Embed Size (px)
Citation preview

Vol. 167, No. 2, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
March 16, 1990 Pages 784-789
EXPRESSION OF THE BROWN FAT MITOCHONDRIA UNCOUPLING PROTEIN IN XENOPUS
OOCYTES AND IMPORT INTO MITOCHONDRIAL MEMBRANE
Susanne Klaus, Louis Casteilla, Frederic Bouillaud, Serge Raimbault, and Daniel Ricquier
Centre National de la Recherche Scientifique
Centre de Recherches sur la Nutrition, 9, rue Jules Hetzel
F-921 90 Meudon-Bellevue, France
Received January 23, 1990
Non shivering thermogenesis of brown adipose tissue is due to the uncoupling protein (UCP), located in the inner mitochondrial membrane, which functions as a proton translocator and can thus uncouple mitochondrial respiration. We describe here the expression of UCP in Xenopus laevis oocytes after injection of UCP mRNA, which was transcribed in vitro. UCP seems to be correctly transported into mitochondria and integrated into the membrane, but we were not able to establish definitely the functionality of this UCP. We conclude that this expression system could be suitable for the study of the mitochondrial import mechanism but not for the examination of physiological properties of UCP. 01990 Academic mess, 1°C.
The mitochondrial uncoupling protein (UCP) is responsible for the heat dissipating ability of
brown adipose tissue (BAT) by its function as a proton translocator and thus its ability to
uncouple mitochondrial respiration from ATP synthesis. This 33 kD protein is located ex-
clusively in mitochondria of BAT and spans as a dimer the inner mitochondrial membrane. Its
proton translocating activity is regulated positively by fatty acids and negatively by purine
di- or triphosphate nucleotides (1). UCP is coded for by a nuclear gene which seems to exist
as only one copy and is expressed only in BAT (2,3). Its primary sequence (4,5,6) shows
that UCP belongs to a protein family including the ADP/ATP carrier (AAC)(G) and the
mitochondrial phosphate carrier (7), which are mitochondrial carrier proteins, and a very
recently identified protein called hML-7, whose function is not yet known (8). Like AAC and
hML-7 this mitochondrial protein does not possess a precursor with a transient, targeting
signal sequence (4,5) and despite some studies on AAC and UCP import (9,iO) the regions
implicated in the import are not yet well determined. Moreover, the molecular mechanism of
the proton translocation and its regulation by nucleotides and fatty acids are not yet well
understood and no molecular study has been made to characterize them. The integration of
UCP into the inner mitochondrial membrane as a prerequisite for its physiological function
complicates the study of functional domains of UCP. We have tried to find an expression
0006-291X/90 $1.50
Copyright 0 1990 by Academic Press, Inc.
All rights of reproduction in any form reserved. 784

Vol. 167, No. 2, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
system for UCP, able to process correctly the protein and containing numerous
mitochondria, in order to study UCP import in vitro and eventually to elucidate the
relationships between UCP structure and function. Among different expression sytems,
Xenopus oocytes are a useful tool for the study of protein expression and function. They do
not only translate injected mRNAs very efficiently, but they also carry out correctly
numerous post-translational modifications of proteins and can direct them into the correct
intracellular membranes (11). In this study we describe the expression of UCP in Xenopus
oocytes and its import into oocyte mitochondrial membranes. To our knowledge this system
has not yet been used for the study of mitochondrial membrane carrier proteins. It is also the
first report of UCP expression in an eukaryotic expression system.
