Embed Size (px)
Citation preview

Review
Evolution and cell biology of dopamine receptors in vertebrates
Sophie Callier, Marina Snapyan, Stéphane Le Crom, Delphine Prou,Jean-Didier Vincent, Philippe Vernier *
Développement, évolution, plasticité du système nerveux, UPR 2197, Institut de Neurobiologie Alfred Fessard, CNRS, Gif-sur-Yvette, France
Received 30 June 2003; accepted 2 July 2003
Abstract
Dopamine, one of main modulatory neurotransmitters of the nervous system acts on target cells through two classes of G protein-coupledreceptors, D1 and D2. The two dopamine receptor classes display different structures, interact with different regulatory partners (includingheterotrimeric G proteins) and, accordingly, have independent evolutionary origins. In vertebrates, each of these receptor classes comprisesseveral subtypes, generated by two steps of gene duplications, early in vertebrate evolution. In the D1 receptor class, the D1A, D1B, D1C andD1D subtypes, and in the D2 class, the D2, D3 et D4 receptor subtypes have been conserved in most vertebrate groups. This conservation hasbeen driven by the acquisition, by each receptor subtype, of a small number of specific properties, which were selected for adaptive purposein vertebrates. Among these properties, affinity for dopamine, the natural ligand, intrinsic receptor activity, and agonist-induced desensitiza-tion clearly distinguish the receptor subtypes. In addition, each dopamine receptor subtype is addressed to a specific location within neuronalnetworks, although detailed information is lacking for several receptor subtypes. Receptors localization at diverse subcellular places inneurons may also differ from one subtype to another, resulting in different ways of regulating cell signalisation. One challenge for futureresearch on dopamine and its receptors would be to identify the nature of the protein partners and the molecular mechanisms involved inlocalizing receptors to the neuronal plasma membrane. In this respect, the evolutionary approach we have undertaken suggests that, due to geneduplications, a reasonable degree of freedom exists in the tight organisation of dopamine receptors in neurons. This “evolvability” of dopaminesystems has been instrumental to adapt the vertebrate species to nearly all the possible environments.
© 2003 Éditions scientifiques et médicales Elsevier SAS. All rights reserved.
Keywords: Receptor trafficking; Phylogeny; Desensitization; Cell signalling; Oligomerization
In Vertebrates, dopamine is a neurotransmitter acting as amodulator of neuronal activity, regulating thereby many dif-ferent functions in the central nervous system. This modula-tory action applies to sensory perception in the retina andolfactory bulb, regulation of prolactine release in the pitu-itary gland, control of body temperature, food intake andsexual behavior in the hypothalamus, tuning of sensori-motor cues in the basal ganglia. In addition, dopamine sys-tems are central to the maintenance and expression of thequalitative values of novelty in life experiences and, thus,motivation or aversion.
The various effects of dopamine on physiological systemsor organs are mediated by multiple metabotropic membranereceptors coupled to heterotrimeric G proteins (GPCR). As
much as seven receptor proteins encoded by different geneshave been isolated so far in vertebrates. The interest fordopamine receptors arose from the discovery of alteration indopamine transmission in several human pathologies, essen-tially Parkinson’s disease, addiction to drugs of abuse, disor-ders of mood and affect (schizophrenia), prolactinomas ofthe anterior pituitary gland, hypertension or cardiovascularfailure. Accordingly, dopamine receptors are the targets ofmajor drug classes (neuroleptics, antiparkinsonian drugs,etc. (Strange, 1993; Seeman and Van Tol, 1994; Sokoloff andSchwartz, 1995).
The fact that dopamine is able to act on target cells viaseveral different receptor molecules is common to most neu-rotransmitters, but the necessity for such a molecular diver-sity, as well as its physiological consequences are far fromobvious. This is a frequently asked question in the field ofmolecular biology and genetics. Dopamine receptor sub-types share similar sequences, structures and functions. Ac-
* Corresponding author: DEPSN, UPR 2197, Institut de NeurobiologieA. Fessard, CNRS, Avenue de la Terrasse, 91198 Gif-sur-Yvette cédex,France. Tel: 33–169 82 41 88; Fax: 33–169 82.
E-mail address: [email protected] (P. Vernier).
Biology of the Cell 95 (2003) 489–502
www.elsevier.com/locate/bicell
© 2003 Éditions scientifiques et médicales Elsevier SAS. All rights reserved.doi:10.1016/S0248-4900(03)00089-3

cordingly, they are encoded by a gene family, the members ofwhich have been generated by events of gene duplications,followed by selection of the duplicated genes (Vernier et al.,1995). Therefore, understanding how the genetic multiplicityof G protein coupled receptors has been created, and which isits functional counterpart, can only be achieved through anevolutionary approach (Kirschner and Gerhart, 1998). In-deed, diversification of dopamine receptors is primarily anhistorical phenomenon, and a better knowledge of this his-tory is a prerequisite to any experimental analysis of thestructures and functions of dopamine receptors present indifferent animal species. On the one hand, resemblances ingene structures highlight the conservation of fundamentalfunctions of dopamine receptors in organisms. On the otherhand, differences rely on the occurrence of discrete or largemutational events that led to modify tissue-specific geneexpression and function of corresponding proteins. Thesedifferences have been selected for adaptive purpose all alongevolution. Comparative analysis that brings together molecu-lar data and adaptive physiological features is more and morenecessary to build an integrated view of dopamine receptors,from molecular neurobiology to animal behaviour.
1. A brief history of the evolution of dopamine receptorgenes in vertebrates
In vertebrates, dopamine receptors belong to two classesof G protein-coupled receptors. The classes of dopaminereceptor were initially defined by their ability to modulateadenylyl cyclase activity and cAMP accumulation in cells(Kebabian and Calne, 1979). D1 receptors are able to acti-vate, whereas dopamine D2 receptors decrease adenylyl cy-clase activity. However, it is now established that the D2
receptors are mainly responsible for modulating the activityof voltage-sensitive Ca++ and K+ channels. Lately, manypharmacological compounds were used to discriminate thesetwo receptor classes, establishing the concept of multiplereceptors for dopamine (reviewed in Sokoloff and Schwartz,1995; Missale et al., 1998).
As for most other neurotransmitter receptors, molecularcloning confirmed that D1 and D2 receptors constituted twodifferent classes of dopamine receptors. In mammals, tworeceptor subtypes have been assigned to the D1 receptor class(D1A/D1 and D1B/D5), but other subtypes exist in nonmam-malian vertebrates (D1C and D1D; reviewed in Kapsimali etal., 2003). Similarly, three subtypes of D2 receptors havebeen isolated in jawed vertebrates (D2, D3, D4). The overallprotein structure and topology deduced from receptor se-quences reveals major differences between the two classes ofdopamine receptors. The D2 receptors have a long thirdcytoplasmic loop and a short cytoplasmic C–terminal end,whereas D1 receptors exhibit a shorter third cytoplasmic loopbut a very long C terminal tail. These protein segments areinvolved in coupling with heterotrimeric G protein, and ac-cordingly, D1 and D2 receptors interact with different classesof heterotrimeric G proteins. The D1 receptors are coupled to
the Gs/Golf class of Ga proteins, whereas the D2 receptorscoupled essentially to Gi/Go proteins, therby modulatingdifferent intracellular signalling pathways. These facts leadto regard the two classes of dopamine receptors as verydistinct, except they are both able to bind dopamine as neu-rotransmitter. In addition, the existence of multiple receptorsubtypes, as found in vertebrates, cannot be understood with-out knowing more of their evolutionary origin and physi-ological roles.
The question of the evolutionary origin of the dopaminereceptor subtypes in vertebrates can been addressed by ana-lyzing the molecular phylogeny of the corresponding genes.When dopamine receptors are compared to other monoaminereceptors (adrenergic a1, a2, b, or trace amines, or serotoner-gic 5HT1, 5HT2, 5HT4, 5HT5, 5HT6, 5HT7), molecular phy-logenies clearly indicate that D1 and D2 receptor classes areunrelated, or at least not more closely related to each otherthan they are to other classes of monoamine receptors(Fig. 1). This is consistent with the distinct structural charac-teristics and functional properties of the D1 and D2 dopaminereceptors. It implicates that they have acquired indepen-dently and convergently the ability to bind dopamine, as inthe case of the other classes of monoamine receptors.
However, if molecular phylogeny highlights the structuralrelationships of dopamine receptors, it cannot provide, alone,robust hypotheses on the duplications and evolution ofdopamine receptors in the vertebrate phylum. With this per-spective in mind, the whole complement of dopamine recep-tors has been isolated in species belonging to groups ofvertebrates diverging at different times of vertebrate evolu-tion. The D1 receptors having no intron, all the members ofthe gene family can be isolated rather easily from genomicDNA. A similar approach is more difficult for the D2 recep-tors, the genes of which bear several large introns inside theirsequence.
In jawless vertebrates (agnathans), only one D1–like se-quence has been found in lampreys and in hagfish. It is thusvery likely that only one type of D1 receptor exist in thesespecies that descend from the earliest diverging vertebrates.Interestingly, these receptor sequences are not related to anyof the known D1 receptor sequences from jawed vertebrates.Thus, they probably diverged from the ancestor of the verte-brate D1 receptor before the occurrence of gene duplicationsgenerating the dopamine receptor subtypes found in jawedvertebrates (gnathostomes; Fig. 1). Accordingly, from carti-laginous fish (the electric ray Torpedo marmorata) to am-phibians (the toad Xenopus lævis), three different D1 receptorsequences have been found (Sugamori et al., 1994). In thesespecies, the phylogenetic tree easily depicts three subtypes,namely, D1A, D1B and D1C. In teleost fish (European eel,puffer fish, medaka, zebrafish) extra D1 receptor sequencesare found (Cardinaud et al., 1997), probably correspondingto gene duplication events that occurred specifically in te-leost fish (Aparicio, 2000).
