissueculture for the genotoxicity testing of contaminatedseawater. This research is an initial pilot study carriedout with mussel mantle tissue using seawater artificiallycontaminated with cadmium and polluted seawater.Mussel mantle cultures were exposed to cadmium asreference toxicant because cadmium is one of the mostsignificant heavy metals in the Garonne river —Gironde estuary system (France), mainly as the resultof historical mining activity and discharges from metalsmelters located on the Lot, a tributary of the Garonneriver (Grousset et al., 1999; Schäfer et al., 2002; Audryet al., 2004). This contamination deserve to be studiedgenotoxically because of the prominent economicalpotential of the Marennes-Oléron bay (mussel andoyster production) and of the Nord Medoc breedingbasins (prawn production) located nearby the Girondeestuary (Baudrimont et al., 2005).
It is well recognised that cadmium is a toxic heavymetal of environmental concern with a wide variety ofadverse effects. Several studies have shown thatcadmium produces DNA strand breaks, DNA-proteincross-links, oxidative DNA damage, chromosomalaberrations, dysregulation of gene expression resultingin depressed apoptosis and/or altered DNA repair(Mourón et al., 2004). Cadmium is generally regardedas highly toxic but only weakly mutagenic (Rizki et al.,
2004). Marine bivalves including mussels are exposedto varying levels of cadmium in their natural habitats.They have an ability to concentrate cadmium in softtissues from water and sediments on a large scalecovering the whole range of Cd concentrations in eventhe most contaminated estuaries and coastal waters(Cossa, 1989). In their natural environment, musselswould be protected from any potential direct genotoxiceffect of Cd by mechanisms such as the induction ofmetallothioneins or the removal of damaged molecules(Pruski and Dixon, 2002). In oysters, Boisson et al.(2003) have observed that cadmium is normally boundto metallothioneins which would be responsible for thereduction of the bioavailability and toxic effects of Cd.
DNA strand breaks represent a convenient quantifi-able response to genotoxic insult. Strand breaks occurunder normal conditions, but exposure to genotoxicantscan increase the levels of strand breakage, either directlythrough chemical impact, or indirectly through theprocess of excision repair (Shugart and Theodarakis,1994; Shugart, 2000; Dixon and Wilson, 2000).
Among the cytogenetic endpoints most frequentlyused in DNA strand breakage evaluation is the sisterchromatid exchange (SCE). SCE results from thereciprocal exchange of DNA between two sisterchromatids of a duplicated chromosome. Although themolecular mechanism of formation of SCEs is notknown, it has been sugested that DNA damage anddefective DNA repair may cause SCEs after exposure togenotoxic agents (Kanaar et al., 1998). In vitro, SCEscan be visualized by growing cells with 5-bromo-2′-deoxyuridine (BrdU) for two consecutive replicationcycles (Wojcik et al., 2004). The differentially stainedchromatids (the chromatid with more BrdU stain lighter)allow to evaluate the exchange frequency as anindication of treatment-induced genetic damage. Aconvenient alternative to the FPG technique utilizesthe substitution of thymidine with biotin-dUTP (Bruck-mann et al., 1999).
In this context, primary cultures of somatic tissues ofadult mussel were tested for their suitability to assess thegenotoxic potential of polluted seawater samples bymeans of sister chromatid exchange induction.
2. Materials and methods
2.1. Sampling
Mussels of the species Mytilus galloprovincialis(25–30 mm shell length) were collected at Cap Ferret, inthe Bay of Arcachon (French Atlantic coast) in April2002. In the laboratory, they were kept in a 5 l capacity

Fig. 1. Schematic representation of the culture medium composition.The solvent consist of a mixture of seawater (70%) and distilled water(30%). The dissolved powdered MEM contains salts, amino acids,vitamins, phenol red and glucose. Three other components wereadded: chicken serum, HEPES and antibiotics (penicillin andstreptomycin). The test of genotoxicity is based on the presence ofthis high proportion of seawater: if it contains contaminants it isexpected to yield DNA damage in the proliferating cells grown in themedium.
24 M. Cornet / Science of the Total Environment 382 (2007) 22–29
aquaria of natural aerated seawater maintained at 18 °C,with a photoperiod of 14 h light/10 h dark, until theywere required for tissue culture. Half of the water wasrenewed every two days. Animals were fed with anindividual daily ration of 0.05 mL of "Liquifry marine"(Interpet Ltd, Dorking, UK), a liquid artificial foodintended for marine filter feeders.
