Embed Size (px)
Citation preview

ABDELLATIF MAAMRI1*, FELIX BÄRLOCHER2, ERIC PATTEE3 and HASSAN CHERGUI4
1* Hydrobiologie et Ecologie Générale, Faculté des Sciences, Université Mohamed Ier, Oujda, Moroccoe-mail: [email protected]
2 Dept. of Biology, Mt. Allison University, Sackville, N.B., Canada3 UMR Ecologie des Hydrosystèmes Fluviaux, Université Claude Bernard Lyon-I, Villeurbanne, France
4 Hydrobiologie et Ecologie Générale, Faculté des Sciences Dhar el Mehraz, Fès, Morocco
Fungal and Bacterial Colonisation of Salix pedicellataLeaves Decaying in Permanent and Intermittent Streams
in Eastern Morocco
Key words: aquatic hyphomycetes, bacteria, Salix pedicellata, decay rates, intermittent and permanentstreams
Abstract
Leaf degradation was investigated at four sites in the Zegzel/Cherraa river system (Oueds). Two sites(Upper Zegzel and Lower Zegzel) carry water throughout the year and two (Upstream and DownstreamCherraa) are dry for 5–7 months each year.
The dynamics of leaf weight loss and microorganisms associated with Salix pedicellata leaves decay-ing at the four sites were compared for the first time during the same period over this permanent andintermittent system. Overall decay rates of leaves were significantly higher in the permanent Zegzel stream sections (k = 0.0094 d–1 upstream and 0.0056 downstream) than in the intermittently flowingCherraa sites (k = 0.0046 upstream, and 0.0036 downstream). In the latter, decay was much slowerduring dry than during wet periods (Upstream Cherraa: k = 0.0028 and 0.0446, respectively; Down-stream Cherraa, k = 0.0008 and 0.0357, respectively).
Similar gradients from permanent to intermittent sites were observed in numbers of bacteria per unit areaor per weight of decaying leaves (direct counts by epifluorescence microscopy), in numbers of fungalspecies and in sporulation rates, from leaves recovered at the four sites. Ten hyphomycete species werenew for Morocco.
1. Introduction
Microorganisms play a fundamental role in the biogeochemistry of aquatic and terrestrial eco-systems, contributing to the humification and mineralisation of organic matter. However, fungiand bacteria are not simply decomposers restoring nutrients to primary producers, a role inwhich they have traditionally been identified, but also producers of biomass that is available tohigher trophic levels (GESSNER and CHAUVET, 1994). These dual roles have received consider-able attention in marine (FINDLAY and ARSUFFI, 1989; NEWELL et al., 1989) and freshwater(SRIDHAR and BÄRLOCHER, 1993; BALDY et al., 1995; MAAMRI et al., 1998 and 1999) systems.In streams, rates of leaf litter decay and production of fungal biomass and spores (conidia) arebroadly correlated (GESSNER and CHAUVET, 1994; MAHARNING and BÄRLOCHER, 1996).
Most studies on leaf decomposition in streams have been conducted in temperate, per-manently flowing waters. These processes have been studied in streams in North-EasternMorocco, where climate is dry Mediterranean, by CHERGUI and PATTEE (1992) and MAAMRI
et al. (1997a). A consequence of this climate is that many streams dry out for several months
Internat. Rev. Hydrobiol. 86 2001 3 337–348
*To whom correspondence should be sent

each year. A recent study compared leaf breakdown in several sections of the intermittentOued Cherraa (MAAMRI et al., 1997a), which is an extension of the permanently flowingOued Zegzel. The objective of the present study was to compare litter breakdown and itsmain microbial agents in permanent and intermittent reaches of the same stream by intro-ducing leaves simultaneously at four sites along the Zegzel/Cherraa River system and usingthe same methods along with direct counts of bacteria. The experiment was initiated whenthe two Cherraa sites were dry, and extended into the wet period. In this study, we used wil-low leaves (Salix pedicellata) because this species dominates allochthonous inputs into theZegzel/Cherraa system (MAAMRI et al., 1994a and 1997b).