In amniotes, the situation is even more complicated(Fig. 1). A fourth D1 receptor-related sequence has been
490 S. Callier et al. / Biology of the Cell 95 (2003) 489–502
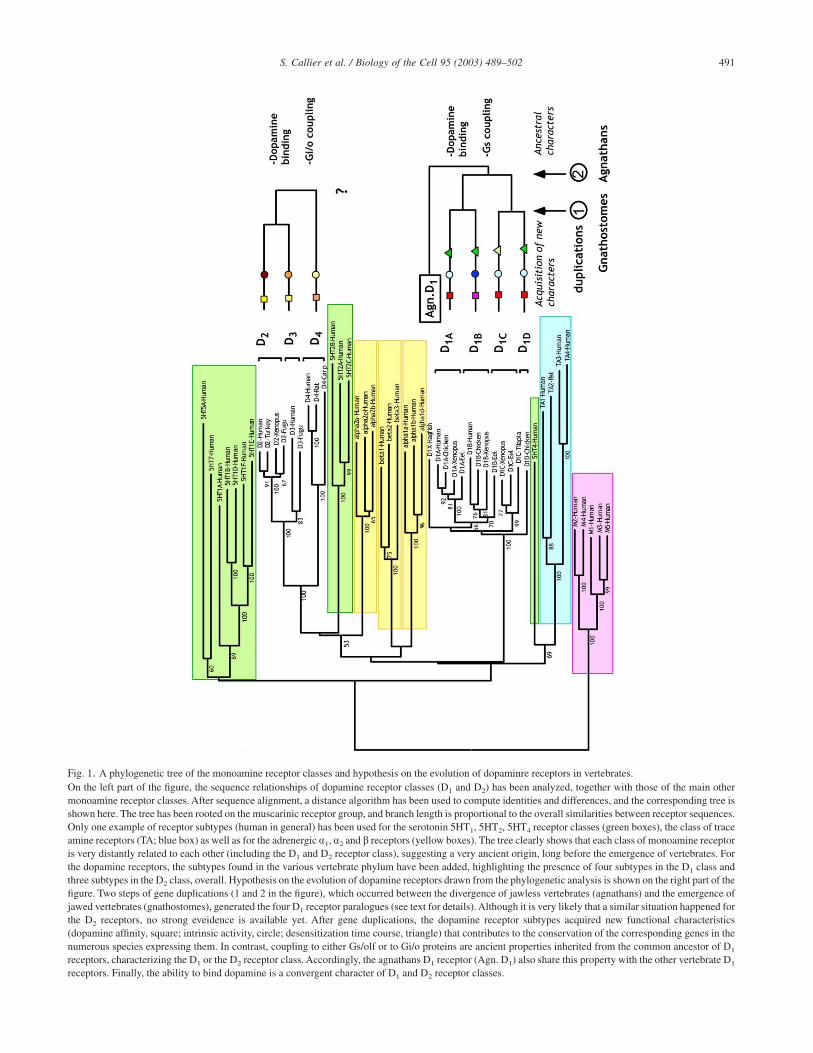
Fig. 1. A phylogenetic tree of the monoamine receptor classes and hypothesis on the evolution of dopaminre receptors in vertebrates.On the left part of the figure, the sequence relationships of dopamine receptor classes (D1 and D2) has been analyzed, together with those of the main othermonoamine receptor classes. After sequence alignment, a distance algorithm has been used to compute identities and differences, and the corresponding tree isshown here. The tree has been rooted on the muscarinic receptor group, and branch length is proportional to the overall similarities between receptor sequences.Only one example of receptor subtypes (human in general) has been used for the serotonin 5HT1, 5HT2, 5HT4 receptor classes (green boxes), the class of traceamine receptors (TA; blue box) as well as for the adrenergic a1, a2 and b receptors (yellow boxes). The tree clearly shows that each class of monoamine receptoris very distantly related to each other (including the D1 and D2 receptor class), suggesting a very ancient origin, long before the emergence of vertebrates. Forthe dopamine receptors, the subtypes found in the various vertebrate phylum have been added, highlighting the presence of four subtypes in the D1 class andthree subtypes in the D2 class, overall. Hypothesis on the evolution of dopamine receptors drawn from the phylogenetic analysis is shown on the right part of thefigure. Two steps of gene duplications (1 and 2 in the figure), which occurred between the divergence of jawless vertebrates (agnathans) and the emergence ofjawed vertebrates (gnathostomes), generated the four D1 receptor paralogues (see text for details). Although it is very likely that a similar situation happened forthe D2 receptors, no strong eveidence is available yet. After gene duplications, the dopamine receptor subtypes acquired new functional characteristics(dopamine affinity, square; intrinsic activity, circle; desensitization time course, triangle) that contributes to the conservation of the corresponding genes in thenumerous species expressing them. In contrast, coupling to either Gs/olf or to Gi/o proteins are ancient properties inherited from the common ancestor of D1
receptors, characterizing the D1 or the D2 receptor class. Accordingly, the agnathans D1 receptor (Agn. D1) also share this property with the other vertebrate D1
receptors. Finally, the ability to bind dopamine is a convergent character of D1 and D2 receptor classes.
491S. Callier et al. / Biology of the Cell 95 (2003) 489–502

isolated from chicken and crocodiles (archosaurs) and namedD1D (Demchyshyn et al., 1995). This subtype is also presentin lepidosaurs (lizzards and snakes). However, the thirdgroup of amniotes, turtles, expresses D1A and D1B receptorsequences, but they possess only one more sequence dis-tantly related to the D1C subtype. In mammals, only D1A andD1B receptor-related sequences have been isolated in euth-erian (placental) mammals, as well as in metatherians (mar-supials). Interestingly, in the two main species of protothe-rian mammals, platypus and echidnea, two D1 receptorsequences have also been found, but they are related to theD1A and D1D subtypes. The most parsimonious interpretationof these data is that four different subtypes of D1–relatedreceptors existed in stem amniotes. Two of them have disap-peared in mammals, D1B and D1C in prototherian mammalsand D1C and D1D in the other groups of mammals. In otherwords, mammals are an exception among vertebrates sincethe D1C receptor subtype is present in all the other groups ofjawed vertebrates, and the D1D subtype is present in all theamniotes, with the exception of placental and marsupialmammals.
A detailed analysis of the molecular phylogeny of the D2
receptor class is not possible yet. However, based on currentdata, the phylogenetic picture and the evolutionary history ofthe D2–like receptors is very similar to that of the D1 receptorclass. Again, three subtypes are easily identified, (D2, D3 andD4 subtypes; Fig. 1), and they are found in most groups ofjawed vertebrate. Whether one or several D2–like receptorsexist in agnathans is not known at present, and we cannotwrite a comprehensive history of the D2 receptor class yet.
Overall, the existence of one subtype of dopamine recep-tor in agnathans and of two, three or four paralogous recep-tors in gnathostomes, supports the hypothesis of a doublewhole-genome duplication in the evolution of Craniates.However, it is impossible to know precisely when theseduplications took place, except they should have occurredbetween the emergence of the earliest vertebrates and that ofgnathostomes (Fig. 1). After these two gene duplications thatgave birth to four dopamine receptor paralogues, severalgroups of species have lost one or two of these genes. Forexample, bony fish and amphibians have lost one D1 receptorsequence (D1D), and the mammals have lost two of thesegenes (D1C and either D1B or D1D).
The D1A receptor exhibits the most conserved sequenceand it is the only one found in all the jawed vertebratespecies. It may thus be functionally the most important of thedopamine receptors, a statement clearly confirmed by theresults of specific knock-out of the D1–related genes inmouse (Sibley, 1999). In contrast, other dopamine receptorsequences are relatively more divergent (apparent “evolu-tionary speed”), as shown by a “long branch” in the phyloge-netic tree. This is for example the case for the D4 receptorsubtype in the D2 class, and for the D1C and D1D subtype inthe D1 class. This “long-branch” effect conceals the “true”evolutionary relationships of the D1 receptors in vertebrates(see Fig. 1) since a fast evolving sequence will branch deep in
the tree, although they have emerged late during evolution.Such a biased relationship can only be detected when the allthe paralogous and orthologous genes have been isolated fora large sample of species, as it is the case for the dopamine D1
receptors.The molecular analysis of sequence relatedness (molecu-
lar phylogenies) cannot tell why some receptor sequenceshave been conserved and why others have been lost. Acqui-sition of a novel functional character or a novel expressionterritory, are means by which duplicated genes gain non-redundant characters and were conserved in ancestral spe-cies. The following paragraphs give an overview of ourcurrent knowledge of the functional properties of dopaminereceptors.
2. Tissue distribution and subcellular localizationof dopamine receptors
A fundamental aspect of dopamine function in wholeorganism is the localization of dopamine receptors in thevarious areas of the nervous system or at the periphery of thebody. From a genetic point of view, as soon as two receptorsare differentially expressed in cells, they are no longer redun-dant, provided the cells remain submitted to the action of thetransmitter. Similarly, the precise subcellular localization ofreceptor within the target cells are major features of receptorfunction, especially in highly polarized cells such as neurons.Obviously, a receptor located at a post-synaptic position onthe terminal dendrites will not play the same role as a presyn-aptic receptor located on nerve terminals. Differential ex-pression or localization of receptors often occurs after geneduplication, either by acquisition of “new” expression terri-tories or by splitting the expression areas of the ancestralgene (Aparicio, 2000).
2.1. General features of dopamine receptor distribution inthe vertebrate nervous system
A comparative analysis of dopamine receptors has beencarried out in several important groups of vertebrates includ-ing teleost fish (Kapsimali et al., 2000), xenopus (unpub-lished results), birds (Schnell et al., 1999) and mammals(Tiberi et al., 1991; Meador-Woodruff et al., 1996; reviewedin Kapsimali et al., 2003). The D1 and D2 receptors arepresent in all of the known target areas of dopamine in thecentral nervous system of vertebrates and their expressionterritories exhibit considerable overlap. However the ques-tion about whether the different receptor subtypes are presentor not in the same cells has not been definitely resolved(Gerfen, 2000). Mostly, however, neurons are not expressingsimultaneously D1 and D2 receptors, or only at very differentlevels, revealing that the transcriptional regulation of the twodopamine receptor classes is essentially non redundant.
The overall distribution of dopamine receptors in the cen-tral nervous system is probably conserved in many vertebratespecies, since, as far as we know, the distribution pattern of
492 S. Callier et al. / Biology of the Cell 95 (2003) 489–502

the D1 and D2 receptor subtypes is generally similar in fish,amphibians birds and mammals. A summary of this tissuedistribution is given in Table 1. The distribution of the three,well-studied paralogous D1 and D2 receptor subtypes dis-plays a picture of large overlap, differential abundance, witha few specific localizations. This overview indicates that,after gene duplications, the new paralogous genes have con-served, to a large extent, a similar regulation of tissue-specific expression. However, the few –but significant– dif-ferences that exist between subtypes, certainly contributed tothe conservation of the duplicated genes at the origin ofjawed vertebrates and were crucial in setting up many of theexpression characteristics of dopamine receptors found inmodern vertebrates.