The non-contaminated reference seawater used formedium preparation was sampled in an oceanic sector ofthe Bay of Arcachon (buoy no. 13, salinity 35‰) at theend of the rising tide.
The polluted seawater used for experiments wassampled at a depth of 0.5 m at the entrance of the port ofArcachon in June 2002 (salinity 33.5‰).
2.2. Tissue culture
The techniques used for culturing, harvesting andslide preparation were identical to those describedpreviously for Mytilus edulis (Cornet, 1993, 1995) andCrassostrea gigas (Cornet, 2000). After dissecting out,mantles were washed in three changes (20 min each) ofantibiotic-antimycotic enriched medium and rinsed insterile serum-free medium. Mantles were then cut into4 mm2 fragments (explants). Cultures were performedin plastic culture tubes containing 3 ml of medium and 4explants of mantle. Basal growth medium was Eagle'sMinimum Essential Medium (powdered MEM, Sigma)with Hanks' salts and L-glutamine. Osmolarity wasincreased with seawater (see medium B, Cornet, 1993):700 mL of seawater+300 mL of distilled water wereadded to the powdered MEM to reconstitute 1 L ofcomplete growth medium (Fig. 1). The pH was adjustedto 7.60 with 1 M NaOH and stabilizided with HEPES(15 mM). This technique allows a good maintenance ofthe pH without the necessity for a CO2 incubator. Themedium was supplemented with 13% chicken serum(Eurobio) and, to prevent contamination, antibioticswere used at a final concentration of 100 IU/mL forpenicillin and 100 μg/mL for streptomycin. Culturetubes were incubated at 18 °C for 6–7 days in dynamiccondition by placing in a roller drum (0.25 rpm). Theroller drum maintains cultures in dynamic condition thathave proven to be beneficial for cell growth. It preventsexplants from adhering to the tube walls and favour O2/CO2 exchanges between the different compartments ofthe culture system, the result being an acceleration ofcell metabolism (Cornet, 2000).
In order to obtain chromatid differentiation, on thethird, fourth, and fifth day of culture, the medium wasrenewed with medium containing BrdU (5.10–5 M finalconcentration).
2.3. Tests
For this preliminary approach, only a limited numberof cultures were performed. Seven culture tubes contain-ing each four mussel mantle explants were prepared(Table 1). The first tube contained medium with nocontaminant (control). The next four tubes receivedincreasing concentrations of CdCl2, 2H2O. A series from10–7 M (=2.10–5 g L−1) to 10–4 M (=2.10–2 g L−1) wasused to determine the sensitivity of the method. The twolast tubes contained medium prepared with seawaterfrom the port of Arcachon.
2.4. Chromosome preparation
Harvesting included blocking cell divisions atmetaphase using colchicine (50–60 μg/mL final con-centration) for 1 h 50, treatment with a hypotonicsolution (0.6% tri-sodium citrate) for 45 min, andfixation in ice-cold 3:1 methanol-acetic acid (fourchanges for a minimum of 2 h). For slide preparation,explants were dissociated with 45% acetic acid and theresulting cell suspension was then dropped from aPasteur pipette onto warm slides (42±0.5 °C) from aheight of 15 cm. Bromodeoxyuridine-incorporatedmetaphase-chromosomes were stained using the fluo-rescence plus Giemsa technique (FPG) as described byWolf and Perry (1974).
