Embed Size (px)
Citation preview

Original Paper
Expression and cellular localization of the urokinase-typeplasminogen activator and its receptor in humanhepatocellular carcinoma
L. Dubuisson1*, A. Monvoisin1, B. S. Nielsen2, B. Le Bail1, P. Bioulac-Sage1 and J. Rosenbaum1
1 Laboratoire de Pathologie et Groupe de Recherches pour l'Etude du Foie, INSERM E9917, Universite Victor Segalen Bordeaux 2, Bordeaux, France2 Finsen Laboratory, Rigshospitalet, Strandboulevarden 49, 2100 Copenhagen, Denmark
*Correspondence to:Dr Liliane Dubuisson, Groupe deRecherches pour l'Etude du Foie,INSERM E9917, Universite VictorSegalen Bordeaux 2,146 Rue LeÂo-Saignat, 33076Bordeaux Cedex, France.E-mail: [email protected]
Received: 8 February 1999
Revised: 9 June 1999
Accepted: 6 September 1999
Abstract
The urokinase-type plasminogen activator (uPA) and its receptor (uPAR) play an important role
in tumour invasion. Previous studies have shown by RT-PCR that uPA and uPAR mRNAs are
expressed in human hepatocellular carcinoma (HCC). Here, in situ hybridization, immuno-
histochemistry, and double immuno¯uorescence were used to identify the cells expressing uPA and
uPAR in 26 HCCs. The results indicate that uPA and uPAR were expressed in every case, almost
exclusively in stromal cells, mostly myo®broblasts and macrophages, except for rare tumoural
hepatocytes expressing cytokeratin 7. These results show the important role of stromal cells of
HCC in the pericellular proteolysis which facilitates cancer cell invasion. Copyright # 2000 John
Wiley & Sons, Ltd.
Keywords: urokinase-type plasminogen activator; urokinase-type plasminogen activator recep-
tor; hepatocellular carcinoma; in situ hybridization; immunohistochemistry; double immuno¯uor-
escence
Introduction
Human hepatocellular carcinoma (HCC) is the mostfrequent primary liver cancer. HCC is associated witha high rate of intrahepatic invasion, which requires thedegradation of the extracellular matrix by proteolyticenzymes including serine proteinases and matrixmetalloproteinases (MMPs) [1]. Plasmin is a serineproteinase derived from plasminogen, which is able todegrade most extracellular matrix proteins and also toactivate MMPs and growth factors [2,3]. Plasminogenactivation on the cell surface involves at least uro-kinase-type plasminogen activator (uPA) and its cellsurface receptor (uPAR) [4]. uPA is generated assingle-chain inactive pro-uPA, which is converted intoa two-chain active form. Following binding to uPAR,uPA activates plasminogen. Plasmin itself activatespro-uPA, leading to an ampli®cation of plasmingeneration on the cell surface [4]. Several studies havereported overexpression of uPA and/or uPAR inmalignant tumours, including cancer of the breast,colon, lung, and skin [5]. In breast and colon cancers, ahigh expression level of uPA or uPAR is associatedwith a poor prognosis [6]. Immunohistochemicalstudies indicate that stromal cells are often the mainproducers of both uPA and uPAR [7].
We [8] and others [9] have found that uPA anduPAR mRNAs are up-regulated in human HCC. Itwas reported that tumoural hepatocytes were the mainproducers of uPAR mRNA in HCC [10]. However, inanother study, uPAR immunoreactivity was found inthe stromal tumour component [11]. Here we have
used both in situ hybridization and immunohistochem-istry to study the cellular localization of uPA anduPAR in human HCCs.
Materials and methods
Material
HCC samples (n=26) came from patients undergoingpartial hepatectomy (n=11) or liver transplantation(n=15). Histologically normal liver at a distance froma colon carcinoma metastasis (n=2) or from focalnodular hyperplasia (n=2) was also used. Fresh tissuesamples from the tumour and the surrounding tissuewere frozen and stored at x80uC. Five-micrometre-thick serial frozen sections of the tumour were air-dried on Super Frost*/Plus slides (Menzel GlaÈser,Germany) and ®xed in acetone for immunohistochem-istry, or in 4% paraformaldehyde in phosphate-buffered saline for in situ hybridization.