2. Material and Methods
2.1. Sampling Sites (see also MAAMRI et al., 1995 and 1997b)
Oued Zegzel, which is called Oued Cherraa in its lower reaches, is a tributary of the River Moulouyain the north-east of Morocco (Fig. 1). The Zegzel-Cherraa system drains 190 km2 in the mountains and82 km2 on the plains. Oued Zegzel starts with three branches. Even though all of these carry waterthroughout the year, discharge is extremely irregular like in most Mediterranean streams. Water levelsare closely related to rainfall, they are high in winter and spring, and low in summer.
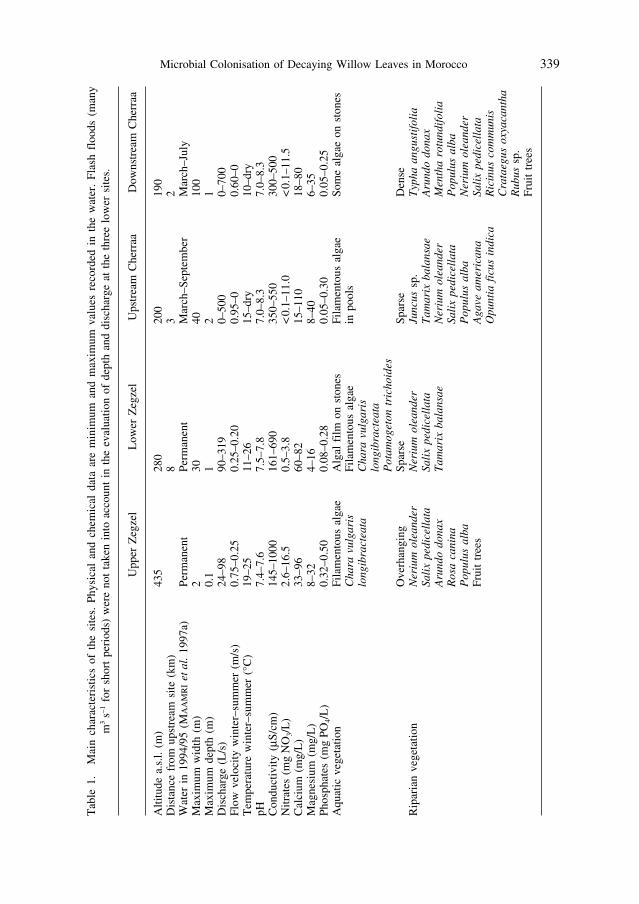
Table 1 provides data on the sites. Two of them were selected on Oued Zegzel:Its South-western Branch (hereafter called Upper Zegzel) includes a succession of short cascades over
large boulders and plant remains, and small sandy pools. Despite the small size of this stream, somewater is removed for irrigation by local inhabitants.
338 A. MAAMRI et al.
Figure 1. Location of the four sampling sites in Oued Zegzel (permanent stream) and Oued Cherraa(intermittent stream).
Microbial Colonisation of Decaying Willow Leaves in Morocco 339
Tab
le 1
.M
ain
char
acte
rist
ics
of t
he s
ites.
Phy
sica
l an
d ch
emic
al d
ata
are
min
imum
and
max
imum
val
ues
reco
rded
in
the
wat
er.
Flas
h fl
oods
(m
any
m3
s–1fo
r sh
ort
peri
ods)
wer
e no
t ta
ken
into
acc
ount
in
the
eval
uatio
n of
dep
th a
nd d
isch
arge
at
the
thre
e lo
wer
site
s.
Upp
er Z
egze
lL
ower
Zeg
zel
Ups
trea
m C
herr
aaD
owns
trea
m C
herr
aa
Alti
tude
a.s
.l. (
m)
435
280
200
190
Dis
tanc
e fr
om u
pstr
eam
site
(km
)8
32
Wat
er i
n 19
94/9
5 (M
AA
MR
Iet
al.