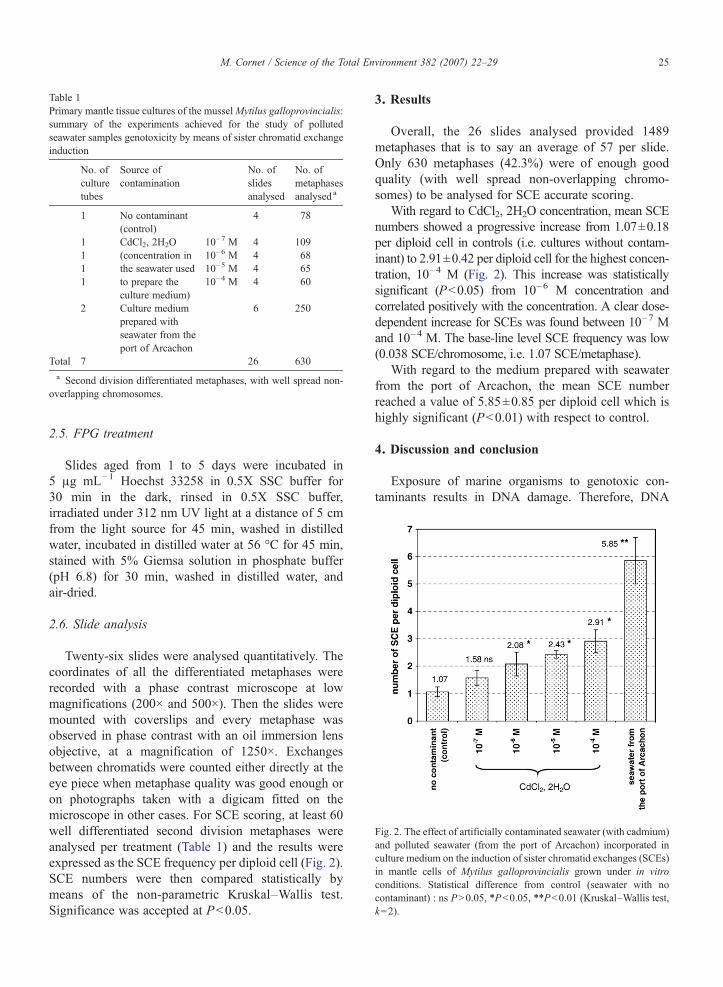
Fig. 2. The effect of artificially contaminated seawater (with cadmium)and polluted seawater (from the port of Arcachon) incorporated inculture medium on the induction of sister chromatid exchanges (SCEs)in mantle cells of Mytilus galloprovincialis grown under in vitroconditions. Statistical difference from control (seawater with nocontaminant) : ns PN0.05, ⁎Pb0.05, ⁎⁎Pb0.01 (Kruskal–Wallis test,k=2).
Table 1Primary mantle tissue cultures of the musselMytilus galloprovincialis:summary of the experiments achieved for the study of pollutedseawater samples genotoxicity by means of sister chromatid exchangeinduction
No. ofculturetubes
Source ofcontamination
No. ofslidesanalysed
No. ofmetaphasesanalysed a
1 No contaminant(control)
4 78
1 CdCl2, 2H2O(concentration inthe seawater usedto prepare theculture medium)
10–7 M 4 1091 10–6 M 4 681 10–5 M 4 651 10–4 M 4 60
2 Culture mediumprepared withseawater from theport of Arcachon
6 250
Total 7 26 630
a Second division differentiated metaphases, with well spread non-overlapping chromosomes.
25M. Cornet / Science of the Total Environment 382 (2007) 22–29
2.5. FPG treatment
Slides aged from 1 to 5 days were incubated in5 μg mL−1 Hoechst 33258 in 0.5X SSC buffer for30 min in the dark, rinsed in 0.5X SSC buffer,irradiated under 312 nm UV light at a distance of 5 cmfrom the light source for 45 min, washed in distilledwater, incubated in distilled water at 56 °C for 45 min,stained with 5% Giemsa solution in phosphate buffer(pH 6.8) for 30 min, washed in distilled water, andair-dried.
2.6. Slide analysis
Twenty-six slides were analysed quantitatively. Thecoordinates of all the differentiated metaphases wererecorded with a phase contrast microscope at lowmagnifications (200× and 500×). Then the slides weremounted with coverslips and every metaphase wasobserved in phase contrast with an oil immersion lensobjective, at a magnification of 1250×. Exchangesbetween chromatids were counted either directly at theeye piece when metaphase quality was good enough oron photographs taken with a digicam fitted on themicroscope in other cases. For SCE scoring, at least 60well differentiated second division metaphases wereanalysed per treatment (Table 1) and the results wereexpressed as the SCE frequency per diploid cell (Fig. 2).SCE numbers were then compared statistically bymeans of the non-parametric Kruskal–Wallis test.Significance was accepted at Pb0.05.
3. Results
Overall, the 26 slides analysed provided 1489metaphases that is to say an average of 57 per slide.Only 630 metaphases (42.3%) were of enough goodquality (with well spread non-overlapping chromo-somes) to be analysed for SCE accurate scoring.
With regard to CdCl2, 2H2O concentration, mean SCEnumbers showed a progressive increase from 1.07±0.18per diploid cell in controls (i.e. cultures without contam-inant) to 2.91±0.42 per diploid cell for the highest concen-tration, 10–4 M (Fig. 2). This increase was statisticallysignificant (Pb0.05) from 10–6 M concentration andcorrelated positively with the concentration. A clear dose-dependent increase for SCEs was found between 10–7 Mand 10–4 M. The base-line level SCE frequency was low(0.038 SCE/chromosome, i.e. 1.07 SCE/metaphase).