Liver histology
HCCs were graded from 1 (well differentiated) to 4(poorly differentiated) as previously described [12].Their architecture was designated according to theWHO classi®cation [13].
cDNAs and antibodies
We used two non-overlapping uPA cDNA subclones,pHUPA10 and pHUPA13, and two non-overlappinguPAR cDNAs, pHUR-06 and pHUR-04 [14].
Journal of PathologyJ Pathol 2000; 190: 190±195.
Copyright # 2000 John Wiley & Sons, Ltd.ccc 0022-3417/2000/020190±06$17.50

Two monoclonal antibodies that recognize differentepitopes (R2, R4) and a preparation of rabbit poly-clonal antibodies against uPAR (puPAR) have beendescribed [15]. An af®nity-puri®ed IgG fraction ofrabbit polyclonal antibodies against uPA (puPA) was agift from Marianne Nielsson, Dako A/S, Glostrup,Denmark. The CD68 (clone PG-M1), CD31 (cloneJC/7OA), and cytokeratin 7 (clone OV-TL 12/30)antibodies were from Dako; the anti-alpha-smoothmuscle actin (a-SMA clone 1A4) was from Sigma(St Louis, MO, USA); and the anti-human cytokeratin19 was from Amersham Life Sciences (UK).
Immunoperoxidase histochemistry
Sections were blocked with 30% goat serum and thenincubated for 2 h with the antibodies to uPAR or touPA in Tris-buffered saline, pH 7.4+1% bovine serumalbumin, at the following dilutions: 1/50 for R2 andR4; 1/1000 for puPAR; and 1/2500 for puPA. Theepitopes were detected with an Envision+systemHRP detection kit (Dako) and revealed with diamino-benzidine. As a negative control, we used a rabbitnon-immune immunoglobulin fraction (Dako), or aclari®ed mouse myeloma ascites (Cappel ResearchProducts, Durham, USA), at the same concentrationas the respective antibodies.
Double immuno¯uorescence
Samples from ten patients were double-stained forpuPA or puPAR, together with monoclonal antibodiesagainst either a-SMA (1/400), CD68 (1/100), cytoker-atin 7 (1/100), cytokeratin 19 (1/20) or CD31 (1/100).After saturation of non-speci®c sites, sections wereincubated with puPA or puPAR mixed with one of themonoclonal antibodies. Detection was achieved with amix of AlexaTM 594-goat anti-rabbit IgG conjugateand Oregon greenTM 488-goat anti-mouse IgG con-jugate (1/100) (Molecular Probes, OR, USA) respec-tively. Sections were mounted in antifade mountingmedium (Dako) and examined with a MolecularDynamics (Sunnyvale, CA, USA) confocal scanninglaser microscope equipped with a krypton/argon laserand connected to a Silicon Graphics 3D workstation.
In situ hybridization
In situ hybridization with [a33P]UTP-labelled sense andanti-sense riboprobes was performed as previouslydescribed [16] with minor modi®cations concerningthe hybridization mixture (5 mM EDTA and 10 mM
sodium phosphate, pH 8.0).
Results
Liver specimens and clinical data
The HCCs came from 21 males and ®ve females of age27±73 years. Sixteen patients had cirrhosis, due to virusB, C, alcohol or a combination of these; the ten otherpatients had ®brotic (n=7) or only mildly abnormal
(n=3) livers. The tumours ranged from 0.7 to 12.5 cmin diameter and from grade 1 to 4, grades 2±3 being themost frequent. One carcinoma was ®brolamellar, whilemost of the others were of trabecular architecture.