Among the D1 receptors, the D1A receptor subtype is themost abundant, as well as the D2 receptor subtype in the D2
class. This holds true in all the vertebrate species examinedso far. In contrast, the D1C receptor, which is not found inmammals, is only weakly expressed in the fish and amphib-ian brain and only in limited areas of the hypothalamus. TheD1B receptor subtype is also less abundant than the D1A
receptor in mammals, birds, amphibians and fish, but it isspecifically present in important functional areas such as thehippocampus. This characteristic fits well with the higherconservation of D1A sequences as compared to D1B or D1C
sequences in vertebrates. Similarly, in the D2 class, the D3
and D4 receptors are by far much less abundant in the brain,as compared to D2 receptors. Moreover, the distribution ofthe D4 receptor transcript strikingly overlaps with that of the
D2 receptor, in the dorsal striatum and cortical areas forexample, suggesting a large degree of redundancy betweenthe two receptor subtypes. Accordingly, the sequence D4
subtype is rapidely diverging, and null mutations are knownin human without phenotypic consequences (Nothen et al.,1994). The D3 receptor is, on average, expressed at a lowerlevel than the D2 receptor subtype, but it is found in areaswhere no or very little D2 receptors are present, such as in theislands of Calleja and in the nucleus accumbens . In this lattercase, it is clearly co-express with the D1 receptor, suggestingspecific interactions between the two receptors (Sokoloff etal., 2003).
These observations have important implications to under-stand how the various subtypes of the D1 or D2 receptors haveevolved after the duplication of the ancestral genes in verte-brates. The similarities of the expression territories of the D1
receptor genes implicates that the main cis–regulatory ele-ments controlling tissue-specific transcription of genes havebeen conserved after the gene duplication process. However,a few -but significant– differences are observed among re-gional distributions of the D1 subtypes, probably sustainingenough functional relevance in order to be conserved.
2.2. Subcellular localization of dopamine receptors
Recent investigations have pointed out that dopamine re-ceptor subtypes exhibit discrete subcellular localizations
Table 1Distribution of dopamine receptors in vertebrate brains:The table reports in a semi-quantitative manner the relative tissue distribution of the various dopamine receptor subtypes. This relative distribution is wellconserved for all the species of gnathostomes that have been analyzed so far
Brain region
Dopamine receptorsD1like D2 likeD1A/D1 D1B/D5 D2 D3 D4
Cortex + + + + +Hippocampus – ++ + + +Basal gangliaStriatum ventral ++ + ++ ++ +Striatum dorsal ++ + ++ +Nucleus accumbens ++ ++ + ++Olfactory tubercle + – ++ +Globus pallidus + + + +Ventral tegmental area + – +Substantia nigra + + ++ + +Midbrain and BrainstemNucleus tractus solitarius ++ + ++Optic tract +Vegetative areas of spinal cord? – +Thalamus + ++ + +HypothalamusDorsal hypothalamus + +Ventral hypothalamus + – + + +Hypophyse – – ++ –Amydaloid complex ++ + + + +Retina + + + ++
++ high expression, + low expression, – not detected
493S. Callier et al. / Biology of the Cell 95 (2003) 489–502

within neuronal networks. A detailed analysis has been madefor the D1A/D1, the D1B/D5 and the D2 receptor subtypesonly, due to the availability of good antibodies. Both D1A/D1
and D2 receptors are located at a post-synaptic location indopamine-target neurons in the mammalian brain. For in-stance, electron microscopy analysis has shown that, withinthe pyramidal cells of the primate cortex, the D1 receptor isexpressed predominantly on dendritic spines, whereas theD1B/D5 receptor is found primarily on dendritic shafts (Smi-ley et al., 1994; Bergson et al., 1995). In the rat striatum, D2
receptors are more concentrated on spiny dendrites and spineheads of the medium spiny neurons than on the plasmamembrane of the somata (Levey et al., 1993). Similarly, inthe olfactory bulb, the D2 receptor is found mainly on thedistal part of dendrites and in the spines (Levey et al., 1993;Yung et al., 1995).
In addition to this typical post-synaptic location, both theD1A/D1 and the D2 receptor subtypes have been found atpresynaptic sites within axon terminals, although typicallynot within the same projection pathway (Huang et al., 1992;Levey et al., 1993; Smiley et al., 1994). In the dopaminergicneurons of the nigrostriatal and tegmental ventral area, D2
receptors are highly concentrated in the distal region ofdendrites and proximal part of dendrites, but also on thenerve terminals located far away from the cell bodies in thestriatum. Interestingly, the mammalian D2 receptor exists intwo isoforms generated by the alternative splicing of a smallregion corresponding to the third cytoplasmic loop of thereceptor structure. Semi-quantitative PCR and immunohis-tochemical studies showed that the short D2S receptor pre-dominates in the cell bodies and projection axons of dopam-inergic neurons of the mesencephalon and hypothalamus(Guivarch et al., 1995). In contrast, the long D2L receptorisoform is more highly expressed in the neurons of striatumand nucleus accumbens (Khan et al., 1998), suggesting thatthe differential splicing of the D2 receptor may result inpreferential subcellular localization within neurons. Thesestudies also revealed an unusual localization of D2 receptorsin perinuclear and other intracellular membrane compart-ments, probably the endoplasmic reticulum (ER).
This observation agrees with a detailed analysis of thesubcellular localization of the mammalian dopamine D2 re-ceptor carried out in various cell types (Prou et al. 2001).Indeed, after cell transfection, most of the newly synthesizedreceptors accumulate in large intracellular compartments,whereas the plasma membrane was only weakly labelled.Double-labelling experiments showed that the D2 receptorwas mostly retained in the ER, the long isoform morestrongly than the short one. This retention is accompanied bya striking vacuolization of the ER, roughly proportional tothe expression levels of the two receptor isoforms. Thisphenomenon seems to be a consequence of the high intrinsicactivity of the D2 receptor, promoting activation of heterotri-meric G protein inside the secretory pathway and blockingfurther transport in the ER. Such a mechanism may contrib-
ute to the regulation of the intracellular trafficking of the D2
receptors isoforms (Prou et al., 2001).However, the mechanisms involved in transport, sorting
and targeting of dopamine receptors subtypes to discretesubcellular locations remain almost entirely unknown. Re-cently, the proximal C–terminus of the protein sequence wasshown to be crucial for the targeting of D1–like receptors to thecell surface. A four amino acid spacing of hydrophobicresidues within the proximal C terminus of D1A/D1 and of theD1B/D5, the FxxxFxxxF motif, is highly conserved amongGPCR and has been suggested to interact with caveolin(Bermak et al., 2001). All three F residues within this motifare required for normal D1 targeting to the plasma mem-brane. In fact, the single substitution of any of the F to Aresulted in improper processing and trapping of the mutatedreceptor in the ER. The membrane-bound protein DRiP78,which is specifically localized in the ER and physicallyinteracts with the FxxxFxxxF motif, is able to regulate trans-port of the D1 receptor from the endoplasmic reticulum to thecell surface (Bermak et al., 2001). Whether similar mecha-nism is used for the D1B/D5 receptor is not known. However,it was found, that N–linked glycosylation, which takes placein the ER, is required for the correct membrane targeting ofD1B/D5 receptor, but not for the D1A/D1 subtype (Karpa et al.,1999). Interaction between the c subunit of COP-I coatomerand the C–terminus of D1 receptor has also been established,suggesting that this interactions may have a role in receptortransport (Bermak et al., 2002). Finally, the D1 receptorshave been shown to interact with neurofilament-M, theselatter being able to reduce the number of receptors in theplasma membrane and to decrease agonist-induced desensi-tization of the receptor (Kim et al., 2002).
A few laboratories have undertaken two-hybrid screens inyeast to identify dopamine receptor-interacting proteins thatmay regulate intracellular trafficking of the dopamine recep-tors subtypes. The D2 receptors have now been shown tointeract with several proteins, such as spinophilin (Smith etal., 1999), filamin-A (Lin et al., 2001), actin binding proteinABP-280 (Li et al., 2000) and protein 4.1 N (Binda et al.,2002, Kabbani et al., 2002). These proteins all arecytoskeleton-associated proteins and they are specificallyenriched in neurones. However, the role they have in target-ing or stabilizing dopamine receptors at specific subcellulardomains is not well understood yet. It is also possible thatthese cytoskeletal proteins may participate as scaffoldingproteins in organizing the dopamine receptor-signallingcomplex. Almost all the experiments performed to analyzeinteractions between these proteins and dopamine receptorswere carried out in cells lines and we do not know if theseinteractions also take place in neurons. The existence ofhighly specific structures such as neurites, postsynaptic den-sities or synaptic boutons, together with the absence of manyprotein partners present in neurons provide a level of com-plexity not easily mimicked in cells lines. Clearly, furtherstudies are needed to establish these cytoskeletal proteins asfunctionally relevant in neuronal cells.
494 S. Callier et al. / Biology of the Cell 95 (2003) 489–502

3. Functional properties of dopamine receptorsand modulation of signalling pathways
As already stated, dopamine receptors have been dividedinto two groups, based on ligand affinity and specificity andeffector coupling. Differences in ligand binding affinity,basal activity, efficacy and potency for G protein activation,or desensitization mechanisms may be expected to be derivedcharacters that drove conservation of the dopamine receptorsubtypes during vertebrate evolution. These derived charac-ters would be in fact the discriminating properties of receptorsubtypes found in modern vertebrate species. Here we showthat it is indeed the case. A detailed comparison of thefunctional characteristics of dopamine receptors among spe-cies is only available for the D1 receptor class, but someinferences can also be proposed for the D2 receptors.
3.1. D1 receptor signalling
Receptors of the D1 class, D1A/D1 and D1B/D5, activateadenylyl cyclase through the G protein Gas in most tissues,or Gaolf in the striatum and the olfactory bulb (Monsma et al.,1990, Tiberi et al., 1991; Herve et al., 1993), resulting inenhanced levels of cAMP in cells. As a consequence, cAMP-dependent protein kinase (PKA) is turned on, and inducesphosphorylation of cellular protein, such as DARPPP-32,ARPP-21, ARPP-16 and STEP. There is also evidence thatD1B/D5 receptor couples to Gaz and that D1A/D1 receptorcouples to Gao, thereby modulating the activity of Ca2+, K+,and Na+ channels (Sidhu, 1998; Sidhu and Niznik, 2000).Nevertheless, all D1 receptor subtypes preferentially activateGas, the coupling to others Ga proteins depending on thenature of the Ga proteins promiscuous to the receptors, onthe presence of associated proteins such as calcyon, and ofthe activation level of the receptor (Kimura et al., 1991;Kimura et al., 1995; Lezcano et al., 2000; Sidhu and Niznik,2000).