With regard to the medium prepared with seawaterfrom the port of Arcachon, the mean SCE numberreached a value of 5.85±0.85 per diploid cell which ishighly significant (Pb0.01) with respect to control.
4. Discussion and conclusion
Exposure of marine organisms to genotoxic con-taminants results in DNA damage. Therefore, DNA

26 M. Cornet / Science of the Total Environment 382 (2007) 22–29
integrity in marine organisms reflect the effects ofexposure to chemicals and is widely accepted as a goodindicator for environmental stress (Shugart, 2000). DNAalterations can be detected at the chromosomal level inthe form of aberrations, micronuclei or sister chromatidexchanges (Natarajan, 1993). In the present study, thesister chromatid exchange (SCE) frequency analysiswas used as genotoxic test system for this purpose.
In the absence of established cell lines with activeproliferation in aquatic invertebrates, in vitro SCEassays have been reported only in insects (Mukherjeeet al., 1986; Wei et al., 1993). However, in vivo, SCEassays have been developed for aquatic invertebrates,including platyhelminthes (Edelman and Lin, 1994),mussels (Dixon and Clarke, 1982; Dixon et al., 1985;Brunetti et al., 1986; Pasantes et al., 1996), and marineworms (Pesch et al., 1981; Jha et al., 1996; Hagger et al.,2002). In all these reports, SCE frequencies were foundto increase with higher xenobiotic levels.
The data presented in Fig. 2 show that the values arelow in comparison with those reported in the literature inembryo-larval stages (Dixon and Prosser, 1986) andadults (Dixon and Clarke, 1982) of Mytilus exposed toother sorts of contaminants at different concentrations.In in vitro studies, cadmium chloride induced astatistically significant increase in the frequency ofSCEs in mammalian chromosomes (Fahmy and Aly,2000). In human, Mourón et al. (2004) have shown thatsister chromatid exchanges assays frequencies weresignificantly increased in cells exposed to cadmium saltsin relation to controls. In the range of concentrationsused in the present study, it was demonstrated thatcadmium lead to DNA alteration in cell cultures ofrainbow trout and mice between 10–6 and 10–4 M(Risso-de Favernay et al., 2001; Biagioli et al., 2001). Agenotoxic effect of CdCl2 for a concentration of 10
–5 Mwas reported in an other species of bivalve, Pectenmaximus, by Le Pennec and Le Pennec (2001).
The analysis of SCEs induced by cadmium chloridein the present study showed DNA alterations even at thelowest concentration (i.e. 10–7 M). Of course, theexposure concentration employed in this study is not ofenvironmental relevance. Dissolved Cd concentrationsin the Gironde Estuary vary according to the salinity,ranging from 0.179 10–9 M for a salinity of 0.5 g L−1 to1.165 10–9 M for a salinity of 15.6 g L−1 (Kraepiel et al.,1997). Nevertheless, this sensitivity is in agreement withpreviously published values obtained for other contami-nants e.g. cyclophosphamide (Pesch et al., 1981) ormethyl methane sulfonate (Jha et al., 2000) in Mytilusedulis but the chemical compounds have different levelof toxicity related also to a given organism used.
The sister chromatid exchange assay detects theability of a chemical to enhance the exchange of DNAbetween two sister chromatids of a duplicating chromo-some. This technique is useful for studying the effect ofa wide range of environmental genotoxic agents withoutidentifying them singularly, (without the need of labor-intensive extraction procedures with expensive organicsolvents (Dayeh et al., 2002)) and can be used forassessing the toxicity of complex pollutant mixtures inthe marine environment. This is especially the case forthe port of Arcachon where the seawater tested here wassampled. The presence of polycyclic aromatic hydro-carbons (PAHs), polychlorinated biphenyls (PCBs),tributyltin (TBT) and heavy metals reported in the Bay(Devier et al., 2005) readily explain the SCE valueobtained (5.85 SCE per metaphase). PAHs and TBT areparticularly known to induce exchanges between sisterchromatids (Jha et al., 1996, 2000; Hagger et al., 2002).Mixtures of contaminants would have an additive effecton SCE frequency (Backhaus et al. 2003) but the kind ofeffect of a mixture of unknown compounds can't bepredicted.