In situ hybridization detection of uPA and uPARtranscripts
Low-level uPA transcripts were detected in every HCCwith the pHuPA10 probe. Similar results wereobtained for the ten HCCs tested with the pHuPA13probe. The signal appeared as spotty labelling overspindle-shaped cells with elongated nuclei and overgroups of smaller roundish cells with round nuclei(Figure 1a). These cells were mainly located within®brous bands at the periphery of the tumour, insmaller ®brous tracts dissecting the tumour, and some-times in the sinusoids between hepatocytes.
Similarly, for uPAR, the signal obtained with thepHURO6 probe was mainly found on stromal cellswithin ®brous bands around the tumour or in the small®brous tracts within the tumour (Figure 1d). The samesignal was also obtained with the pHURO4 probe usedin 13 patients.
In control liver, a weak signal over spindle cells wasfound only around the portal tracts for uPA anduPAR. In one liver sample at a distance from a coloncancer metastasis, we also found faint expression ofuPAR on roundish cells inside the sinusoids (notshown). Sense probes gave no signal (Figure 1c).
Immunoperoxidase detection of uPA and uPARproteins
uPA protein was detected in the same areas as itsmRNA, namely in the ®brous tissue surrounding thetumour (Figures 1b and 1h) and in thin ®brotic bandsinside the tumour (Figure 2a). In all the specimens, weobserved a strong signal in spindle cells and smallerroundish cells inside the tumour or at its periphery.The labelling was essentially cytoplasmic in thespindle-shaped cells and membranous or