MATERIALS AND METHODS
uncoupling orotein mRNA: The complete UCP cDNA was cloned into pTZ 19 (Pharmacia) behind the promoter T7 (fig. la). The plasmid was linearized by Eco Rl and transcription in vitro was performed with T7 polymerase using the mCAP kit from Stratagene according to the instructions. Functionality of the in vitro transcription product was tested by per- forming a cell-free in vitro translation using a kit from Bethesda Research Laboratories. The translation products were analyzed by SDS PAGE and autoradiography. Isolation and injection of oocytes; Oocytes were obtained from female Xenopus lawis kept in a watertank at 15 “C. Animals were put in icewater for anaesthesia, and lobes of ovaries were removed according to need. Ovaries were washed and stored in a modified Barth’s saline (OR2: 82.5 mM NaCI, 2.5 mM KCI, 1 mM CaCl2, 1 mM MgCI2, 1 mM Na2HP04, 5 mM Hepes, 100 U/ml Streptomycin and Penicillin G, pH 7.5). Individual, mature oocytes were dissected manually. For UCP expression 50 nl RNA (lpg/trI) were microinjected into the vegetative pole, and oocytes were incubated individually at 17 “C in OR2 which was renewed every day. Control oocytes were injected with 50 nl of sterile water and treated in the same way. Isolation of mitochondria; Mitochondria were isolated by differential centrifugation in 250 mM Sucrose, 10 mM TRIS and 1 mM EDTA, pH 7.2. Oocytes were crushed and suspended in a large volume of isolation buffer. Two subsequent 5 min. centrifugations at 8009 and 25009, respectively, were performed after which the pellet was discarded. After a final 20 min. centrifugation at 90009 the pellet was resuspended in a small volume of isolation buffer. Purification of mitochondria was followed by monitoring cytochrome c oxidase activity spectrophotometrically (12). Routinely a lo- to 15- fold purification and a yield between 10 and 20% were obtained by this method. Immunoblou For Western blotting proteins were separated by SDS PAGE at 12% acrylamide and electrotransfered onto a nitrocellulose membrane. lmmunoblotting was performed using an antibody raised against purified rat UCP, with a second antibody coupled to peroxidase (13).
RESULTS AND DISCUSSION
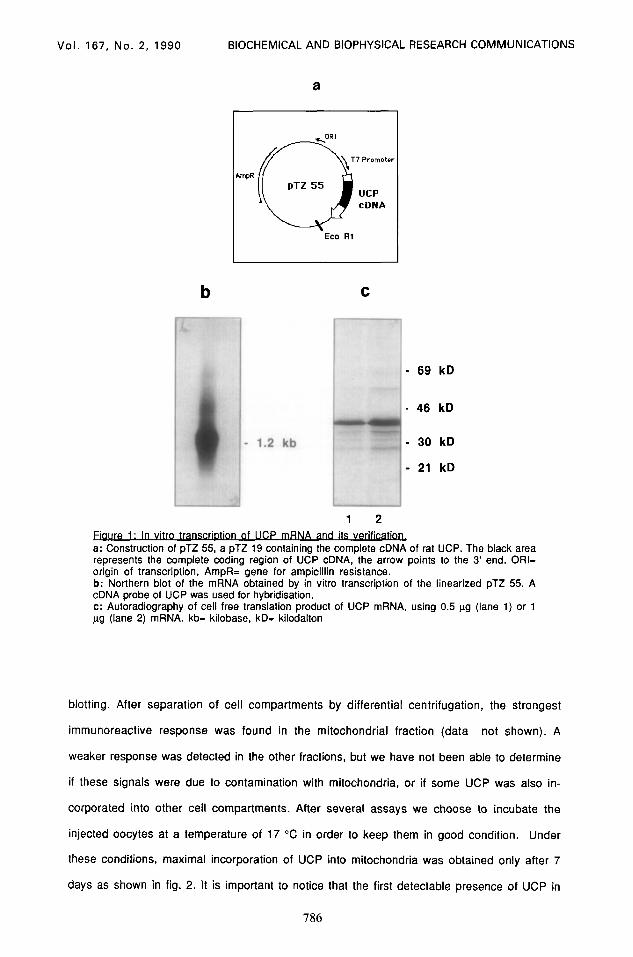
The in vitro transcription of the UCP cDNA by means of the T7 polymerase produced a mRNA
of the expected size (about 1.2 kb), which was translated into a protein of 33 kD (fig 1).
Routinely between 12 and 15 trg RNA were obtained from1 ug plasmid. Injected Xenopus
oocytes were able to translate this mRNA into uncoupling protein, as revealed by immuno-
785
Vol. 167, No. 2, 1990 BIOCHEMICAL AND RIOPHYSICAL RESEARCH COMMUNICATIONS
a
-Em Rl
. 1.2 kb
- 69 kD
- 46 kD
- 30 kD
I - 21 kD
1 2 . . . . .