The type of culture medium and the type of serummayinfluence the frequency of SCEs (Albertini et al., 2000). Inrainbow trout primary epithelial cell cultures, mediumcomposition have been found to have effect on the tox-icity of chemical substances (Dowling and Mothersill,2001). Lyons-Alcantara et al. (1996) have shown thatserum-free media are probably preferable than serum-containing media when testing pollutants in vitro becauseof the reduction of activity of free heavy metal ions due tobinding with serum components which significantly re-duce their bioavailability (Ni Shúilleabháin et al., 2006).Nevertheless, we showed recently that the technique ofculture developed, can alsoworkwithout serum but with areduction of the cellular proliferation (Cornet, 2006a).
In conclusion, the highlights from this procedure areas follows: the fundamental characteristic is the presenceof active cell proliferation in the culture which is anessential prerequisite since the detection of SCEs requireincorporation of BrdU into chromosomal DNA for twocell cycles. It is relevant to note that any other techniqueas well which needed an active cell proliferation such aschromosome aberrations or micronuclei (Kirsch-Volderset al., 2003) could be employed. Other important factorsthat may influence the success in demonstratinggenotoxic effects of contaminants include, first, the usewhole-water samples and thus the individual extractionof contaminants is not required (Dayeh et al., 2002).Second, provided a relative reduction of cell prolifera-tion, the culture technique can also work without serum(Cornet, 2006a) avoiding the reduction of heavy metal

27M. Cornet / Science of the Total Environment 382 (2007) 22–29
toxicity due to binding with the serum components assuggested by Lyons-Alcantara et al. (1996). Third, thereis no need of cell dissociation before culturing (Cornet,1993) avoiding tissue disaggregation resulting in celldamage and release of toxic enzymes such as proteases(Stacey and MacDonald, 2001). Fourth, a cellular prolif-eration can be maintained in cultures carried out in amedium containing seawater the salinity of which iscomprise between 28 and 35‰, providedMEMwas usedas basal medium (Cornet, 2006b) that ensure any ob-served toxicitywas due to contaminants and not as a resultof osmotic stress which is known to induce SCE in humanperipheral lymphocytes (Kroger and Nowak, 1990).
This work demonstrated the feasibility of the primarytissue culture system for the determination of genotoxi-city in seawater samples. Although the study presentedhere is only the first one approach in real size of theproposed technique, we know now that it can be inte-grated as a part of multi-biomarker processes such asthose that are developed in our laboratory for the detec-tion of biological effects of pollution, e.g. MRX protein(Achard et al., 2004), rpS9 gene expression (Achard-Joris et al., 2006). It is essential to be able of taking intoaccount in an early way the effects of contaminants onthe marine species and populations in order to act as fastas possible, before irreversible damage was caused. Forthat purpose, the analysis must concern the DNA aboveall other levels of organisation (e.g. physiology, mor-phology) (Brock, 1991; Steinert, 1999), since geneticalchanges may reflect a rapid response of organisms toenvironmental toxicant exposure and can provide earlywarning signs of adverse long-term effects in the pop-ulations of molluscs (Bolognesi et al., 1999; Baršienéand Baršyté Lovejoy, 2000).
Acknowledgements
Many thanks are due to Dr. Manuelle Maurice andDr. Thierry Burgeot (Ifremer). This work was supportedby a grant of the Institut Français de Recherche pourl'Exploitation de la Mer (Ifremer). This paper iscontribution number 1580 from UMR 5805 (CNRS),Université Bordeaux 1, Laboratoire d'OcéanographieBiologique, Observatoire Aquitain des Sciences del'Univers (OASU).
References
Achard M, Baudrimont M, Boudou A, Bourdineaud JP. Induction of amultixenobiotic resistance protein (MXR) in the Asiatic clamCorbicula fluminea after heavy metals exposure. Aquat Toxicol2004;67:347–57.
Achard-Joris M, Gonzalez P, Marie V, Baudrimont M, Bourdineaud JP.cDNA cloning and gene expression of ribosomal S9 protein genein the mollusk Corbicula fluminea: a new potential biomarker ofmetal contamination up-regulated by cadmium and repressed byzinc. Environ Toxicol Chem 2006;25:527–33.
Albertini RJ, Anderson D, Douglas GR, Hagmar L, Hemminki K,Merlo F, et al. IPCS guidelines for the monitoring of genotoxiceffects of carcinogens in humans. Mutat Res 2000;463:111–72.
Audry S, Blanc G, Schäfer J. Cadmium transport in the Lot-Garonneriver system (France) — temporal variability and a model for fluxestimation. Sci Total Environ 2004;319:197–213.