perinuclearin the roundish cells. Some immunoreactivity was alsofound in the extracellular matrix which surrounded thestromal cells. Immunostaining of uPAR followed thesame pattern as uPA in stromal cells at the peripheryof the tumour (Figures 1g and 2d) or in ®brous tractsinside the tumour (Figure 1e). Similar results wereobtained with the two monoclonal antibodies and thepolyclonal antibody. uPA and uPAR were often co-localized, as can be seen on adjacent sections (Figures1g and 1h).
In control liver, the immunostaining of uPA anduPAR was very low (not shown). No labelling was seenwith control IgGs (Figure 1f).
Identi®cation of cells expressing uPA and uPARby confocal scanning laser microscopy
We used a mix of uPA or uPAR antibodies withmarkers of myo®broblasts and smooth muscle cells
uPA and uPAR in human hepatocellular carcinoma 191
Copyright # 2000 John Wiley & Sons, Ltd. J Pathol 2000; 190: 190±195.
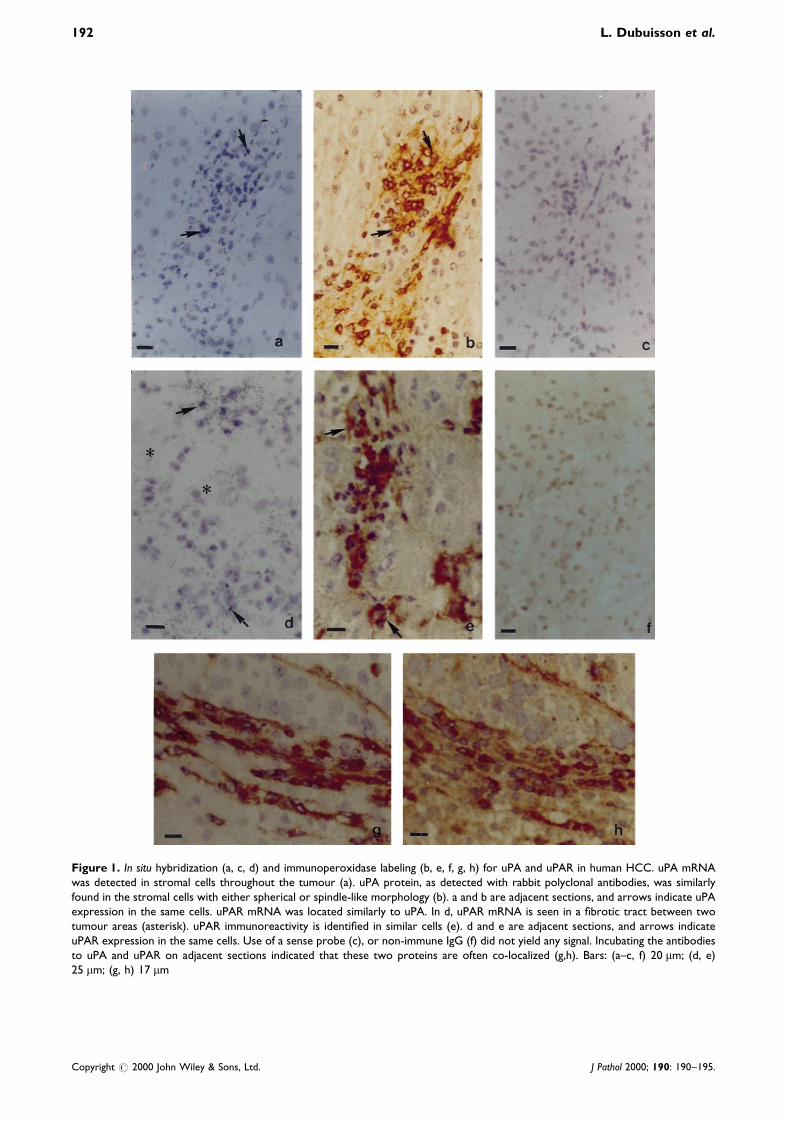
Figure 1. In situ hybridization (a, c, d) and immunoperoxidase labeling (b, e, f, g, h) for uPA and uPAR in human HCC. uPA mRNAwas detected in stromal cells throughout the tumour (a). uPA protein, as detected with rabbit polyclonal antibodies, was similarlyfound in the stromal cells with either spherical or spindle-like morphology (b). a and b are adjacent sections, and arrows indicate uPAexpression in the same cells. uPAR mRNA was located similarly to uPA. In d, uPAR mRNA is seen in a ®brotic tract between twotumour areas (asterisk). uPAR immunoreactivity is identi®ed in similar cells (e). d and e are adjacent