Embed Size (px)
Citation preview

1/
nce
ymptotice and thehe divisionwhere the
neoduit d’unedonné par
nt ou non-dire pour
ctéries etL’objectifn modélise
C. R. Acad. Sci. Paris, Ser. I 341 (2005) 731–736http://france.elsevier.com/direct/CRASS
Partial Differential Equations
Fitness optimization in a cell division modelPhilippe Michela,b
a Département de mathématiques et applications, UMR 8553, École normale supérieure, 45, rue d’Ulm, 75230 Paris cedex 05, Frab CEREMADE, Paris Dauphine, place de Lattre de Tassigny, 75775 Paris cedex 16, France
Received 19 September 2005; accepted 11 October 2005
Available online 11 November 2005
Presented by Carlo Cercignani
Abstract
We consider a size structured cell population model where a mother cell gives birth to two cells. We know that the asbehavior of the density of cells is given by the solution to an eigenproblem. The eigenvector gives the asymptotic shapeigenvalue gives the exponential growth rate and so the Maltusian parameter. The Maltusian parameter depends on trule for the mother, i.e., symmetric (the two daughter cells have the same size) or asymmetric. We give some examplesymmetrical division is not the best fitted division, i.e., the Maltusian parameter is not optimal.To cite this article: P. Michel, C. R.Acad. Sci. Paris, Ser. I 341 (2005). 2005 Académie des sciences. Published by Elsevier SAS. All rights reserved.
Résumé
Optimisation du taux de croissance dans un modèle de division cellulaire.Nous considérons un modèle d’évolution d’udensité de cellules structurée en taille. Nous savons que la densité de cellules se comporte en temps long comme le prfonction indépendante du temps et d’une exponentielle dépendant du temps. Le taux de croissance exponentiel étantun paramètre Maltusien dont nous allons montrer qu’il depend de la manière dont la cellule se divise, symétriquemesymétriquement. On donnera des exemples pour lesquels la division symétrique n’est pas la mieux adaptée, c’est-àlesquels le taux de croissance exponentiel n’est pas maximal.Pour citer cet article : P. Michel, C. R. Acad. Sci. Paris, Ser. I 341(2005). 2005 Académie des sciences. Published by Elsevier SAS. All rights reserved.
Version française abrégée
La division cellulaire asymétrique est observée pour les levures (par bourgeonement), pour certaines bad’autres cellules. Elle peut être expliquée par une meilleure capacité de développement (sélection naturelle).de cette note est de montrer que certains modèles mathématiques peuvent représenter ce phénomène. Ol’évolution d’une densité de cellulesn(t, y) structurée en tailley, en un tempst , par l’équation,
∂n
∂t(t, y) + ∂nV (y)
∂y(t, y) + B(y)n(t, y) =
∞∫y
b(y′, y)n(t, y′)dy′, y � 0,
n(·,0) = 0,
(1)
E-mail address:[email protected] (P. Michel).
1631-073X/$ – see front matter 2005 Académie des sciences. Published by Elsevier SAS. All rights reserved.doi:10.1016/j.crma.2005.10.012

732 P. Michel / C. R. Acad. Sci. Paris, Ser. I 341 (2005) 731–736
trs
ifié
e et
i-
ision inyeasts,
ve birthon is notion anddivision
où V est le taux de croissance,B le taux de naissance etb la manière dont une cellule de tailley′ se divise. On sai(voir [8,9]) qu’en temps longn(t, ·) se comporte comme eλtN(·), oùN etλ sont solutions d’un problème aux valeupropres. Nous allons montrer à travers ce modèle simple de division cellulaire que le taux de croissanceλ depends dela manière dont la cellule se divise. Par exemple on peut se restreindre à la paramétrisation par un paramètreσ ∈ ]0,1[qui exprime la division en deux cellules de tailleσy et (1− σ)y. Nous nous intéressons donc au problème simpl
∂n
∂t(t, y) + ∂nV (y)
∂y(t, y) + B(y)n(t, y) = 1
σB
(y
σ
)n
(t,
y
σ
)+ 1
1− σB
(y
1− σ
)n
(t,
y
1− σ
). (2)
On sait (voir [9]) quen(t, y) ∼t→∞ ρNσ (y)eλσ t , où ρ est une constante dépendant de la condition initial(λσ ,Nσ ,φσ ) est une solution d’un problème aux valeurs propres et de son dual, défini par
∂Nσ
∂y+ (
B(y) + λσ
)Nσ (y) = 1
σB
(y
σ
)Nσ
(y
σ
)+ 1
1− σB
(y
1− σ
)Nσ
(y
1− σ
),
−∂φσ
∂y+ (
B(y) + λσ
)φσ (y) = B(y)
[φσ (σy) + φσ
((1− σ)y
)],
Nσ (0) = 0, Nσ (y) � 0, φσ (y) � 0, ∀y � 0,
∞∫0
Nσ (y′)φσ (y′)dy′ = 1.