Backhaus T, Altenburger R, Arrhenius A, Blanck H, Faust M, FinizioA, et al. The BEAM-project: prediction and assessment of mixturetoxicities in the aquatic environment. Cont Shelf Res 2003;23:1757–69.
Baršiené J, Baršyté Lovejoy D. Environmental genotoxicity inKlaipéda Port Area. Int Rev Hydrobiol 2000;85:663–72.
Baudrimont M, Schäfer J, Marie V, Maury-Brachet R, Bossy C,Boudou A, et al. Geochemical survey and metal bioaccumulationof three bivalve species (Crassostrea gigas, Cerastoderma eduleand Ruditapes philippinarum) in the Nord Médoc salt marshes(Gironde estuary, France). Sci Total Environ 2005;337:265–80.
Biagioli M, Wätjen W, Beyersmann D, Zoncu R, Cappellini R, deRagghianti M, et al. Cadmium-induced apoptosis in murinefibroblasts is suppressed by Bcl-2. Arch Toxicol 2001;75:313–20.
Birmelin C, Mitchelmore CL, Goldfarb PS, Livingstone DR.Characterisation of biotransformation enzyme activities andDNA integrity in isolated cells of the digestive gland of thecommon mussel, Mytilus edulis L. Comp Biochem Physiol1998;120A:51–6.
Birmelin C, Pipe RK, Golfarb PS, Livingstone DR. Primary cell-culture of the digestive gland of the marine mussel Mytilus edulis:a time-course study of antioxydant- and biotransformation-enzymeactivity and ultrastructural changes. Mar Biol 1999;135:65–75.
Boisson F, Goudard F, Durand JP, Barbot C, Pieri J, Amiard JC, et al.Comparative radiotracer study of cadmium uptake, storage,detoxification and depuration in the oyster Crassostrea gigas:potential adaptative mechanisms. Mar Ecol Prog Ser 2003;254:177–86.
Bolognesi C, Landini E, Roggieri P, Fabbri R, Viarengo A.Genotoxicity biomarkers in the assessment of heavy metal effectsin mussels: experimental studies. Environ Mol Mutagen 1999;33:287–92.
Brock V. Genetic studies of the aquatic environment. Why and How.In: Hummel H, editor. The genetics of marine and estuarineorganisms. Proceedings of the first workshop of the Network ofEuropean Association of Marine Sciences and Techniques(EAMST) on the genetics of marine and estuarine organisms,28 February–2 March 1990, 251. Yerseke: The Netherlands;1991. p. 21–6.
Bruckmann E, Wojcik A, Obe G. Sister chromatid differentiation withbiotin-dUTP. Chromosom Res 1999;7:185–9.
Brunetti R, Gola I, Majone F. Sister-chromatid exchange in developingeggs of Mytilus galloprovincialis Lmk. (Bivalvia). Mutat Res1986;174:207–11.
Burgeot T, His E, Galgani F. The micronucleus assay in Crassostreagigas for the detection of seawater genotoxicity. Mutat Res1995;342:125–40.
Cornet M. A short-term culture method for chromosome preparationfrom somatic tissues of adult mussel (Mytilus edulis). Experientia1993;49:87–90.

28 M. Cornet / Science of the Total Environment 382 (2007) 22–29
Cornet M. A study of some factors affecting the efficiency of in vitromussel tissue culture for chromosome preparation. Cytobios1995;84:7–12.
Cornet M. Obtaining cell proliferation for chromosome preparation ingill tissue culture of the oyster Crassostrea gigas. Cytotechnology2000;32:1–7.
Cornet M. Primary mantle tissue culture from the bivalve molluscMytilus galloprovincialis: investigations on the growth promotingactivity of the serum used for medium supplementation. JBiotechnol 2006a;123:78–84.
Cornet M. Effects of seawater salinity fluctuations on primary tissueculture from the mussel Mytilus galloprovincialis. Potentialapplication to the detection of seawater genotoxicity. Toxicol InVitro 2006b;20:1500–5.
Cossa D. A review of the use ofMytilus spp. as quantitative indicatorsof cadmium and mercury contamination in coastal waters. OceanolActa 1989;12:417–32.
Dayeh R, Schirmer K, Bols NC. Applying whole-water samplesdirectly to fish cell cultures in order to evaluate the toxicity ofindustrial effluent. Water Res 2002;36:3727–38.
Depledge MH. The ecotoxicological significance of genotoxicity inmarine invertebrates. Mutat Res 1998;399:109–22.