sections, and arrows indicateuPAR expression in the same cells. Use of a sense probe (c), or non-immune IgG (f) did not yield any signal. Incubating the antibodiesto uPA and uPAR on adjacent sections indicated that these two proteins are often co-localized (g,h). Bars: (a±c, f) 20 mm; (d, e)25 mm; (g, h) 17 mm
192 L. Dubuisson et al.
Copyright # 2000 John Wiley & Sons, Ltd. J Pathol 2000; 190: 190±195.
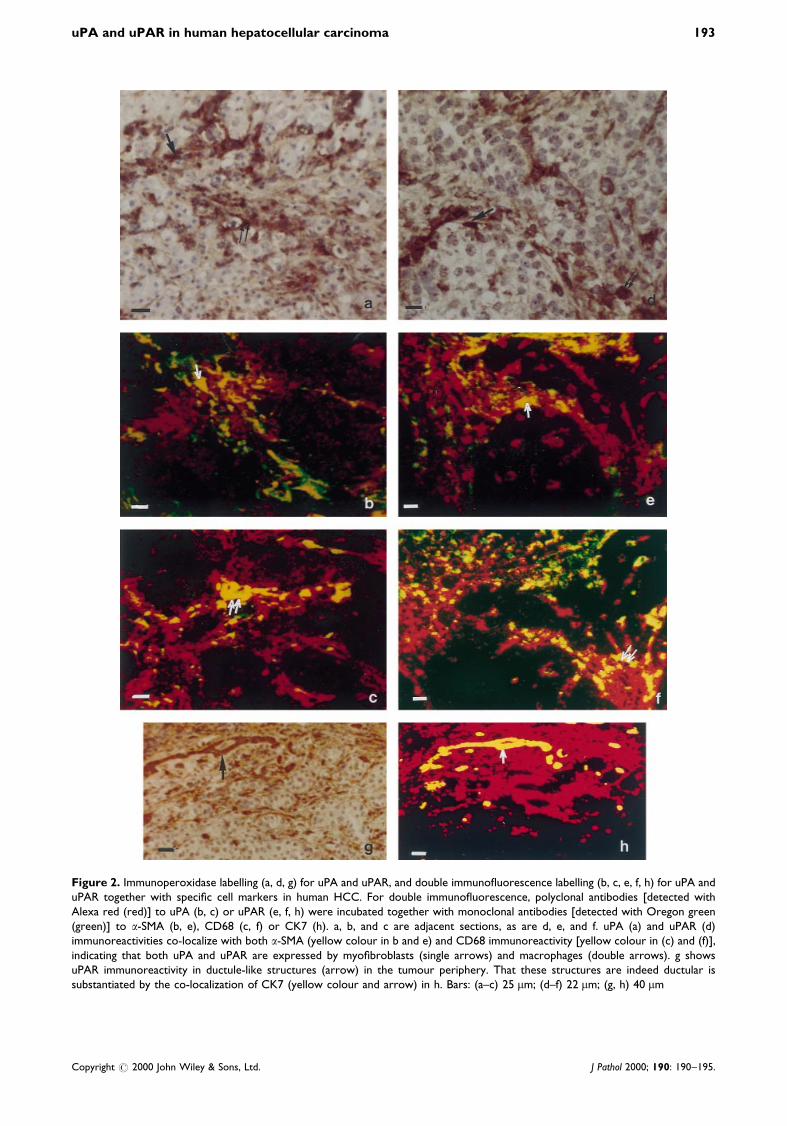
Figure 2. Immunoperoxidase labelling (a, d, g) for uPA and uPAR, and double immuno¯uorescence labelling (b, c, e, f, h) for uPA anduPAR together with speci®c cell markers in human HCC. For double immuno¯uorescence, polyclonal antibodies [detected withAlexa red (red)] to uPA (b, c) or uPAR (e, f, h) were incubated together with monoclonal antibodies [detected with Oregon green(green)] to a-SMA (b, e), CD68 (c, f) or CK7 (h). a, b, and c are adjacent sections, as are d, e, and f. uPA (a) and uPAR (d)immunoreactivities co-localize with both a-SMA (yellow colour in b and e) and CD68 immunoreactivity [yellow colour in (c) and (f)],indicating that both uPA and uPAR are expressed by myo®broblasts (single arrows) and macrophages (double arrows). g showsuPAR immunoreactivity in ductule-like structures (arrow) in the tumour periphery. That these structures are indeed ductular issubstantiated by the co-localization of CK7 (yellow colour and arrow) in h. Bars: (a±c) 25 mm; (d±f) 22 mm; (g, h) 40 mm
uPA and uPAR in human hepatocellular carcinoma 193
Copyright # 2000 John Wiley & Sons, Ltd. J Pathol 2000; 190: 190±195.