(3)
On obtient alors
Théorème 0.1.Soit(λσ ,Nσ ,φσ ) solution de(3) alors
(∂
∂σλσ
)σ=σ0
=∞∫
0
B(y)yNσ0(y)
[(∂
∂y
φσ0
)(yσ0) −
(∂
∂y
φσ0
)(y(1− σ0)
)]dy. (4)
De plus si(( ∂∂y
φσ0)(yσ0) − ( ∂∂y
φσ0)(y(1− σ0))) = 0 pour touty alors
(∂2
∂σ∂σλσ
)σ=σ0
=∞∫
0
B(y)y2[(
∂2
∂y∂yφσ0
)(yσ0) +
(∂2
∂y∂yφσ0
)(y(1− σ0)
)]Nσ0(y)dy. (5)
Par conséquent on a
Théorème 0.2. Si V = 1 et K = SuppB est un intervalle tel que(σ0K)⋂
K = ∅ avec σ0 ∈ ]1/2,1[ alors(∂σλσ )σ=σ0 > 0. En particulier pourK = SuppB ⊂ [a, b] aveca > b/2, la division symétrique n’est pas la divsion la mieux adaptée, i.e., il existeσ > 1/2 tel queλσ > λ1/2.
SiV = 1 etK = SuppB = [0, β] tel que
λσ (λσ − B(0))
B(0)� B ′(y)
B(y)� 0, ∀y ∈ ]0, β[,
alors la division symétrique est la mieux adaptée, i.e., pour toutσ ∈ ]0,1[, λσ � λ1/2. En particulier pourB = 1[0,β]avecβ > 0 on aλ1/2 � λσ pour toutσ dans]0,1[.
1. Introduction
General models of cell division have been known for a long time. Although the most classical case is divtwo equal new cells, it is now well established that this is not always the case. In the large class of buddingE. Coli or for some bacteria or for instance Physcomitrella protoplast, division is not always symmetric and gito a bigger and a smaller cell. We can explain this by adaptive dynamic. In some cases, the symmetric divisithe best fitted way of division. Our aim is to exhibit a parameter that characterizes the fitness for a cell divisto optimize it under some constraints on the cell division parameter. The natural model to study it, is a cell

P. Michel / C. R. Acad. Sci. Paris, Ser. I 341 (2005) 731–736 733
e
rowthon
[9])
he
eter and
if
ted
model (see [2,7]) in which the density of cellsn(t, y) is structured by their sizey and evolution is describes by thmaster equation
∂n
∂t(t, y) + ∂nV (y)
∂y(t, y) + B(y)n(t, y) = 1
σB
(y
σ
)n
(t,
y
σ
)+ 1
1− σB
(y
1− σ
)n
(t,
y
1− σ
), (6)
where a cell of sizey gives birth to a cell of sizeyσ and an another of size(1− σ)y, with σ ∈ ]0,1[, i.e.,b(y, y′) =B(y′)[δy′=σy + δy′=(1−σ)y]. A Similar model also arises to describe fragmentation in physics [4,6] and the gterm∂yn arises after rescaling [1,3]. The asymptotic behavior ofn(t, y) gives the invasive capacity of the populatiand thus a fitness measure of populations under different rates and probabilities in (6).