DevierMH,Augagneur S, BudzinskiH, LeMenachK,Mora P,NarbonneJF, et al. One-year monitoring survey of organic compounds (PAHs,PCBs, TBT), heavy metals and biomarkers in blue mussels from theArcachon Bay, France. J Environ Monit 2005;7:224–40.
Dixon DR, Clarke KR. Sister chromatid exchange — a sensitivemethod for detecting damage caused by exposure to environmentalmutagens in the chomosomes of adult Mytilus edulis. Mar BiolLett 1982;3:163–72.
Dixon DR, Prosser H. An investigation of the genotoxic effects of anorganotin antifouling compound (bis(tributyltin) oxide) on thechromosomes of the edible mussel, Mytilus edulis. Aquat Toxicol1986;8:185–95.
Dixon DR, Wilson JT. Genetics and marine pollution. Hydrobiologia2000;420:29–43.
Dixon DR, Jones IM, Harrison FL. Cytogenetic evidence of inducibleprocesses linked with metabolism of a xenobiotic chemical in adultand larval Mytilus edulis. Sci Total Environ 1985;46:1–8.
Domart-Coulon I, Auzoux-Bordenave S, Doumenc D, Khalanski M.Cytotoxicity assessment of antibiofouling compounds and by-products in marine bivalve cell cultures. Toxicol In Vitro2000;14:245–51.
Dowling K, Mothersill C. The further development of rainbow troutprimary epithelial cell cultures as a diagnostic tool in ecotoxicol-ogy risk assessment. Aquat Toxicol 2001;53:279–89.
Edelman JR, Lin YJ. Differential staining of Dugesia tigrina sisterchromatids. Cytobios 1994;78:123–8.
FahmyMA, Aly FA. In vivo and in vitro studies on the genotoxicity ofcadmium chloride in mice. J Appl Toxicol 2000;20:231–8.
Ferro M, Doyle A. Standardisation for in vitro toxicity tests. Cell BiolToxicol 2001;17:205–12.
Freshney I. Application of cell cultures to toxicology. Cell BiolToxicol 2001;17:213–30.
Grousset FE, Jouanneau JM, Castaing P, Lavaux G, Latouche C. A70 year record of contamination from industrial activity along theGaronne river and its tributaries (SW France). Estuar Coast ShelfSci 1999;48:401–14.
Hagger JA, Fisher AS, Hill SJ, Depledge MH, Jha AN. Genotoxic,cytotoxic and ontogenic effects of tri-n-butyltin on the marineworm, Platynereis dumerilii (Polychaeta: Nereidae). AquatToxicol 2002;57:243–55.
Jha AN, Hutchinson TH, Mackay JM, Elliott BM, Dixon DR.Development of an in vivo genotoxicity assay using the marineworm Platynereis dumerilii (Polychaeta: Neridae). Mutat Res1996;359:141–50.
Jha AN, Cheung VV, Foulkes ME, Hill SJ, DepledgeMH. Detection ofgenotoxins in the marine environment: adoption and evaluation ofan integrated approach using embryo-larval stages of the marinemussel, Mytilus edulis. Mutat Res 2000;464:213–28.
Kanaar R, Hoeijmakers JHJ, Van Gent DC. Molecular mechanisms ofDNA double-strand break repair. Trends Cell Biol 1998;8:483–9.
Kirsch-Volders M, Sofuni T, Aardema M, Albertini S, Eastmond D,Fenech M, et al. Report from the in vitro micronucleus assayworking group. Mutat Res 2003;540:153–63.
Kraepiel AML, Chiffoleau JF, Martin JM, Morel FMM. Geochemistryof trace metals in the Gironde estuary. Geochim Cosmochim Acta1997;61:1421–36.
Kroger I, Nowak C. Induction of sister chromatid exchanges with hy-potonic treatment. Teratog Carcinog Mutagen 1990;10:295–305.
Le Pennec G, Le Pennec M. Evaluation of the toxicity of chemicalcompounds using digestive acini of the bivalve mollusc Pectenmaximus L. maintained alive in vitro. Aquat Toxicol 2001;53:1–7.
Le Pennec G, Le Pennec M. Induction of glutathione-S-transferase inprimary cultured digestive gland acini from the mollusk bivalvePecten maximus (L.) : application of a new cellular model inbiomonitoring studies. Aquat Toxicol 2003;64:131–42.