1997
a)Pe
rman
ent
Perm
anen
tM
arch
–Sep
tem
ber
Mar
ch–J
uly
Max
imum
wid
th (
m)
230
4010
0M
axim
um d
epth
(m
)0.
11
21
Dis
char
ge (
L/s
)24
–98
90–3
190–
500
0–70
0Fl
ow v
eloc
ity w
inte
r–su
mm
er (
m/s
)0.
75–0
.25
0.25
–0.2
00.
95–0
0.60
–0T
empe
ratu
re w
inte
r–su
mm
er (
°C)
19–2
511
–26
15–d
ry10
–dry
pH7.
4–7.
67.
5–7.
87.
0–8.
37.
0–8.
3C
ondu
ctiv
ity (
µS/c
m)
145–
1000
161–
690
350–
550
300–
500
Nitr
ates
(m
gN
O3/
L)
2.6–
16.5
0.5–
3.8
<0.
1–11
.0<
0.1–
11.5
Cal
cium
(m
g/L
)33
–96
60–8
215
–110
18–8
0M
agne
sium
(m
g/L
)8–
324–
168–
406–
35Ph
osph
ates
(m
gPO
4/L
)0.
32–0
.50
0.08
–0.2
80.
05–0
.30
0.05
–0.2
5A
quat
ic v
eget
atio
nFi
lam
ento
us a
lgae
Alg
al f
ilm o
n st
ones
Fila
men
tous
alg
aeSo
me
alga
e on
sto
nes
Cha
ra v
ulga
ris
Fila
men
tous
alg
aein
poo
lslo
ngib
ract
eata
Cha
ra v
ulga
ris
long
ibra
ctea
taP
otam
oget
on t
rich
oide
sO
verh
angi
ngSp
arse
Spar
seD
ense
Rip
aria
n ve
geta
tion
Ner
ium
ole
ande
rN
eriu
m o
lean
der
Junc
ussp
.T
ypha
ang
usti
foli
aSa
lix
pedi
cell
ata
Sali
x pe
dice
llat
aT
amar
ix b
alan
sae
Aru
ndo
dona
xA
rund
o do
nax
Tam
arix
bal
ansa
eN
eriu
m o
lean
der
Men
tha
rotu
ndif
olia
Ros
a ca
nina
Sali
x pe
dice
llat
aP
opul
us a
lba
Pop
ulus
alb
aP
opul
us a
lba
Ner
ium
ole
ande
rFr
uit
tree
sA
gave
am
eric
ana
Sali
x pe
dice
llat
aO
punt
ia f
icus
ind
ica
Ric
inus
com
mun
isC
rata
egus
oxy
acan
tha
Rub
us s
p.Fr
uit
tree
s

The site Lower Zegzel was located a short distance downstream from the locality Takarboust. In this section, the valley becomes more open. The water is more exposed to the sun and its temperaturefluctuates more than upstream. Especially during the summer, there is considerable diversion of waterfor agricultural and domestic purposes. The stream bed is dominated by round boulders and cobblescovered with filamentous algae, with large accumulations of sand and mud in between. Coarse particu-late organic matter is less abundant than in the Upper Zegzel (MAAMRI et al., 1996).
Oued Cherraa (the lower reaches of Oued Zegzel; Fig. 1) receives most of its discharge from OuedZegzel. It is an intermittent river with short spates and variable discharge.
The site Upstream Cherraa was located near the village of Tazarine in a valley with steep slopes. Forsome months it is completely dry; any remaining water is diverted by concrete channels (‘seguias’) toirrigate the cultivated terraces of the valley. The river bed consists of large boulders, cobbles and grav-el, with occasional accumulations of sand. Temporary or permanent pools of water are rapidly filledwith filamentous algal growth. The wet season often begins abruptly with a rapid flow of water due tothe first showers in autumn or winter. Its duration is variable and depends primarily on total yearly pre-cipitation. In the year 1994/95, it lasted 7 months (March to September, MAAMRI et al., 1997a). Towardsthe end of the season, the water becomes stagnant and most of it disappears completely.