Fiaure 1. In vitro transcrlDtlon its vehmm a: Construction of pTZ 55, a pTZ 19 containing the complete cDNA of rat UCP. The black area represents the complete coding region of UCP cDNA, the arrow points to the 3’ end. ORI= origin of transcription, AmpR= gene for ampicillin resistance. b: Northern blot of the mRNA obtained by in vitro transcription of the linearized pTZ 55. A cDNA probe of UCP was used for hybridisation. c: Autoradiography of cell free translation product of UCP mRNA, using 0.5 ug (lane 1) or 1 ug (lane 2) mRNA. kb= kilobase, kD= kilodalton
blotting. After separation of cell compartments by differential centrifugation, the strongest
immunoreactive response was found in the mitochondrial fraction (data not shown). A
weaker response was detected in the other fractions, but we have not been able to determine
if these signals were due to contamination with mitochondria, or if some UCP was also in-
corporated into other cell compartments. After several assays we choose to incubate the
injected oocytes at a temperature of 17 “C in order to keep them in good condition. Under
these conditions, maximal incorporation of UCP into mitochondria was obtained only after 7
days as shown in fig. 2. It is important to notice that the first detectable presence of UCP in
786

Vol. 167, No. 2, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
- 33 kD
2 4 7 7c BAT days
lmmunoblotting of mitochondria of oocytes injected with 50 ng UCP mRNA and incubated for 2, 4, or 7 days, respectively. Each lane corresponds to mitochondria of eight oocytes. c= control oocytes, injected with water, BAT= 4 f.rg mitochondrial protein of brown adipose tissue of cold-acclimated rats.
mitochondria was found after 2 days. This latency apparently corresponds to the time
necessary for translation of the mRNA and processing of the protein. In order to investigate
the correct targeting of UCP into mitochondrial membranes, mitochondria were disrupted
either by suspension in a hypoosmotic medium or by sonication (followed in both cases by
freezing and rethawing). The membranes were then collected by ultracentrifugation and
subjected to immunoblotting (fig. 3a). We still found a strong immunoreactive signal of the
disintegrated membranes. We therefore concluded that nearly all UCP detected in the mito-
chondrial fraction was integrated into membranes. Further evidence was obtained by
trypsin, i.e. proteolytic treatment of the mitochondria. After mitochondria were incubated
with trypsin for 30 or 60 minutes, UCP could still be detected (fig 3b). This shows clearly
that the whole information for the targeting of UCP into mitochondria is still present in
artificially produced mRNA and that no mechanism specific for brown adipocytes is
necessary for integration of UCP into mitochondrial membranes. It is not yet clear which
exact signal sequences are involved in the targeting of UCP or AAC into mitochondria and
their insertion into the inner membrane. Recent findings (14) point to the amino terminal
one third of UCP as being essential for both import and membrane insertion. The authors
obtained this information from transcription of plasmids containing complete or partially
deleted UCP cDNA, followed by translation in vitro and incorporation of the obtained protein
into isolated heart muscle mitochondria. They also suggested the presence of a second import
signal within the carboxyl-terminal two thirds, which does not mediate membrane insertion.
As for AAC, different groups suggest that domains distant from the amino terminus may carry
specific targeting information (9,lO). The expression of modified UCP in an eucaryotic
787

Vol. 167, No. 2, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
a b
- 33 kD
M H S 0 30 60 6Oc
minutes
Ewe 3: lntearation of UCP IntO mltochondrial lmmunoblotting of mitochondrial membranes of oocytes injected with 50 ng UCP mRNA. a: mitochondria were dissrupted by suspension in hypotonic medium (TRIS lOmM, EDTA 1 mM, pH 7.2) followed by freezing and rethawing (H) or sonicated 2x20 set followed by freezing and rethawing (S). Membranes were then collected by ultracentrifugation at 150000g for 20 min. M= intact mitochondria. Each lane corresponds to mitochondria of 15 oocytes. b: mitochondria were incubated at 37 “C with trypsin (0.25 pg/kg protein) for 0, 30, or 60 minutes. The reaction was stopped by adding trypsin inhibitor (0.5 pgglpg protein). 6Oc = mitochondria incubated for 60 min without trypsin. Each lane corresponds to mitochon- dria of 13 oocytes.
expression system could prove valuable for further examination of mitochondrial import
mechanism.