Livingstone DR, Chipman JK, Lowe DM, Minier C, Mitchelmore CL,Moore MN, et al. Development of biomarkers to detect the effectsof organic pollution on aquatic invertebrates : recent molecular,genotoxic, cellular and immunological studies on the commonmussel (Mytilus edulis) and other mytilids. Int J Environ Pollut2000;13:56–91.
Lyons-Alcantara M, Tarazona JV, Mothersill C. The differential effectsof cadmium exposure on the growth and survival of primary andestablished cells from fish and mammals. Cell Biol Toxicol1996;12:29–38.
Mitchelmore CL, Birmelin C, Livingstone DR, Chipman JK.Detection of DNA strand breaks in isolated mussel (Mytilusedulis) digestive gland cells using the "Comet" assay. EcotoxEnviron Saf 1998;41:51–8.
Mourón SA, Grillo CA, Dulout FN, Golojow CD. A comparativeinvestigation of DNA strand breaks, sister chromatid exchangesand K-ras gene mutations induced by cadmium salts in culturedhuman cells. Mutat Res 2004;568:221–31.
Mukherjee AB, Herrera RJ, Luckett DC. Spontaneous and inducedsister-chromatid exchanges in cultured mosquito cells. Cytobios1986;46:181–7.
Natarajan AT. Mechanisms for induction of mutations and chromo-some alterations. Environ Health Perspect 1993;101S3:225–9.
Ní Shúilleabháin S, Mothersill C, Sheehan D, O'Brien NM,O'Halloran J, van Pelt FNAM, et al. Cellular responses in primaryepidermal cultures from rainbow trout exposed to zinc chloride.Ecotoxicol Environ Saf 2006;65:332–41.
Pasantes JJ, Martinez-Exposito MJ, Torreiro A, Mendez J. The sisterchromatid exchange test as an indicator of marine pollution : somefactors affecting SCE frequencies in Mytilus galloprovincialis.Mar Ecol Prog Ser 1996;143:113–9.
Pesch GG, Pesch CE, Malcolm AR. Neanthes arenaceodentata, acytogenetic model for marine genetic toxicology. EnvironMutagen 1981;3:386–7.
Pruski AM, Dixon DR. Effect of cadmium on nuclear integrity andDNA repair efficiency in gill cells of Mytilus edulis L. AquatToxicol 2002;57:127–37.

29M. Cornet / Science of the Total Environment 382 (2007) 22–29
Risso-de Favernay C, Devaux A, Lafaurie M, Girard JP, Bailly B,Rahmani R. Cadmium induces apoptosis and genotoxicity inrainbow trout hepatocytes through generation of reactive oxygenespecies. Aquat Toxicol 2001;53:65–76.
Rizki M, Kossatz E, Creus A, Marcos R. Genotoxicity modulation bycadmium treatment : studies in the Drosophila wing spot test.Environ Mol Mutagen 2004;43:196–203.
Schäfer J, Blanc G, Lapaquellerie Y, Maillet N, Maneux E, Etcheber H.Ten-year observation of the Gironde tributary fluvial system:fluxes of suspended matter, particulate organic carbon andcadmium. Mar Chem 2002;79:229–42.
Shugart LR. DNA damage as a biomarker of exposure. Ecotoxicology2000;9:329–40.
Shugart LR, Theodakaris CW. Environmental genotoxicity : probingthe underlying mechanisms. Environ Health Perspect 1994;102:13–7.
Stacey G, MacDonald C. Immortalization of primary cells. Cell BiolToxicol 2001;17:231–46.
Steinert SA. DNA damage as a bivalve biomarker. Biomarkers1999;4:492–6.
Theodorakis CW. Integration of genotoxic and population geneticendpoints in biomonitoring and risk assessment. Ecotoxicology2001;10:245–56.
Wei LY, Hou RF, Jan KY,Wang TC. Factors affecting zero backgroundfrequency of sister-chromatid exchange in mosquito cells.Mutagenesis 1993;8:179–82.
Wojcik A, Bruckmann E, Obe G. Insights into the mechanisms of sisterchromatid exchange formation. Cytogenet Genome Res 2004;104:304–309.
Wolff S, Perry P. Differential giemsa staining of sister chromatids andthe study of sister chromatid exchanges without autoradiography.Chromosoma 1974;48:341–53.
Zaroogian G, Anderson S. Comparison of cadmium, nickel and benzo(a)pyrene uptake into cultured brown cells of the hard shell clam,Mercenaria mercenaria. Comp Biochem Physiol 1995;111C:109–16.





