2. Asymptotic result and definition of a fitness parameter
We recall the result of asymptotic behavior ofn(t, y) solution to (6), given by the General Relative Entropy (seen(t, y) ∼t→∞ ρNσ (y)eλσ t , whereλσ is the first eigenvalue associated to the following eigenproblem
∂V Nσ
∂y+ (
B(y) + λσ
)Nσ (y) = 1
σB
(y
σ
)Nσ
(y
σ
)+ 1
1− σB
(y
1− σ
)Nσ
(y
1− σ
),
−V (y)∂φσ
∂y+ (
B(y) + λσ
)φσ (y) = B(y)
(φσ (σy) + φσ
((1− σ)y
)),
Nσ (0) = 0, Nσ (y) � 0, φσ (y) � 0, ∀y � 0,
∞∫0
Nσ (y′)φσ (y′)dy′ = 1,
(7)
andρ is a constant depending on the initial data. We notice thatφσ is a solution to the dual eigenproblem of teigenproblem inNσ . This means thatλσ is the Maltusian parameter that gives the exponential growth andNσ isthe asymptotic shape. Our point here is to focus on the Maltusian parameter that gives an invasive paramwe chooseλσ as the fitness parameter. Therefore we fix the birth rateB and the speed growthV , and we study thevariation ofλσ with respect to the parameterσ that describes all sizes after division.
3. Dependence of the fitness parameter with respect toσ
Here we show that the symmetric division is not necessarily the best fitted division. We study two cases,B has acompact support andB(y) = yp. We notice that we only have to considerσ ∈ [1/2,1[, indeed (7) does not changewe exchangeσ and 1− σ , therefore we haveλσ = λ1−σ . Then we have the following results.
3.1. AssumeB has a compact support
In this case, symmetric division might or not be the best fitted division. More precisely, we have
Theorem 3.1.AssumeV = 1, K = SuppB = [a, b] ⊂ [0,∞[ s.t.
(σ0K) ∩ K = ∅,
with σ0 ∈ ]1/2,1[ then ( ∂∂σ
λσ )σ=σ0 > 0. In particular, if a > b/2 then the symmetric division is not the best fitdivision, i.e., there existsσ > 1/2 s.t.λσ > λ1/2.
AssumeV = 1 andSuppB = [0, β] s.t.B ≡ 0, decreases and satisfies
λσ (λσ − B(0))
B(0)� B ′(y)
B(y)� 0, ∀y ∈ ]0, β[,
then the symmetric division is the best fitted division, i.e., for allσ ∈ ]0,1[, we haveλσ � λ1/2. In particular forB = 1[0,β], with β > 0, we haveλ1/2 � λσ for all σ in ]0,1[.
Numerically, we find, by settingb = 1 anda = 0, 0.1, 0.5 thatλσ is increasing on[1/2,1[ for a = 0, reaches amaximum atσ0 ∈ ]1/2,1[ for a = 0.1 and is decreasing on[1/2,1[ for a = 0.5 (see Fig. 1).
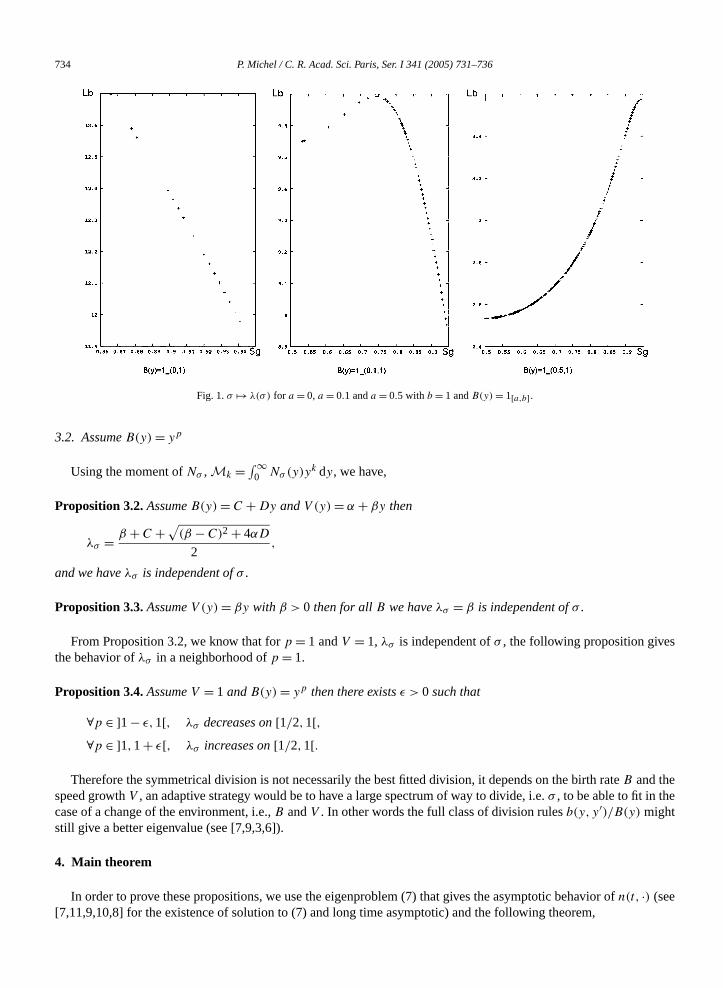
734 P. Michel / C. R. Acad. Sci. Paris, Ser. I 341 (2005) 731–736
s
Fig. 1.σ �→ λ(σ) for a = 0, a = 0.1 anda = 0.5 with b = 1 andB(y) = 1[a,b].