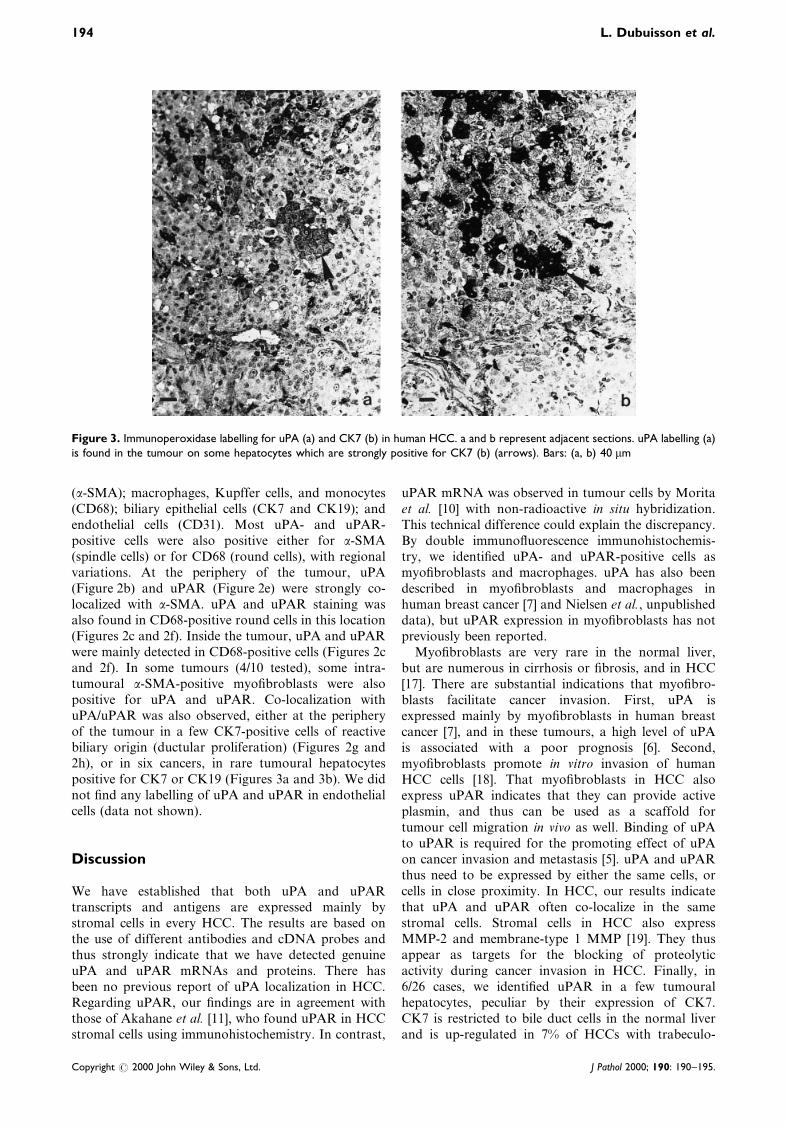
(a-SMA); macrophages, Kupffer cells, and monocytes(CD68); biliary epithelial cells (CK7 and CK19); andendothelial cells (CD31). Most uPA- and uPAR-positive cells were also positive either for a-SMA(spindle cells) or for CD68 (round cells), with regionalvariations. At the periphery of the tumour, uPA(Figure 2b) and uPAR (Figure 2e) were strongly co-localized with a-SMA. uPA and uPAR staining wasalso found in CD68-positive round cells in this location(Figures 2c and 2f). Inside the tumour, uPA and uPARwere mainly detected in CD68-positive cells (Figures 2cand 2f). In some tumours (4/10 tested), some intra-tumoural a-SMA-positive myo®broblasts were alsopositive for uPA and uPAR. Co-localization withuPA/uPAR was also observed, either at the peripheryof the tumour in a few CK7-positive cells of reactivebiliary origin (ductular proliferation) (Figures 2g and2h), or in six cancers, in rare tumoural hepatocytespositive for CK7 or CK19 (Figures 3a and 3b). We didnot ®nd any labelling of uPA and uPAR in endothelialcells (data not shown).
Discussion
We have established that both uPA and uPARtranscripts and antigens are expressed mainly bystromal cells in every HCC. The results are based onthe use of different antibodies and cDNA probes andthus strongly indicate that we have detected genuineuPA and uPAR mRNAs and proteins. There hasbeen no previous report of uPA localization in HCC.Regarding uPAR, our ®ndings are in agreement withthose of Akahane et al. [11], who found uPAR in HCCstromal cells using immunohistochemistry. In contrast,
uPAR mRNA was observed in tumour cells by Moritaet al. [10] with non-radioactive in situ hybridization.This technical difference could explain the discrepancy.By double immuno¯uorescence immunohistochemis-try, we identi®ed uPA- and uPAR-positive cells asmyo®broblasts and macrophages. uPA has also beendescribed in myo®broblasts and macrophages inhuman breast cancer [7] and Nielsen et al., unpublisheddata), but uPAR expression in myo®broblasts has notpreviously been reported.