One of the clearest distinctions between the three D1A,D1B and D1C subtypes is made by the relative affinity of thereceptors for ligand and for the basal activity of the receptor.The binding for ligands and coupling properties of theD1B/D5 receptor differ from that of the D1A/D1 receptor by ahigher constitutive agonist-independent activity, an in-creased affinity and potency for dopamine and other agonists,and a lower affinity for inverse agonist, these properties beinghighly conserved among jawed vertebrates (Tiberi and Ca-ron, 1994; Cardinaud et al., 1997; Sugamori et al., 1998). Incontrast, the D1C receptor subtype exhibits essentially thesame dopamine affinity and low intrinsic activity as the D1A
subtype. These characteristics could be related to the struc-tural difference of cytoplasmic tails of each receptor since inthe three D1 receptor subtypes, it contains phosphorylationmotif, which are both subtype-specific and conserved in allthe species studied so far. Using chimeric constructions ofD1–like receptors, Jackon et al. (2000) have shown that thecytoplasmic tail of human dopamine D1–like receptors regu-late the binding affinity of dopamine and interacts with the
other intracellular loops of the receptor to modulate Gprotein-coupling and second messenger activation. In addi-tion, studies conducted in Xenopus with chimeric D1A, D1B
and D1C receptors showed that the cytoplasmic tail alsoplayed a role in the desensitization mechanisms of theseD1–like receptors (Sugamori et al., 1998; see below).
3.2. D2 receptor signalling
No systematic analysis of the functional characteristics ofthe three D2–like receptor subtypes has been undertaken, incontrast to the D1–like receptors. However, a few generalfeatures can be highlighted for the members of this dopaminereceptor class. Beside synthetic compounds, some of thembeing fairly specific of either the D2, the D3 or the D4
receptor, the natural agonist dopamine has a significantlyhigher affinity (ten-to-thirty fold) for the D3 and the D4 thanfor the D2 receptor subtype (Sokoloff and Schwartz, 1995.
Although the inhibition of adenylyl cyclase activity is oneof the most conserved characteristics of the D2–like receptors,the D3 receptor are not able to promote inhibition of cAMPproduction in fibroblastic cell lines. It does so only inneuronal-like cells, revealing that specific –but unknown–intracellular components may be necessary for receptor ac-tivity (Cussac et al., 1999). All three D2–like receptors are alsoable to modulate the activity of voltage-gated ionic channels.The D2 receptor subtype inhibits several types of Ca2+ ionicchannels through Gao and activates K+ currents through Gaiin pituitary cells (Lledo et al., 1992). Similarly, the D3 and D4
receptor subtypes are able to inhibit L and N–types of Ca2+
currents in transfected pituitary and neuronal cell lines(Seabrook et al., 1994).
Among the other effects of the D2–like receptors that havebeen observed (mobilisation of intracellular calcium stores,PKC activation, release of arachidonic acid, Na+/H+ ex-change, inhibition of the Na+/K+ ATPase{ etc), no evidenceof a direct modulation of these effectors has been demon-strated (reviewed in Missale et al., 1998). However, a directactivation of PLC (Vallar et al., 1990) or the modulation ofsome of the MAP-kinase pathways (Lajiness et al., 1993) areprobably important but underestimated effects of the D2
receptors.In amniotes, the D2 receptor subtype exists in two isofoms
of 444 (D2L) and 415 (D2S) amino acids generated by thealternative splicing of the pre-mRNA (reviewed in Missale etal., 1998). These two proteins are identical except for thethird intracytoplasmic loop, where there is an insertion of29 amino acids in D2L, relative to D2S. The question of thephysiological counterpart of these two has long been de-bated, and a definitive answer has not been obtained yet. Wehave discussed above the possibility of differential localiza-tion of the D2 receptor isoforms in neurons. At first sight, anobvious idea was that the 29 amino acid insertion might elicitdifferential coupling of the receptor to G proteins. As amatter of fact, in some cell lines, D2L but not D2S, couple toGai2 to inhibit forskolin-stimulated adenylyl cyclase (Mont-mayeur et al., 1993; Liu et al., 1994; Guiramand et al., 1995).
495S. Callier et al. / Biology of the Cell 95 (2003) 489–502

This is not found in other cells where no discriminatingproperties of the two D2 receptor isoforms could be seen(Senogles, 1994; Gharemani et al., 1999). However, the abil-ity of the D2 receptor isoforms to block Ca++ channels seemsto vary between D2S and D2L (Liu et al. 1994). In addition,the D2L receptor activates K+ channels through PTX-sensitive Ga subunits, whereas D2S receptor effect coulddepend on Gas inactivation (Liu et al., 1996). In conclusion,some clues exist for a differential activity of the two splicingisoforms of the dopamine D2 receptor. However, it mayreflect a differential localization in microdomains of theplasma membrane, as proposed above. It is the formation of alocal network of interacting proteins, regulating both trans-duction phenomenon as well as the dynamic of the receptorsbetween the subcellular compartments, which ultimately de-termines the cellular response to dopamine binding.
3.3. Dopamine receptors oligomerization and functionalprotein-protein interactions
Among the molecular interactions that may be crucial fordopaminergic transmission, the possibility that dopaminereceptors can form homo or heteromers shed a new light onthe mechanisms of transduction and receptor regulation.Many single-transmembrane receptors including growth-factor receptors and cytokine receptors depend on dimeriza-tion for proper function. In addition, metabotropic glutamatereceptors and GABAB receptors obligatorily dimerize to betransported to the plasma membrane and to transduce extra-cellular signals through G proteins. Whether monoamine Gprotein-coupled receptors also undergo dimerization is still amatter of debate. In addition to signal transduction, receptoroligomerization may be a prerequisite to desensitization,endocytosis and recycling of receptors (Hébert et al., 1996).Agonist or antagonist exposure may change oligomer:mono-mer ratio suggesting that interconversion between monomersand oligomers may be important for biological activity(Hébert and Bouvier, 1998).
Dopamine receptors have also been shown to formsdimers and higher order oligomers. Evidence has been pro-vided for dopamine D2 dimerization in the human and ratbrain by showing discrepancy in binding parameters fordifferent ligands. Indeed binding measurements of labelledbenzamide and butyrophenone derivatives led to the hypoth-esis that [3H]–spiperone binds to the D2 receptor monomerswhereas [3H]–raclopride interacts with receptor dimers,since [3H]–raclopride labelled about half the number of D2
receptor sites labelled by [3H]–spiperone (Zawarynski et al.,1998). This finding was supported by the demonstration ofdimers and oligomers formation of D1 and D2 receptors intransfected Sf9 cells (Ng et al., 1994a; Ng et al., 1996).Indirect evidence of D2 receptor oligomerization has beenobtained by the inhibition of wild-type D2 receptor functionby truncated amino and carboxy-terminal D2 receptor por-tions (Lee et al., 2000). In addition, agonist-independent anddependent oligomerization was suggested by using a bio-physical method based on energy transfer from fluorescent
protein-tagged D2 receptors in close spacial proximity inliving cells (Wurch et al., 2001). However, no conclusiveproof has been provided yet to demonstrate that oligomeriza-tion is mandatory for dopamine receptor activity.
In addition to homodimerization, it has been suggestedthat dopamine receptors not only form homodimers but alsoheterodimers with other GPCRs or even ligand-gated ionchannels. Rocheville et al. (2000) have shown by a FRET-based assay that D2 receptor interacts with somatostatinSSTR5 receptors, resulting in enhanced functional activity.Evidence for heteromerization of other dopamine receptorsubtypes has been also provided. Oligomeric D2:D3 receptorprotein associations have been detected by co-immunoprecipitation in primate and rodent brain homoge-nates, as well as in rat GH3 cells stably expressing a D3
receptor (Nimchinsky et al., 1997). Scarselli et al. (2001)used a truncated receptor approach to show that the N–termi-nal part of a D2 receptor could functionally associate withC–terminal portion of a D3 receptor. In addition, evidencesexist for a direct interaction between D1 and D3 receptors inventral striatum and islands of Calleja (reviewed in Sokoloffet al., 2003).
The D1 receptors certainly make hetero-oligomers withadenosine A1 receptors (Gines et al. 2000). The A1 and D1A
receptors are highly co-localized and directly interact instable lines of mouse fibroblast Ltk– cells and in primarycultures of cortical neurons. This selective A1/D1A receptorheteromerization seems to depend upon the simultaneousactivation of the two receptors. Indeed, pre-treatment of thecells byA1 receptor agonists prevents D1A receptor activationand desensitization (Gines et al. 2000; Le Crom et al., 2002).Another evidence of adenosine-dopamine receptor antago-nism was obtained by Hillion et al, (2002), who showed thatadenosine A2A and D2 receptors are co-localized in mem-branes of SH-SY5Y human neuroblastoma cells stably trans-fected with human D2 receptors and in cultured striatal cells.Moreover, long-term exposure to dopamine or adenosinereceptor agonists resulted in aggregation, internalization anddesensitization of both A2A and D2 receptors (Hillion et al.,2002).
Dopamine receptors heterodimerization can also modu-late trafficking and activity of ligand-gated ion channelsthrough direct protein-protein interactions. The D1 receptorwas shown to interact with the N-methyl-D-aspartate(NMDA) receptor, mediating changes in the NMDA-dependent currents Lee et al., 2002). Two regions in the D1
receptor carboxyl tail directly and selectively couple thesubunits NR1a and NR2A of the NMDA receptor. One inter-action inhibits NMDA receptor-gated currents, and the otherattenuates NMDA receptor-mediated excitotoxicity througha PI-3 kinase-dependent pathway (Lee et al., 2002). Directinteraction was also shown between D5 receptor of dopamineand ligand-gated ion channel GABAA receptor (Liu et al.,2000). GABAA–ligand–gated channels complex selectivelywith D5 receptors through the direct binding of the D5
carboxy-terminal domain with the second intracellular loop
496 S. Callier et al. / Biology of the Cell 95 (2003) 489–502
of the c2 subunit (short) of the GABAA receptor. This physi-cal association makes possible reciprocal inhibitory interac-tions between these receptors.
These data highlight new mechanisms of signal transduc-tion regulation that increase the dynamic of receptor regula-tion and functioning at synapses in the central nervous sys-tem, independently of classically defined second-messengersystems. Thus, it is probable that, in a near future carefulexamination of the dopamine receptor homo et hetero-oligomerization will lead to a better understanding of thespecificity and diversity of dopaminergic signalling. Thelong-separated fields of receptor transduction and receptorlocalization are clearly merging now, to provide a moreunitary view of receptor function.