3.2. AssumeB(y) = yp
Using the moment ofNσ , Mk = ∫ ∞0 Nσ (y)yk dy, we have,
Proposition 3.2.AssumeB(y) = C + Dy andV (y) = α + βy then
λσ = β + C + √(β − C)2 + 4αD
2,
and we haveλσ is independent ofσ .
Proposition 3.3.AssumeV (y) = βy with β > 0 then for allB we haveλσ = β is independent ofσ .
From Proposition 3.2, we know that forp = 1 andV = 1, λσ is independent ofσ , the following proposition givethe behavior ofλσ in a neighborhood ofp = 1.
Proposition 3.4.AssumeV = 1 andB(y) = yp then there existsε > 0 such that
∀p ∈ ]1− ε,1[, λσ decreases on[1/2,1[,∀p ∈ ]1,1+ ε[, λσ increases on[1/2,1[.
Therefore the symmetrical division is not necessarily the best fitted division, it depends on the birth rateB and thespeed growthV , an adaptive strategy would be to have a large spectrum of way to divide, i.e.σ , to be able to fit in thecase of a change of the environment, i.e.,B andV . In other words the full class of division rulesb(y, y′)/B(y) mightstill give a better eigenvalue (see [7,9,3,6]).
4. Main theorem
In order to prove these propositions, we use the eigenproblem (7) that gives the asymptotic behavior ofn(t, ·) (see[7,11,9,10,8] for the existence of solution to (7) and long time asymptotic) and the following theorem,

P. Michel / C. R. Acad. Sci. Paris, Ser. I 341 (2005) 731–736 735
ffunction
n
3.inistic
Theorem 4.1.Assume there existsλσ , Nσ , φσ solution to(7) for all σ ∈ [1/2,1[ and forσ0 ∈ [1/2,1[ we have
(∂
∂σλσ
)σ=σ0
=∞∫
0
B(y)yNσ0(y)
[(∂
∂y
φσ0
)(yσ0) −
(∂
∂y
φσ0
)(y(1− σ0)
)]dy. (8)
Moreover if we have( ∂∂y
φσ0)(yσ0) = ( ∂∂y
φσ0)(y(1 − σ0)) = 0 for all y � 0, for instance whenσ0 = 1/2, B = Cst or
B(y) = y, then
(∂2
∂σ 2λσ
)σ=σ0
=∞∫
0
B(y)y2[(
∂2
∂y2φσ0
)(yσ0) +
(∂2
∂y2φσ0
)(y(1− σ0)
)]Nσ0(y)dy. (9)
Proof of Theorem 4.1. Indeed, we haveλσ = ∫ ∞0 L∗
σ (φσ )(y)Nσ (y)dy, where the operatorLσ and its dualL∗σ are
defined by
Lσ (g) = − ∂
∂y(Vg) − B(y)g + B(y/σ)g(y/σ)
σ+ B(y/(1− σ))g(y/(1− σ))
1− σ,
L∗σ (g) = V
∂
∂yg − B(y)g + B(y)g(yσ ) + B(y)g
(y(1− σ)
).