Myo®broblasts are very rare in the normal liver,but are numerous in cirrhosis or ®brosis, and in HCC[17]. There are substantial indications that myo®bro-blasts facilitate cancer invasion. First, uPA isexpressed mainly by myo®broblasts in human breastcancer [7], and in these tumours, a high level of uPAis associated with a poor prognosis [6]. Second,myo®broblasts promote in vitro invasion of humanHCC cells [18]. That myo®broblasts in HCC alsoexpress uPAR indicates that they can provide activeplasmin, and thus can be used as a scaffold fortumour cell migration in vivo as well. Binding of uPAto uPAR is required for the promoting effect of uPAon cancer invasion and metastasis [5]. uPA and uPARthus need to be expressed by either the same cells, orcells in close proximity. In HCC, our results indicatethat uPA and uPAR often co-localize in the samestromal cells. Stromal cells in HCC also expressMMP-2 and membrane-type 1 MMP [19]. They thusappear as targets for the blocking of proteolyticactivity during cancer invasion in HCC. Finally, in6/26 cases, we identi®ed uPAR in a few tumouralhepatocytes, peculiar by their expression of CK7.CK7 is restricted to bile duct cells in the normal liverand is up-regulated in 7% of HCCs with trabeculo-
Figure 3. Immunoperoxidase labelling for uPA (a) and CK7 (b) in human HCC. a and b represent adjacent sections. uPA labelling (a)is found in the tumour on some hepatocytes which are strongly positive for CK7 (b) (arrows). Bars: (a, b) 40 mm
194 L. Dubuisson et al.
Copyright # 2000 John Wiley & Sons, Ltd. J Pathol 2000; 190: 190±195.

glandular morphology [20], in association with apoorer prognosis.
The presence of the two key elements of theplasminogen activator system mainly in stromal cellsin HCC suggests the implication of these cells intumour progression and metastasis.
Acknowledgements
This work was supported by Ligue contre le Cancer (ComiteÂs
Gironde et Dordogne), Association pour la Recherche sur le
Cancer, and FeÂdeÁration des Groupements d'Entreprises Fran-
cËaises pour la Lutte contre le Cancer. AM was the recipient of a
fellowship from La Ligue contre le Cancer (Comite Dordogne).
We thank the members of the Bordeaux Liver Transplantation
Unit for providing liver samples.
References
1. Danù K, Rùmer J, Nielsen BS, et al. Cancer invasion and tissue
remodeling cooperation of protease systems and cell types.
APMIS 1999; 107: 120±127.
2. Lyons RM, Gentru LE, Purchio AF, et al. Mechanism of
activation of latent recombinant transforming growth factor-1 by
plasmin. J Cell Biol 1990; 110: 1361±1367.
3. Mars WM, Zarnegar R, Michalopoulos GK. Activation of
hepatocyte growth factor by the plasminogen activators uPA and
tPA. Am J Pathol 1990; 143: 949±958.
4. Behrendt N, Stephens R. The urokinase receptor. Fibrinolysis
Proteolysis 1999; 12: 191±204.
5. Andreasen PA, Kjùller L, Christensen L, Nielsen LS, Duffy MJ.
The urokinase-type plasminogen activator system in cancer
metastasis: a review. Int J Cancer 1997; 72: 1±22.
6. Grùndahl-Hansen J, Peters HA, van Putten WLJ, Look MP,
Pappot H. Prognostic signi®cance of the receptor for urokinase
plasminogen activator in breast cancer. Clin Cancer Res 1995; 1:
163±173.
7. Nielsen BS, Sehested M, Timshel S, Pyke C, Danù K. Messenger
RNA for urokinase plasminogen activator is expressed in
myo®broblasts adjacent to cancer cells in human breast cancer.