The site Downstream Cherraa was located at the entrance to the Triffa plain. The wet period is shor-ter, e.g. 5 months in 1994/95 (March to July, MAAMRI et al., 1997a). Even during this period, the varioussections of this site remain separate from each other most of the time. Water circulates from one poolto another through the boulder and cobble substratum without completely covering it. An uninterruptedflow of running water only occurs during floods. In June, very little flowing water remains, and the siteconsists of a collection of essentially stagnant pools, which eventually dry out.
2.2. Processing of Leaves
Naturally-shed leaves of S. pedicellata were collected at the end of October 1998. They were placedfresh in 128 litter bags with a mesh size of 2 mm (5 ± 0.5 g per bag), and were immediately exposedat the four sites on November 1st. Individual bags were attached to riparian vegetation with nylonthreads. The number of replicate bags prepared for determination of weight loss over time was 96(4 sites × 8 collections dates × 3 replicates). The relationship between fresh and dry weight was deter-mined as described in MAAMRI et al., (1997a).
After two, and then after every four weeks, three randomly selected bags were recovered from eachof the four sites. They were placed in plastic bags to avoid the loss of material. In the laboratory, sandand other non-plant material were carefully removed from the leaves with a fine brush. The remainingleaf material was dried for 48 h at 40 °C and weighed to the nearest 0.2 mg.
2.3. Conidia and Bacteria on Leaves
On each sampling occasion, a separate litter bag that had been prepared and exposed as mentionedabove (altogether 32 bags) was collected from each site for microbiological analyses.
In the laboratory, individual leaves from these bags were rinsed, placed in separate 500 ml Erlen-meyer flasks with 300 ml of distilled water, and aerated for 2 d at 15 °C to induce conidium formation(BÄRLOCHER, 1982). The supernatant containing suspended conidia was filtered through an 8 µm mem-brane filter which was then stained with cotton blue in lactophenol for 60 min at 40 °C. The conidiatrapped on the filters were counted and identified using INGOLD (1995), CHAUVET (1990) and MARVA-NOVÁ (1997). The leaves corresponding to the filters were dried (60 °C, 4 d) and weighed, and numbersof conidia per unit weight of substrate were calculated.
Leaves from the same bags were preserved in 37% formaldehyde at –4 °C and transported to Cana-da for counting their bacteria by epifluorescence microscopy. After staining with acridine orange(10 mg L–1) bacterial densities on the upper and the lower surfaces of three discs (1 cm diameter) weredetermined in 20 randomly selected fields of view (Zeiss epifluorescence microscope 46 72 59 9901,100 × objective, filter set 48 77 09). No clumps of detritus or bacteria made counting difficult. Valueswere converted to bacteria per cm2 of upper leaf area. After counting bacteria, the leaf discs were dried(60 °C, 4 d) to determine bacteria per mg dry weight.
340 A. MAAMRI et al.

2.4. Statistics
Daily decay coefficients (k, exponential decay) were estimated by non-linear curve-fitting. Weightloss curves of the four sites were compared by ANCOVA (SYSTAT 5.2 for Macintosh, WILKINSON,1989), and by comparing residual sums of squares (SS) of pooled and separate regressions (MOTUSLKY
and RANSNAS, 1987). Both techniques yielded the same patterns of significance (p ≤ 0.05).
3. Results
The first spate occurred on 3 January 1999, at which time Oued Cherraa began flowing.This was just after the 8-weeks sampling was performed. The first flow in Oued Cherraawas not observed, but the information was gathered from rainfall on the region and confir-med by the local inhabitants.