4. Regulation of dopamine receptor activity
As other G-protein-coupled receptors, dopamine recep-tors undergo a variety of regulatory changes, among which
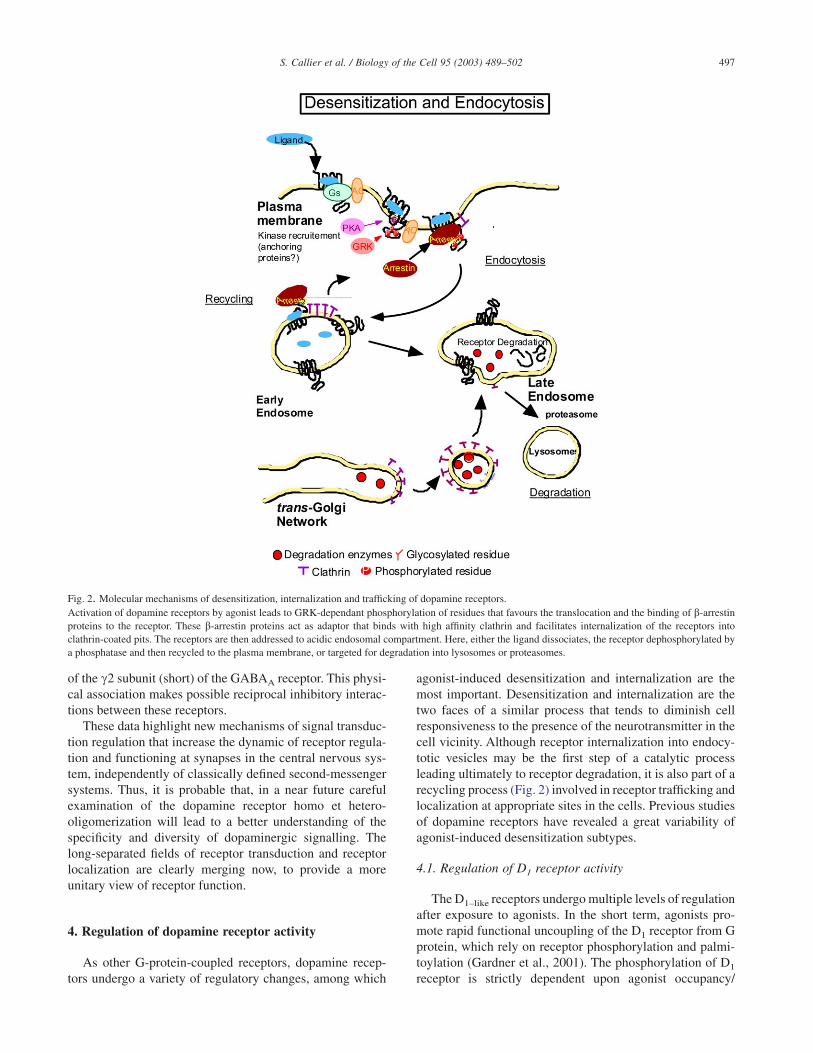
agonist-induced desensitization and internalization are themost important. Desensitization and internalization are thetwo faces of a similar process that tends to diminish cellresponsiveness to the presence of the neurotransmitter in thecell vicinity. Although receptor internalization into endocy-totic vesicles may be the first step of a catalytic processleading ultimately to receptor degradation, it is also part of arecycling process (Fig. 2) involved in receptor trafficking andlocalization at appropriate sites in the cells. Previous studiesof dopamine receptors have revealed a great variability ofagonist-induced desensitization subtypes.
4.1. Regulation of D1 receptor activity
The D1–like receptors undergo multiple levels of regulationafter exposure to agonists. In the short term, agonists pro-mote rapid functional uncoupling of the D1 receptor from Gprotein, which rely on receptor phosphorylation and palmi-toylation (Gardner et al., 2001). The phosphorylation of D1
receptor is strictly dependent upon agonist occupancy/
Fig. 2. Molecular mechanisms of desensitization, internalization and trafficking of dopamine receptors.Activation of dopamine receptors by agonist leads to GRK-dependant phosphorylation of residues that favours the translocation and the binding of b-arrestinproteins to the receptor. These b-arrestin proteins act as adaptor that binds with high affinity clathrin and facilitates internalization of the receptors intoclathrin-coated pits. The receptors are then addressed to acidic endosomal compartment. Here, either the ligand dissociates, the receptor dephosphorylated bya phosphatase and then recycled to the plasma membrane, or targeted for degradation into lysosomes or proteasomes.
497S. Callier et al. / Biology of the Cell 95 (2003) 489–502

activation of the receptor (Gardner et al., 2001). The secondstep consists in agonist-induced redistribution of surfacereceptor from a diffuse pattern of distribution to aggregates.Then, internalization of the ligand-receptor complex takesplace within small vesicles in the cytoplasm (Trogadis et al.,1995). The molecular mechanism of G protein coupled –receptor internalization has not been fully elucidated, butseveral lines of evidence indicate that protein-kinase A, Gprotein-coupled receptors kinases (GRKs), arrestin, as wellas clathrin and dynamin are involved (Fig.2). As mentionedabove, desensitization and internalization of D1–like receptorsmay represent two distinct –although interconnected– bio-chemical processes. For example, in SF9 cells, pretreatmentwith concavalin A or sucrose completely blocked agonist-induced internalization but not desensitization (Ng et al.,1995).
The respective roles of the different protein kinases,GRKs or second messenger-regulated kinases in the agonist-induced desensitization of D1 receptor is controversial.While some authors have underlined the central implicationof GRKs (see Ng et al., 1994a; Tiberi et al., 1996), othershave clearly shown that PKA may also play a role (Zhou etal., 1991). One of the phosphorylation sites located in theC–terminal end of the third cytoplasmic loop of the D1–like
receptors is conserved in the sequences of all the subtypes ofevery species known to date. This site is phosphorylated byPKA in agonist-induced desensitization, and mutation of thissite attenuated the rate of agonist-induced desensitization ofD1 receptors (Jiang and Sibley, 1999; Lamey et al., 2002).Interestingly, each of the D1 receptor subtypes bears alsoputative phosphorylation sites that are both subtype-specificand conserved in all the species analyzed so far. Whether theyare the substrates of different kinases has not yet been inves-tigated. It is possible that they underlie the differences ofinternalization rates displayed by the D1 receptor subtypes.Desensitization of the D1A receptor is occurring very fast,50% of receptor activity being lost during the first five min-utes. As far as the internalization process is concerned, D1A
dopamine receptors seem to preferentially use a b-arrestinand dynamin-dependent mechanism (Vickery and von Zas-trow, 1999, Iwata et al., 1999; Ito et al., 1999). However, howthis process contributes to receptor recycling or degradationis not known yet.
The human D1B/D5 receptor undergoes a similar processof desensitization but with lower amplitude than the D1A/D1
receptor (Jarvie et al., 1993; Iwasiow et al., 1999). Again, thischaracteristic is also found for the D1B receptors of the othervertebrate species (Tiberi and Caron, 1994; Cardinaud et al.,1997; reviewed in Kapsimali et al., 2003). We have proposedthat the differential kinetics of desensitization between D1A
and D1B receptors may be due to constitutive activity of theD1B receptor, this latter favoring a “constitutive” desensitiza-tion of the receptor. The desensitization of the D1B/D5 recep-tor is functionally coupled to endocytosis, as for the D1A/D1
receptor (unpublished observations), but details of the pro-cess need to be investigated further.
In sharp contrast with the two other subtypes of dopamineD1 receptors, D1C receptors from fish and amphibians areneither able to desensitize, nor to be internalized. Thus,although it still needs to be fully proven, it is very probablethat the lack of desensitization is an important discriminatingproperty of the D1C receptors. To conclude, desensitizationresponse to agonist exposure has been clearly diversified inthe different D1 receptor subtypes after gene duplications inearly vertebrates, and this property has probably contributedto the conservation of these subtypes later in evolution.
4.2. Regulation of receptor activity in the D2 class
The regulation of the D2–like receptors is rather differentfrom that of the D1–like receptors and remains highly contro-versial. For the D2 receptor subtype, some authors havedescribed desensitization of D2 receptors as a process closelycorrelated with agonist occupancy (Barton et al., 1991), andsomehow related to receptor sequestration (Itokawa et al.,1996). Others have reported that D2 receptor desensitizationoccurred, but slowly and only after prolonged agonist expo-sure, with some differences between the two D2L and D2S
receptor isoforms (Zhang et al., 1994). Finally and verysurprisingly, an up regulation of D2 receptors promoted byagonist stimulation on cultured cells has also been shown(Filtz et al., 1993; Starr et al., 1995; Ng et al., 1997). Reasonsfor such discrepancies are not clear at all, except to say thatthe regulation of the D2 receptor subtype may have somepeculiarity, perhaps the requirement of specific cell compo-nents, which is not easily recapitulated in heterologous trans-fected cells.
Among eccentricities of the D2 receptor subtype, a highintrisic activity has been described (Gardner et al., 1996;Prou et al, 2001) that may promote a constitutive endocytosisand recycling of the receptor at the cell surface (Vickery andvon Zastrow, 1999; Prou et al., 2001). Nevertheless, earlysteps of D2 receptor internalization seem to involve a rathercommon dynamin-dependent mechanism of endocytosis inearly endosomes (Vickery and von Zastrow, 1999, Iwata etal., 1999). Interstingly, in COS-7 and HEK-293 cells the D2
receptor sequestration induced by agonist treatment is ob-served only after coexpression with GRK2 or GRK5 (Ito etal., 1999; Iwata et al., 1999), but not in Sf9 cells (Ng et al.,1994b).
Very few studies have carefully compared the desensitiza-tion rate and mechanisms of the D2–like receptor subtypes.However, it has recently been shown that D2 and D3 receptorsexhibit some differences in both desensitization and traffick-ing properties (Kim et al., 2001). Contrasting with the D2
receptor, the D3 subtype is not phosphorylated upon agonistexposure, in the absence of GRK. Overexpression of GRKcaused only a subtle increase in the level of phosphorylationand the D3 receptor seems to have a reduced ability totranslocate b-arrestin to the plasma membrane, as comparedto the D2 receptor. Thus, as for the D1–like receptors, the D2
receptor subtypes are likely to be distinguishable by their
498 S. Callier et al. / Biology of the Cell 95 (2003) 489–502
properties of regulation in cells, the mechanisms of which arealso tightly linked to the intracellular trafficking and subcel-lular localization of the receptors.