Lab Invest 1996; 74: 168±177.
8. Monvoisin A, Neaud V, Dubuisson L, et al. Direct evidence that
hepatocyte growth factor-induced invasion of hepatocellular
carcinoma cells is mediated by urokinase. J Hepatol 1999; 30:
511±518.
9. De Petro G, Tavian D, Copeta A, Portolani N, Giulini SM,
Barlati S. Expression of urokinase-type plasminogen activator
(u-PA), u-PA receptor, and tissue-type PA messenger RNAs in
human hepatocellular carcinoma. Cancer Res 1998; 58:
2234±2239.
10. Morita Y, Hayashi Y, Wang Y, et al. Expression of urokinase-
type activator receptor in hepatocellular carcinoma. Hepatology
1997; 25: 856±860.
11. Akahane T, Ishii M, Ohtani H, Nagura H, Toyota T. Stromal
expression of urokinase-type plasminogen activator receptor
(uPAR) is associated with invasive growth in primary liver
cancer. Liver 1998; 18: 414±419.
12. Edmonson HA, Steiner PE. Primary carcinoma of the liver: a
study of 100 cases among 48,900 necropsies. Cancer 1954; 7:
462±503.
13. Ishak KG, Anthony PP, Sobin LM. Histological Typing of
Tumours of the Liver (2nd edn). World Health Organization,
International Histological Classi®cation of Tumours. Springer-
Verlag: Berlin, 1994.
14. Pyke C, Kristensen P, Ralfkiaer E, et al. Urokinase-type
plasminogen activator is expressed in stromal cells and its
receptor in cancer cells at invasive foci in human colon
adenocarcinomas. Am J Pathol 1991; 138: 1059±1067.
15. Rùnne E, Hùyer-Hansen G, BruÈnner N, et al. J. Urokinase
receptor in breast cancer tissue extracts. Enzyme-linked immu-
nosorbent assay with a combination of mono- and polyclonal
antibodies. Breast Cancer Res Treat 1995; 33: 199±207.
16. Le Moine C, Bloch B. D1 and D2 dopamine receptor gene
expression in the rat striatum: sensitive cRNA probes demon-
strate prominent segregation of D1 and D2 mRNAs in distinct
neuronal populations of the dorsal and ventral striatum. J Comp
Neurol 1995; 355: 418±426.
17. Schmitt-GraÈff A, KruÈger S, Bochard F, et al. Modulation of
alpha smooth muscle actin and desmin expression in perisinu-
soidal cells of normal and diseased human liver. Am J Pathol
1991; 138: 1233±1242.
18. Neaud V, Faouzi S, Guirouilh J, et al. Human hepatic
myo®broblasts increase invasiveness of hepatocellular carcinoma
cells: evidence for a role of hepatocyte growth factor. Hepatology
1997; 26: 1458±1466.
19. TheÂret N, Musso O, L'Helgouac'h A, et al. Differential
expression and origin of membrane-type 1 and 2 matrix
metalloproteinases (MT-MMPs) in association with MMP2
activation in injured human livers. Am J Pathol 1998; 153:
945±954.
20. Maeda T, Kajiyama K, Adachi E, et al. The expression of
cytokeratin 7, 19 and 20 in primary and metastatic carcinomas of
the liver. Mod Pathol 1996; 9: 901±909.
uPA and uPAR in human hepatocellular carcinoma 195
Copyright # 2000 John Wiley & Sons, Ltd. J Pathol 2000; 190: 190±195.























![Comportement sous irradiation des géopolymères · [2] Phair et al (2002) Effect of the silicate activator pH on the microstructural characteristics of waste-based geopolymer. International](https://img.pdfslide.fr/doc/110x75/601776993a62b811e91e2dd0/comportement-sous-irradiation-des-gopolym-2-phair-et-al-2002-effect-of-the.jpg)