We tried to examine the functionality of UCP, expressed in Xenopus oocytes, by doing
respiration studies on isolated mitochondria. We did not succeed because of several
experimental problems. The purification of oocyte mitochondria is made difficult by the high
content of vitellogenin which seems to trap mitochondria, resulting in a very low yield. We
also observed that oocyte mitochondria hardly ever showed a coupled respiration, which
would be necessary to study the uncoupling properties of UCP. Regarding the nucleotide
binding capacity of UCP, we performed photoaffinity labeling of oocyte mitochondria (data
not shown). The results seemed to indicate that UCP expressed in oocytes is able to bind ATP
and GDP. However, these assays were hindered by the contamination of the mitochondrial
fraction with vitellogenin, and were thus not conclusive.
CONCLUSIONS
We describe here for the first time the expression of UCP, a mitochondrial carrier protein,
in an eukaryotic expression system, the Xenopus oocyte. This system is not only able to
express UCP, but also to direct it into mitochondria and insert it into the membrane.
However, examination of the functionality of UCP in oocyte mitochondria was not successful.
788

Vol. 167, No. 2, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
We conclude that this system might be useful to study the import mechanism of the protein
into mitochondria, but it does not seem suitable for the study of UCP function. We recently
obtained the expression of UCP in an other eukaryotic expression system, a fibroblast cell
line, in which tests of UCP functionality have been positive (Ricquier et al, in preparation).
We thus have a second eukaryotic expression system avaible, which seems to be more
promising for the study of protein domains involved in nucleotide binding and proton
ACKNOWLEDGMENTS
We wish to thank Dr. Raymond T. Kado for his advise concerning the oocyte injection, as well as for his comments and discussion. This study was supported by CNRS and MRES. S. Klaus was supported by a postdoctoral fellowship from the Deutsche Forschungsgemeinschaft.
REFERENCES
1. Nicholls, D.G., and Locke, R.M. (1984) Physiol. Rev. 64, l-64. 2. Bouillaud, F., Raimbault, S., and Ricquier, D. (1988) Biochem. Biophys. Res. Comm. 157,
783-792. 3. Kozak, L..P., Britton, J.H., Kozak, U.C., Wells, J.M. (1988) J. Biol. Chem. 263, 12274-
12277 4. Bouillaud, F., Weissenbach, J., and Ricquier, D. (1986) J; Biol. Chem. 261, 1487-1490. 5. Aquila, H., Link, T.A., Klingenberg, M. (1985) EMBO J. 4, 2369-2376. 6. Ridley, R.G., Patel, H.V., Gerber, G.E., Morton, R.C., and Freeman, K.B. (1986) Nucl. Acid
Res. 14, 4025-4035. 7. Runswick, M.J., Powell, J.T., Nyren, P., and Walker, J.E. (1987) EMBO J. 6,1367-1373. 8. Zarrilli, R., Oates, E.L., McBride, O.W., Lerman, M.I., Chart, J.Y., Santisteban, P., Ursini,
M.V., Notkins, A.L., and Kohn, L.D. (1989) Mol. Endocrinol. 3, 1498-1508 9. Pfanner, N., Hoeben, P. ,Tropschung, M., and Neupert, W. (1987) J. Biol. Chem. 31,
14851-14854. 10. Smagula, C. and Douglas, M.G. (1988) J. Biol. Chem. 263, 6783-6790. 11. Colman, A. (1984) Transcription and translation. A practical approach, IRL Press. Oxford,
Washington DC. pp 271-302. 12. Wharton, D.C., and Tzagoloff, A. (1967) Meth. Enzymol. 10, 245-250. 13. Ricquier, D., Barlet, J.P., Garel, J.M., Combes-George, M., Dubois, M.P. (1983)
Biochem. J. 210, 859-866. 14. Liu, X., Bell, A.W., Freeman, K.B., and Shore, G.C. (1988) J. Cell Biol. 107, 503-509.
789