5. Conclusions
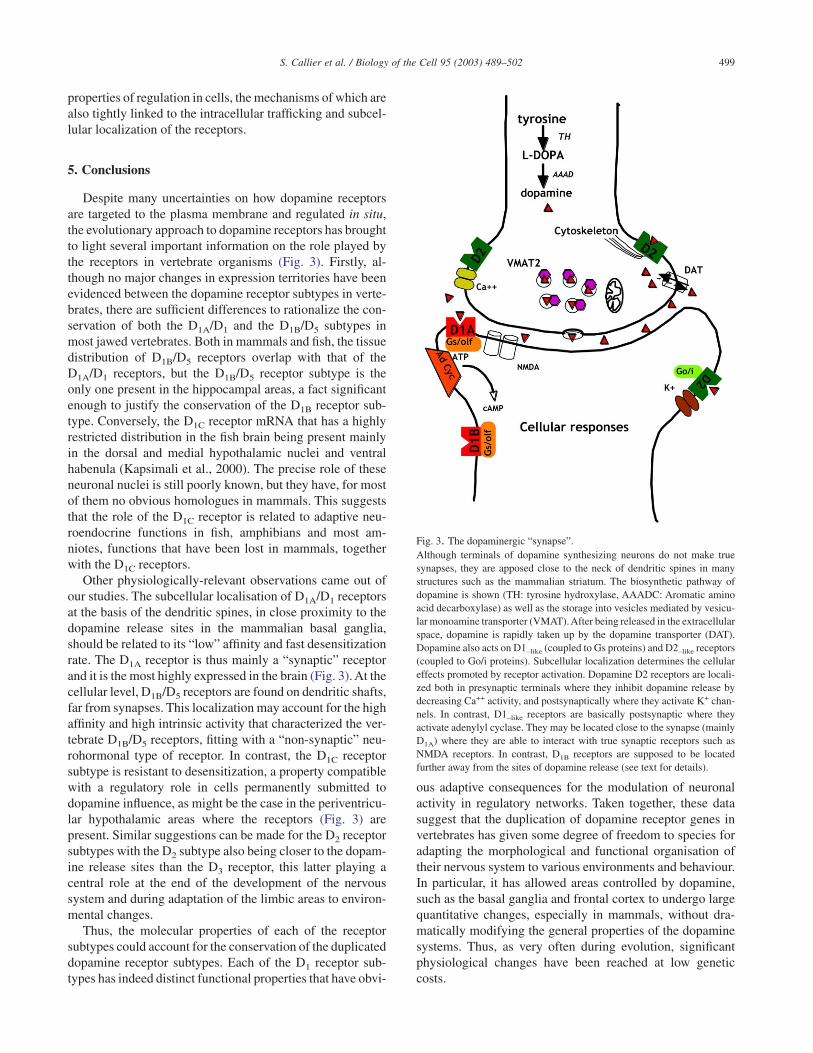
Despite many uncertainties on how dopamine receptorsare targeted to the plasma membrane and regulated in situ,the evolutionary approach to dopamine receptors has broughtto light several important information on the role played bythe receptors in vertebrate organisms (Fig. 3). Firstly, al-though no major changes in expression territories have beenevidenced between the dopamine receptor subtypes in verte-brates, there are sufficient differences to rationalize the con-servation of both the D1A/D1 and the D1B/D5 subtypes inmost jawed vertebrates. Both in mammals and fish, the tissuedistribution of D1B/D5 receptors overlap with that of theD1A/D1 receptors, but the D1B/D5 receptor subtype is theonly one present in the hippocampal areas, a fact significantenough to justify the conservation of the D1B receptor sub-type. Conversely, the D1C receptor mRNA that has a highlyrestricted distribution in the fish brain being present mainlyin the dorsal and medial hypothalamic nuclei and ventralhabenula (Kapsimali et al., 2000). The precise role of theseneuronal nuclei is still poorly known, but they have, for mostof them no obvious homologues in mammals. This suggeststhat the role of the D1C receptor is related to adaptive neu-roendocrine functions in fish, amphibians and most am-niotes, functions that have been lost in mammals, togetherwith the D1C receptors.
Other physiologically-relevant observations came out ofour studies. The subcellular localisation of D1A/D1 receptorsat the basis of the dendritic spines, in close proximity to thedopamine release sites in the mammalian basal ganglia,should be related to its “low” affinity and fast desensitizationrate. The D1A receptor is thus mainly a “synaptic” receptorand it is the most highly expressed in the brain (Fig. 3). At thecellular level, D1B/D5 receptors are found on dendritic shafts,far from synapses. This localization may account for the highaffinity and high intrinsic activity that characterized the ver-tebrate D1B/D5 receptors, fitting with a “non-synaptic” neu-rohormonal type of receptor. In contrast, the D1C receptorsubtype is resistant to desensitization, a property compatiblewith a regulatory role in cells permanently submitted todopamine influence, as might be the case in the periventricu-lar hypothalamic areas where the receptors (Fig. 3) arepresent. Similar suggestions can be made for the D2 receptorsubtypes with the D2 subtype also being closer to the dopam-ine release sites than the D3 receptor, this latter playing acentral role at the end of the development of the nervoussystem and during adaptation of the limbic areas to environ-mental changes.
Thus, the molecular properties of each of the receptorsubtypes could account for the conservation of the duplicateddopamine receptor subtypes. Each of the D1 receptor sub-types has indeed distinct functional properties that have obvi-
ous adaptive consequences for the modulation of neuronalactivity in regulatory networks. Taken together, these datasuggest that the duplication of dopamine receptor genes invertebrates has given some degree of freedom to species foradapting the morphological and functional organisation oftheir nervous system to various environments and behaviour.In particular, it has allowed areas controlled by dopamine,such as the basal ganglia and frontal cortex to undergo largequantitative changes, especially in mammals, without dra-matically modifying the general properties of the dopaminesystems. Thus, as very often during evolution, significantphysiological changes have been reached at low geneticcosts.
Fig. 3. The dopaminergic “synapse”.Although terminals of dopamine synthesizing neurons do not make truesynapses, they are apposed close to the neck of dendritic spines in manystructures such as the mammalian striatum. The biosynthetic pathway ofdopamine is shown (TH: tyrosine hydroxylase, AAADC: Aromatic aminoacid decarboxylase) as well as the storage into vesicles mediated by vesicu-lar monoamine transporter (VMAT).After being released in the extracellularspace, dopamine is rapidly taken up by the dopamine transporter (DAT).Dopamine also acts on D1–like (coupled to Gs proteins) and D2–like receptors(coupled to Go/i proteins). Subcellular localization determines the cellulareffects promoted by receptor activation. Dopamine D2 receptors are locali-zed both in presynaptic terminals where they inhibit dopamine release bydecreasing Ca++ activity, and postsynaptically where they activate K+ chan-nels. In contrast, D1–like receptors are basically postsynaptic where theyactivate adenylyl cyclase. They may be located close to the synapse (mainlyD1A) where they are able to interact with true synaptic receptors such asNMDA receptors. In contrast, D1B receptors are supposed to be locatedfurther away from the sites of dopamine release (see text for details).
499S. Callier et al. / Biology of the Cell 95 (2003) 489–502

Acknowledgements
Work in our laboratory is supported by CNRS, UniversitéParis XI and Institut Universitaire de France.
References
Aparicio, S., 2000. Vertebrate evolution: recent perspectives from fish.Trends Genet 16, 54–56.
Barton, A.C., Black, L.E., Sibley, D.R., 1991. Agonist-induced desensitiza-tion of D2 dopamine receptors in humanY–79 retinoblastoma cells. Mol.Pharmacol. 39, 650–658.
Bergson, C., Mrzljak, L., Smiley, J.F., Pappy, M., Levenson, R., Goldman-Rakic, P.S., 1995. Regional, cellular, and subcellular variations in thedistribution of D1 and D5 dopamine receptors in primate brain. J Neu-rosci. 15, 7821–7836.
Bermak, J.C., Li, M., Bullock, C., Zhou, Q.Y., 2001. Regulation of transportof the dopamine D1 receptor by a new membrane-associated ER protein.Nat Cell Biol 3, 492–498.
Bermak, J.C., Li, M., Bullock, C., Weingarten, P., Zhou, Q.Y., 2002. Inter-action of gamma-COP with a transport motif in the D1 receptor C–ter-minus. Eur J Cell Biol 81, 77–85.
Binda, A.V., Kabbani, N., Lin, R., Levenson, R., 2002. D2 and D3 dopaminereceptor cell surface localization mediated by interaction with protein4.1N. Mol Pharmacol 62, 507–513.
Cardinaud, B., Sugamori, K.S., Coudouel, S., Vincent, J.D., Niznik, H.B.,Vernier, P., 1997. Early emergence of three dopamine D1 receptorsubtypes in vertebrates. Molecular phylogenetic, pharmacological, andfunctional criteria defining D1A, D1B, and D1C receptors in Europeaneel Anguilla anguilla. J Biol Chem 272, 2778–2787.
Cussac, D.A.N.-T., Pasteau, V., Millan, M.J., 1999. Human dopamine D(3)receptors mediate mitogen-activated protein kinase activation via a phos-phatidylinositol 3–kinase and an atypical protein kinase C–dependentmechanism. Mol Pharmacol 56, 1025–1030.
Demchyshyn, L.L., Sugamori, K.S., Lee, F.J., Hamadanizadeh, S.A.,Niznik, H.B., 1995. The dopamine D1D receptor. Cloning and charac-terization of three pharmacologically distinct D1–like receptors fromGallus domesticus. J Biol Chem 270, 4005–4012.
Filtz, T.M., Artymyshyn, R.P., Guan, W., Molinoff, P.B., 1993. Paradoxicalregulation of dopamine receptors in transfected 293 cells. Mol Pharma-col 44, 371–379.
Gardner, B., Hall, D.A., Strange, P.G., 1996. Pharmacological analysis ofdopamine stimulation of (35S)–GTP gamma S binding via humanD2short and D2long dopamine receptors expressed in recombinant cells.Br J Pharmacol. 118 (6), 1544–1550.
Gardner, B., Liu, Z.F., Jiang, D., Sibley, D.R., 2001. The role ofphosphorylation/dephosphorylation in agonist-induced desensitizationof D1 dopamine receptor function: evidence for a novel pathway forreceptor dephosphorylation. Mol Pharmacol 59, 310–321.
Gerfen, C.R., 2000. Molecular effects of dopamine on striatal-projectionpathways. Trends Neurosci. 23 (Suppl), S64–S70.