3.1. Weight Loss
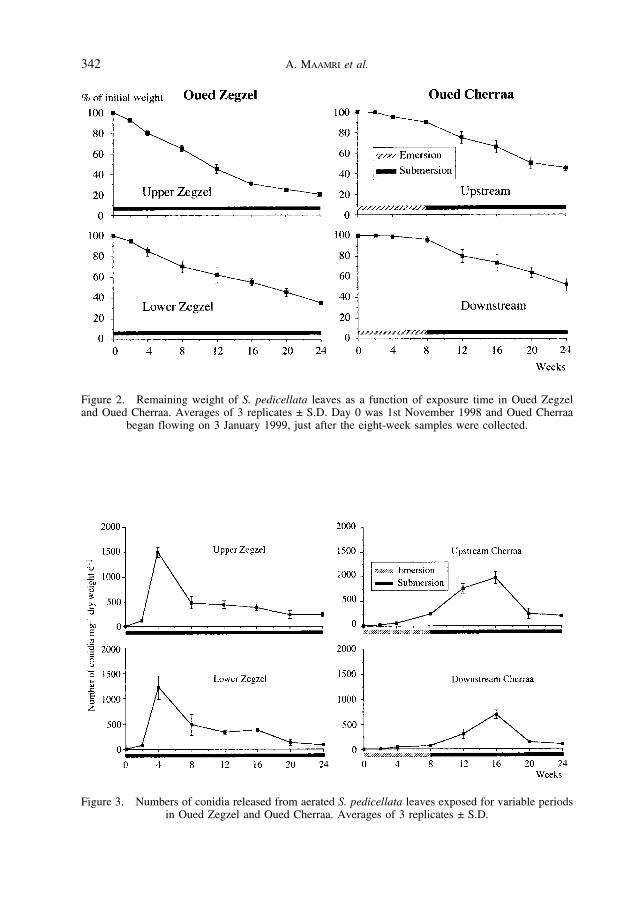
Fig. 2 shows remaining weight of S. pedicellata leaves as a function of exposure time atthe four sites. The exponential rates of decomposition (k, Table 2) varied between 0.0094 d–1
(Upper Zegzel) and 0.0036 d–1 (Downstream Cherraa). Statistical comparisons (ANCOVAand comparisons of residual SS) revealed significant differences (p < 0.05) among the fourdecay curves. The two Zegzel sites also differed significantly from each other, but the twoCherraa sites did not. At these two sites, the arrival of the wet period clearly accelerateddecay: during the first eight weeks (dry), decay rates were 0.0028 (Upstream) and 0.0008(Downstream), respectively; during the remainder of the experiment (wet), they increased to0.0446 and 0.0357, respectively.
3.2. Fungi
Fig. 3 illustrates spore production by aquatic hyphomycetes on leaves recovered from the four sites. Sporulation began earlier, and reached higher levels, at the two permanentZegzel sites (1500 and 1200 mg–1 d–1 at Upper and Lower site, respectively) with peak num-bers recorded after four weeks. By contrast, at the two Cherraa sites, spore production reached maxima after 16 weeks (8 weeks after the onset of the wet period; 900 and700 mg–1 d–1 at Upper and Lower site, respectively). A fully factorial ANOVA revealed highly significant effects of site, length of stream exposure and interaction of the two(p < 0.0001). Tukey’s pairwise comparisons (effects: site) suggested that the sites fall intothree groups: Upper Zegzel, Lower Zegzel and Upstream Cherraa, and Downstream Cher-raa. The same patterns of significance were found when the analyses were done with the ori-ginal and with square-root transformed data.
Microbial Colonisation of Decaying Willow Leaves in Morocco 341
Table 2. Daily weight loss rates (k) of decaying willow leaves, as established by linearcurve-fitting of ln-transformed values.
Stations k r p
Upper Zegzel 0.0094 0.99 0.01Lower Zegzel 0.0056 0.99 0.01Upstream Cherraa 0.0047 0.97 0.05Downstream Cherraa 0.0036 0.96 0.05
342 A. MAAMRI et al.
Figure 2. Remaining weight of S. pedicellata leaves as a function of exposure time in Oued Zegzeland Oued Cherraa. Averages of 3 replicates ± S.D. Day 0 was 1st November 1998 and Oued Cherraa
began flowing on 3 January 1999, just after the eight-week samples were collected.