Thus we deduce
∂
∂σλσ =
∞∫0
(∂
∂σL∗
σ
)(φσ )(y)Nσ (y) +L∗
σ
(∂
∂σφσ
)(y)Nσ (y) +L∗
σ (φσ )(y)∂
∂σNσ (y)dy,
butL∗σ (φσ )(y) = λσ φσ (y), Lσ (Nσ )(y) = λσ Nσ (y) and
∞∫0
(Nσ (y)
∂
∂σφσ (y) + φσ (y)
∂
∂σNσ (y)
)dy = ∂
∂σ
∞∫0
Nσ (y)φσ (y)dy = 0,
and so, finally we have,
∂
∂σλσ =
∞∫0
(∂
∂σL∗
σ
)(φσ )(y)Nσ (y)dy =
∞∫0
B(y)yNσ (y)
[(∂
∂yφσ
)(yσ ) −
(∂
∂yφσ
)(y(1− σ)
)]dy. �
The equality (9) covers the case where( ∂∂σ
L∗σ )σ=σ0 = 0 and thus( ∂
∂σλσ )σ=σ0 = 0 which appears forσ0 = 1/2,
therefore we need( ∂2
∂σ2 λσ )σ=σ0 to find the local behavior ofλσ in the neighborhood ofσ0. We refer to [5] for the prooof this result in the matrix case. We notice that Theorem 4.1 gives some simple conditions on the dual eigenφσ to prove the decay or the growth ofλσ .
Corollary 4.2. Let σ0 fixed in [1/2,1[. Assumeφσ0 is convex(resp. concave) then ( ∂∂σ
λσ )σ=σ0 � 0 (resp.( ∂∂σ
λσ )σ=σ0 � 0).
As a consequence we can prove the second part of Theorem 3.1. Indeed, letσ = 1/2 then using the definitioof φσ , we haveφ1/2(y) = C eλ1/2y for all y ∈ [0, b/2] with C > 0. Thusφ is convex on(1/2K) = [a/2, b/2] and
( ∂2
∂σ∂σλσ )σ=1/2 > 0 and( ∂
∂σλσ )σ=1/2 = 0, thereforeλσ is locally increasing andλ1/2 < λσ for someσ > 1/2.
References
[1] J. Bertoin, A.V. Gnedin, Asymptotic laws for nonconservative self-similar fragmentations, Electron. J. Probab. 9 (19) (2004) 575–59[2] O. Diekmann, M. Gyllenberg, H. Huang, M. Kirkilionis, J.A.J. Metz, H.R. Thieme, On the formulation and analysis of general determ
structured population models. II. Nonlinear theory, J. Math. Biol. 43 (2) (2001) 157–189.

736 P. Michel / C. R. Acad. Sci. Paris, Ser. I 341 (2005) 731–736
els, Ann.
balance
r-Verlag,
ress.
55–177.
[3] M. Escobedo, S. Mischler, M. Rodriguez Ricard, On self-similarity and stationary problem for fragmentation and coagulation modInst. H. Poincaré Non Linéaire Anal. 22 (1) (2005) 99–125.
[4] N. Fournier, S. Mischler, Exponential trend to equilibrium for discrete coagulation equations with strong fragmentation and without acondition, Proc. Roy Soc. London Ser. A 460 (2049) (2004) 2477–2486.
[5] R.A. Horn, C.R. Johnson, Matrix Analysis, Cambridge University Press, Cambridge, 1985 (Chapter 6.3, p. 372).[6] P. Laurençot, Steady states for a fragmentation equation with size diffusion, Banach Center Publ. 66 (2004) 211–219.[7] J.A.J. Metz, O. Diekmann, The Dynamics of Physiologically Structured Populations, Lecture Notes in Biomath., vol. 68, Springe
1986.[8] P. Michel, PhD Thesis, Univ. Paris 9, Dauphine, in preparation.[9] P. Michel, S. Mischler, B. Perthame, General relative entropy inequality: an illustration on growth models, J. Math. Pures Appl., in p
[10] P. Michel, Existence of a solution to the cell division eigenproblem, Math. Models Methods Appl. Sci., in press.[11] B. Perthame, L. Ryzhik, Exponential decay for the fragmentation or cell-division equation, J. Differential Equations 210 (1) (2005) 1





