Gines, S., Hillion, J., Torvinen, M., Le Crom, S., Casado, V., Canela, E.I.,Rondin, S., Lew, J.Y., Watson, S., Zoli, M., et al., 2000. Dopamine D1and adenosine A1 receptors form functionally interacting heteromericcomplexes. Proc Natl Acad Sci U S A 97, 8606–8611.
Guiramand, J., Montmayeur, J.P., Ceraline, J., Bhatia, M., Borrelli, E., 1995.Alternative splicing of the dopamine D2 receptor directs specificity ofcoupling to G–proteins. J. Biol. Chem. 270, 7354–7358.
Guivarc’h, D., Vernier, P., Vincent, J.D., 1995. Sex steroid hormones changethe differential distribution of the isoforms of the D2 dopamine receptormessenger RNA in the rat brain. Neuroscience 69, 159–166.
Hébert, T.E., Moffet, S., Morello, J.-P., Loisel, T.P., Bichet, D.G., Barret, C.,Bouvier, M., 1996. A peptide derived from a ß2–adrenergic receptortransmembrane domain inhibits both receptor dimerization and activa-tion. Journal of Biological Chemistry 271, 16384–16392.
Hébert, T.E., Bouvier, M., 1998. Structural and functional aspects of Gprotein-coupled receptor oligomerization. Biochem. Cell Biol. 76, 1–11.
Herve, D., Levi-Strauss, M., Marey-Semper, I., Verney, C., Tassin, J.P.,Glowinski, J., Girault, J.A., 1993. G(olf) and Gs in rat basal ganglia:possible involvement of G(olf) in the coupling of dopamine D1 receptorwith adenylyl cyclase. J Neurosci 13, 2237–2248.
Hillion, J., Canals, M., Torvinen, M., Casado, V., Scott, R., Terasmaa, A.,Hansson, A., Watson, S., Olah, M.E., Mallol, J., et al., 2002. Coaggrega-tion, cointernalization, and codesensitization of adenosine A2A recep-tors and dopamine D2 receptors. J Biol Chem 277, 18091–18097.
Huang, Q., Zhou, D., Chase, K., Gusella, J.F., Aronin, N., DiFiglia, M.,1992. Immunohistochemical localization of the D1 dopamine receptor inrat brain reveals its axonal transport, pre– and postsynaptic localization,and prevalence in the basal ganglia, limbic system, and thalamic reticularnucleus. Proc Natl Acad Sci U S A 89, 11988–11992.
Ito, K., Haga, T., Lameh, J., Sadee, W., 1999. Sequestration of dopamine D2receptors depends on coexpression of G-protein-coupled receptorkinases 2 or 5. Eur J Biochem 260, 112–119.
Itokawa, M., Toru, M., Ito, K., Tsuga, H., Kameyama, K., Haga, T., Ari-nami, T., Hamaguchi, H., 1996. Sequestration of the short and longisoforms of dopamine D2 receptors expressed in Chinese hamster ovarycells. Mol Pharmacol 49, 560–566.
Iwasiow, R.M., Nantel, M.F., Tiberi, M., 1999. Delineation of the structuralbasis for the activation properties of the dopamine D1 receptor subtypes.J Biol Chem 274, 31882–31890.
Iwata, K., Ito, K., Fukuzaki, A., Inaki, K., Haga, T., 1999. Dynamin and rab5regulate GRK2–dependent internalization of dopamine D2 receptors.Eur J Biochem 263, 596–602.
Jackson, A., Iwasiow, R.M., Tiberi, M., 2000. Distinct function of thecytoplasmic tail in human D1–like receptor ligand binding and coupling.FEBS Lett 470, 183–188.
Jarvie, K.R., Tiberi, M., Silvia, C., Gingrich, J.A., Caron, M.G., 1993.Molecular cloning, stable expression and desentization of the humandopamine D1B/D5 receptor. J. Recept.Res. 13, 573–590.
Jiang, D., Sibley, D.R., 1999. Regulation of D(1) dopamine receptors withmutations of protein kinase phosphorylation sites: attenuation of the rateof agonist-induced desensitization. Mol Pharmacol 56, 675–683.
Kabbani, N., Negyessy, L., Lin, R., Goldman-Rakic, P., Levenson, R., 2002.Interaction with neuronal calcium sensor NCS-1 mediates desensitiza-tion of the D2 dopamine receptor. J Neurosci 22, 8476–8486.
Kapsimali, M., Vidal, B., Gonzalez, A., Dufour, S., Vernier, P., 2000. Distri-bution of the mRNA encoding the four dopamine D(1) receptor subtypesin the brain of the european eel (Anguilla anguilla): comparativeapproach to the function of D(1) receptors in vertebrates. J Comp Neurol419, 320–343.
Kapsimali, M., Le Crom, S., Vernier, P., 2003. Origin and evolution ofdopamine receptors and transporters in vertebrates. In: Sidhu, M.L.A.,Vernier, P. (Eds.), Dopamine receptors and transporter. Marcel Dekker,New York, pp. 1–43.
Karpa, K.D., Lidow, M.S., Pickering, M.T., Levenson, R., Bergson, C.,1999. N–linked glycosylation is required for plasma membrane localiza-tion of D5, but not D1, dopamine receptors in transfected mammaliancells. Mol Pharmacol 56, 1071–1078.
Kebabian, J.W., Calne, D.B., 1979. Multiple receptors for dopamine. Nature277, 93–96.
Khan, Z.U., Gutierrez, A., Martin, R., Penafiel, A., Rivera, A., De LaCalle, A., 1998. Differential regional and cellular distribution of dopam-ine D2–like receptors: an immunocytochemical study of subtype-specific antibodies in rat and human brain. J Comp Neurol 402, 353–371.
Kim, K.M., Valenzano, K.J., Robinson, S.R., Yao, W.D., Barak, L.S.,Caron, M.G., 2001. Differential regulation of the dopamine D2 and D3receptors by G protein-coupled receptor kinases and beta-arrestins. JBiol Chem 276, 37409–37414.
Kim, O.J., Ariano, M.A., Lazzarini, R.A., Levine, M.S., Sibley, D.R., 2002.Neurofilament-M interacts with the D1 dopamine receptor to regulatecell surface expression and desensitization. J Neurosci 22, 5920–5930.
500 S. Callier et al. / Biology of the Cell 95 (2003) 489–502

Kimura, K., Sela, S., Bouvier, C., Grandy, D.K., Sidhu, A., 1995. Differen-tial coupling of D1 and D5 dopamine receptors to guanine nucleotidebinding proteins in transfected GH4C1 rat somatomammotrophic cells. JNeurochem 64, 2118–2124.
Kirschner, M., Gerhart, J., 1998. Evolvability. Proc Natl Acad Sci U S A 95,8420–8427.
Lajiness, M.E., Chio, C.L., Huff, R.M., 1993. D2 dopamine receptor stimu-lation of mitogenesis in transfected Chinese hamster ovary cells: rela-tionship to dopamine stimulation of tyrosine phosphorylations. J. Pharm.Exp. Ther. 267, 1573–1581.
Lamey, M., Thompson, M., Varghese, G., Chi, H., Sawzdargo, M.,George, S.R., O’Dowd, B.F., 2002. Distinct residues in the carboxyl tailmediate agonist-induced desensitization and internalization of thehuman dopamine D1 receptor. J Biol Chem 277, 9415–9421.
Le Crom, S., Prou, D., Vernier, P., 2002. Autocrine Activation of AdenosineA1 Receptors Blocks D1A but not D1B Dopamine Receptor Desensitiza-tion. J. Neurochem. 82, 1549–1552.
Lee, F.J., Xue, S., Pei, L., Vukusic, B., Chery, N., Wang, Y., Wang, Y.T.,Niznik, H.B., Yu, X.M., Liu, F., 2002. Dual regulation of NMDA recep-tor functions by direct protein-protein interactions with the dopamine D1receptor. Cell 111, 219–230.
Lee, S.P., Xie, Z., Varghese, G., Nguyen, T., O’Dowd, B.F., George, S.R.,2000. Oligomerization of dopamine and serotonin receptors. Neuropsy-chopharmacology 23, S32–S40.
Levey, A.I., Hersch, S.M., Rye, D.B., Sunahara, R.K., Niznik, H.B.,KItt, C.A., Price, D.L., Maggio, R., Brann, M.R., Ciliax, B.J., 1993.Localization of D1 and D2 dopamine receptors in brain with subtype-specific antibodies. Proc. Natl Acad. Sci. USA 90, 8861–8865.
Lezcano, N., Mrzljak, L., Eubanks, S., Levenson, R., Goldman-Rakic, P.,Bergson, C., 2000. Dual signaling regulated by calcyon, a D1 dopaminereceptor interacting protein. Science 287, 1660–1664.
Li, M., Bermak, J.C., Wang, Z.W., Zhou, Q.Y., 2000. Modulation of dopam-ine D(2) receptor signaling by actin-binding protein (ABP-280). MolPharmacol 57, 446–452.
Lin, R., Karpa, K., Kabbani, N., Goldman-Rakic, P., Levenson, R., 2001.Dopamine D2 and D3 receptors are linked to the actin cytoskeleton viainteraction with filamin A. Proc Natl Acad Sci U S A 98, 5258–5263.
Liu, F., Wan, Q., Pristupa, Z.B., Yu, X.M., Wang, Y.T., Niznik, H.B., 2000.Direct protein-protein coupling enables cross-talk between dopamineD5 and gamma-aminobutyric acid A receptors. Nature 403, 274–280.
Liu, L.X., Monsma Jr., F.J., Sibley, D.R., Chiodo, L.A., 1996. D2L, D2S,and D3 dopamine receptors stably transfected into NG108–15 cellscouple to a voltage-dependent potassium current via distinct G proteinmechanisms. Synapse 24, 156–164.
Liu, Y.F., Jakobs, K.H., Rasenick, M.M., Albert, P.R., 1994. G proteinspecificity in receptor-effector coupling. Analysis of the roles of G0 andGi2 in GH4C1 pituitary cells. J Biol Chem 269, 13880–13886.
Lledo, P.M., Homburger, V., Bockaert, J., Vincent, J.D., 1992. Differential Gprotein-mediated coupling of D2 dopamine receptors to K+ and Ca2+currents in rat anterior pituitary cells. Neuron 8, 455–463.
Meador-Woodruff, J.H., Damask, S.P., Wang, J., Haroutunian, V.,Davis, K.L., Watson, S.J., 1996. Dopamine receptor mRNA expressionin human striatum and neocortex. Neuropsychopharmacology 15,17–29.
Missale, C., Nash, S.R., Robinson, S.W., Jaber, M., Caron, M.G., 1998.Dopamine receptors: from structure to function. Physiol Rev 78, 189–225.
Monsma, F.J., Mahan, L.C., McVittie, L.D., Gerfen, C.R., Sibley, D.R.,1990. Molecular cloning and expression of a D1 dopamine receptorlinked to adenylyl cyclase activation. Proc. Natl Acad. Sci. USA 87,6723–6727.
Montmayeur, J.P., Guiramand, J., Borrelli, E., 1993. Preferential couplingbetween dopamine D2 receptors and G–proteins. Mol Endocrinol. 7,161–170.
Ng, G.Y., Mouillac, B., George, S.R., Caron, M., Dennis, M., Bouvier, M.,O’Dowd, B.F., 1994. Desensitization, phosphorylation and palmitoyla-tion of the human dopamine D1 receptor. Eur. J. Pharmacol. 267, 7–19.
Ng, G.Y., O’Dowd, B.F., Caron, M., Dennis, M., Brann, M.R., George, S.R.,1994. Phosphorylation and palmitoylation of the human D2L dopaminereceptor in Sf9 cells. J Neurochem 63, 1589–1595.
Ng, G.Y., Trogadis, J., Stevens, J., Bouvier, M., O’Dowd, B., George, S.R.,1995. Agonist-induced desensitization of dopamine D1 receptor-stimulated adenylyl cyclase activity is temporally and biochemicallyseparated from D1 receptor internalization. Proceedings of the NationalAcademy of Sciences of the United States of America 92, 10157–10161.
Ng, G.Y.K., O’Dowd, B.F., Lee, S.P., Chung, H.T., Brann, M.R., Seeman, P.,George, S.R., 1996. Dopamine D2 receptor dimers and receptor-blocking peptides. Biochem. Biophys. Res. Com. 227, 200–204.
Ng, G.Y.K., Varghese, G., Chung, H.T., Trogadis, J., Seeman, P.,O’Dowd, B.F., George, S.R., 1997. Resistance of the dopamine D2(L)receptor to desensitization accompanies the up-regulation of receptorson to the surface of Sf9 cells. Endocrinology 138, 4199–4206.
Nimchinsky, E.A., Hof, P.R., Janssen, W.G.M., Morrison, J.H.,Schmauss, C., 1997. Expression of dopamine D–3 receptor dimers andtetramers in brain and in transfected cells. J Biol Chem 272, 29229–29237.
Nothen, M.M., Cichon, S., Hemmer, S., Hebebrand, J., Remschmidt, H.,Lehmkuhl, G., Poustka, F., Schmidt, M., Catalano, M., Fim-mers, R., et al., 1994. Human dopamine D4 receptor gene: frequentoccurrence of a null allele and observation of homozygosity. Hum MolGenet. 3, 2207–2212.
Prou, D., Gu, W.J., Le Crom, S., Vincent, J.D., Salamero, J., Vernier, P.,2001. Intracellular retention of the two isoforms of the D(2) dopaminereceptor promotes endoplasmic reticulum disruption. J Cell Sci 114,3517–3527.
Rocheville, M., Lange, D.C., Kumar, U., Patel, S.C., Patel, R.C., Patel,Y.C.,2000. Receptors for dopamine and somatostatin: formation of hetero-oligomers with enhanced functional activity. Science 288, 154–157.
Scarselli, M., Novi, F., Schallmach, E., Lin, R., Baragli, A., Colzi, A.,Griffon, N., Corsini, G.U., Sokoloff, P., Levenson, R., et al., 2001.D2/D3 dopamine receptor heterodimers exhibit unique functional prop-erties. J Biol Chem 276, 30308–30314.
Schnell, S.A., You, S.Y., El Halawani, M.E., 1999. D1 and D2 dopaminereceptor messenger ribonucleic acid in brain and pituitary during thereproductive cycle of the turkey hen. Biol Reprod 60, 1378–1383.
Seabrook, G.R., Knowles, M., Brown, N., Myers, J., Sinclair, H., Patel, S.,Freedman, S.B., McAllister, G., 1994. Pharmacology of high-thresholdcalcium currents in GH4C1 pituitary cells and their regulation by activa-tion of human D2 and D4 dopamine receptors. Br J Pharmacol 112,728–734.
Seeman, P., Van Tol, H.H., 1994. Dopamine receptor pharmacology. TrendsPharmacol Sci 15, 264–270.
Senogles, S.E., 1994. The D2 dopamine receptor isoforms signal throughdistinct Gi alpha proteins to inhibit adenylyl cyclase. A study withsite-directed mutant Gi alpha proteins. J Biol Chem 269, 23120–23127.
Sibley, D.R., 1999. New insights into dopaminergic receptor function usingantisense and genetically altered animals. Annu Rev Pharmacol Toxicol39, 313–341.
Sidhu, A., 1998. Coupling of D1 and D5 dopamine receptors to multiple Gproteins: Implications for understanding the diversity in receptor-Gprotein coupling. Mol Neurobiol 16, 125–134.
Sidhu, A., Niznik, H.B., 2000. Coupling of dopamine receptor subtypes tomultiple and diverse G proteins. Int J Dev Neurosci 18, 669–677.
Smiley, J.F., Levey, A.I., Ciliax, B.J., Goldman-Rakic, P.S., 1994. D1dopamine receptor immunoreactivity in human and monkey cerebralcortex: predominant and extrasynaptic localization in dendritic spines.Proc. Natl Acad. Sci. USA 91, 5720–5724.
Smith, F.D., Oxford, G.S., Milgram, S.L., 1999. Association of the D2dopamine receptor third cytoplasmic loop with spinophilin, a proteinphosphatase-1-interacting protein. J Biol Chem 274, 19894–19900.
Sokoloff, P., Schwartz, J.C., 1995. Novel dopamine receptors half a decadelater [see comments]. Trends Pharmacol Sci 16, 270–275.
501S. Callier et al. / Biology of the Cell 95 (2003) 489–502

Sokoloff, P., Griffon, N., Guillin, O., Pilon, C., Diaz, J., Schwartz, J.C., P.,2003. The dopamine D3 receptor in the pathophysiology and treatmentsof neuropsychiatric disorders. In: Sidhu, M.L.A., Vernier, P. (Eds.),Dopamine receptors and transporter. Marcel Dekker, NewYork, pp. 211–253.
Starr, S., Kozell, L.B., Neve, K.A., 1995. Drug-induced up-regulation ofdopamine D2 receptor on cultured cells. J. Neurochem. 65, 569–577.
Strange, P.G., 1993. New insights into dopamine receptors in the centralnervous system. Neurochem Int 22, 223–236.
Sugamori, K.S., Demchyshyn, L.L., Chung, M., Niznik, H.B., 1994. D1A,D1B, and D1C dopamine receptors from Xenopus laevis. Proc Natl AcadSci USA 91, 10536–10540.
Sugamori, K.S., Scheideler, M.A., Vernier, P., Niznik, H.B., 1998. Dopam-ine D1B receptor chimeras reveal modulation of partial agonist activityby carboxyl-terminal tail sequences. J Neurochem 71, 2593–2599.
Tiberi, M., Jarvie, K.R., Silvia, C., Falardeau, P., Gingrich, J.A., Godi-not, N., Bertrand, L., Yang-Feng, T.L., Fremeau Jr, R.T., Caron, M.G.,1991. Cloning, molecular characterization, and chromosomal assign-ment of a gene encoding a second D1 dopamine receptor subtype:differential expression pattern in rat brain compared with the D1Areceptor. Proc Natl Acad Sci U S A 88, 7491–7495.
Tiberi, M., Caron, M.G., 1994. High agonist-independent activity is adistinguishing feature of the dopamine D1B receptor subtype. J BiolChem 269, 27925–27931.
Tiberi, M., Nash, S.R., Bertrand, L., Lefkovitz, R.J., Caron, M.G., 1996.Differential regulation of dopamine D1A receptor responsiveness byvarious G protein-couped receptor kinases. Journal of Biological Chem-istry 271, 3771–3778.
Trogadis, J.E., Ng, G.Y., O’Dowd, B.F., George, S.R., Stevens, J.K., 1995.Dopamine D1 receptor distribution in Sf9 cells imaged by confocalmicroscopy: a quantitative evaluation. J Histochem Cytochem 43, 497–506.
Vallar, L., Muca, C., Magni, M., Albert, P., Bunzow, J., Meldolesi, J.,Civelli, O., 1990. Differential coupling of dopaminergic D2 receptorsexpressed in different cell types. Stimulation of phosphatidylinositol4,5– bisphosphate hydrolysis in LtK– fibroblasts, hyperpolarization, andcytosolic-free Ca2+ concentration decrease in GH4C1 cells. J Biol Chem265, 10320–10326.
Vernier, P., Cardinaud, B., Valdenaire, O., Philippe, H., Vincent, J.D., 1995.An evolutionary view of drug-receptor interaction: the bioamine recep-tor family. Trends Pharmacol Sci 16, 375–381.
Vickery, R.G., von Zastrow, M., 1999. Distinct dynamin-dependent and-independent mechanisms target structurally homologous dopaminereceptors to different endocytic membranes. J Cell Biol 144, 31–43.
Wurch, T., Matsumoto, A., Pauwels, P.J., 2001. Agonist-independent and-dependent oligomerization of dopamine D(2) receptors by fusion tofluorescent proteins. FEBS Lett 507, 109–113.
Yung, K.K., Bolam, J.P., Smith, A.D., Hersch, S.M., Ciliax, B.J., Levey, A.I.,1995. Immunocytochemical localization of D1 and D2 dopamine recep-tors in the basal ganglia of the rat: light and electron microscopy.Neuroscience 65, 709–730.
Zawarynski, P., Tallerico, T., Seeman, P., Lee, S.P., O’Dowd, B.F.,George, S.R., 1998. Dopamine D2 receptor dimers in human and ratbrain. FEBS Lett 441, 383–386.
Zhang, L.J., Lachowicz, J.E., Sibley, D.R., 1994. The D2S and D2L dopam-ine receptor isoforms are differentially regulated in Chinese hamsterovary cells. Molecular Pharmacology 45, 878–889.
Zhou, X., Sidhu, A., Fishman, P.H., 1991. Desensitization of the humandopamine D1 receptor: Evidence for involvement of both cyclic AMP-dependent and receptor-specific kinases. Mol. Cell. Neurosci. 2, 264–272.
502 S. Callier et al. / Biology of the Cell 95 (2003) 489–502