Figure 3. Numbers of conidia released from aerated S. pedicellata leaves exposed for variable periodsin Oued Zegzel and Oued Cherraa. Averages of 3 replicates ± S.D.

Microbial Colonisation of Decaying Willow Leaves in Morocco 343T
able
3.
Fung
al s
peci
es o
n Sa
lix
pedi
cell
ata
leav
es d
ecay
ing
in O
ued
Zeg
zel
and
Oue
d C
herr
aa.
++
+ p
rese
nt i
n 60
–90
% o
f th
e sa
mpl
es,
++
in
30–6
0%
of
the
sam
ples
, +
in
less
tha
n 30
% o
f th
e sa
mpl
es.
The
ten
spe
cies
mar
ked
with
an
aste
risk
are
men
tione
d fo
r th
e fi
rst
time
in M
oroc
co.
Spec
ies
Oue
d Z
egze
lO
ued
Che
rraa
Upp
er Z
egze
lL
ower
Zeg
zel
Ups
trea
m C
herr
aaD
owns
trea
m C
herr
aa
Act
inos
pora
meg
alos
pora
ING
OL
D+
++
++
Ala
tosp
ora
acum
inat
aIN
GO
LD
++
++
++
++
+A
ngui
llos
pora
fil
ifor
mis
GR
EA
TH
EA
D*
++
+A
ngui
llos
pora
lon
giss
ima
(SA
CC.
et S
YD
.) I
NG
OL
D+
++
++
++
++
++
Ang
uill
ospo
ra c
rass
aIN
GO
LD
*+
+A
ngui
llos
pora
ros
eaD
ESC
AL
Set
MA
RV
AN
OV
A*
++
Cla
vato
spor
a lo
ngib
rach
iata
(IN
GO
LD
) M
AR
VA
NO
VA
et S
. N
ILSO
N+
Cla
vari
opsi
s aq
uati
caD
EW
ILD
.+
++
++
+C
lava
riop
sis
sp.
++
+C
ampo
spor
ium
pel
luci
dum
HA
RK
NE
SS*
+D
acty
lell
a su
bmer
sa(I
NG
OL
D)
S. N
ILSO
N*
++
++
++
++
++
Fla
gell
ospo
ra c
urvu
laIN
GO
LD
++
Fus
ariu
msp
.+
++
++
Gen
icul
ospo
ra i
nfla
ta(I
NG
OL
D)
S. N
ILSS
ON
ex M
AR
VA
NO
VA
et N
ILSS
ON
+H
elis
cell
a st
ella
ta(I
NG
OL
Det
CO
X)
MA
RV
AN
OV
Aet
NIL
SSO
N+
++
Lem
onni
era
aqua
tica
DE
WIL
D.
++
++
++
++
++
Lem
onni
era
terr
esti
sT
UB
AK
I+
++
++
Lun
ulos
pora
cur
vula
ING
OL
D+
++
Myc
ocen
tros
pora
ace
rina
(HA
RT
IG)
DE
IGH
TO
N*
+M
ycoc
entr
ospo
ra c
lava
taI Q
BA
L*
+Sp
orid
esm
ium
ens
irif
orm
eL
INK
*+
++
Tet
racl
adiu
m f
urca
tum
DE
SCA
LS*
+T
etra
clad
ium
mar
chal
ianu
mD
EW
ILD
.+
++
++
+T
ricl
adiu
m s
plen
dens
ING
OL
D+
Tri
clad
ium
ang
ulat
umI N
GO
LD
++
++
+T
ricl
adiu
m g
raci
leIN
GO
LD
*+
++
Tri
posp
erm
umsp
.+
Tri
cell
ula
aqua
tica
WE
BST
ER
++
Unk
now
n (C
lava
tosp
ora
sp.?
)+
++
+
Tot
al n
umbe
r of
spe
cies
2617
1611
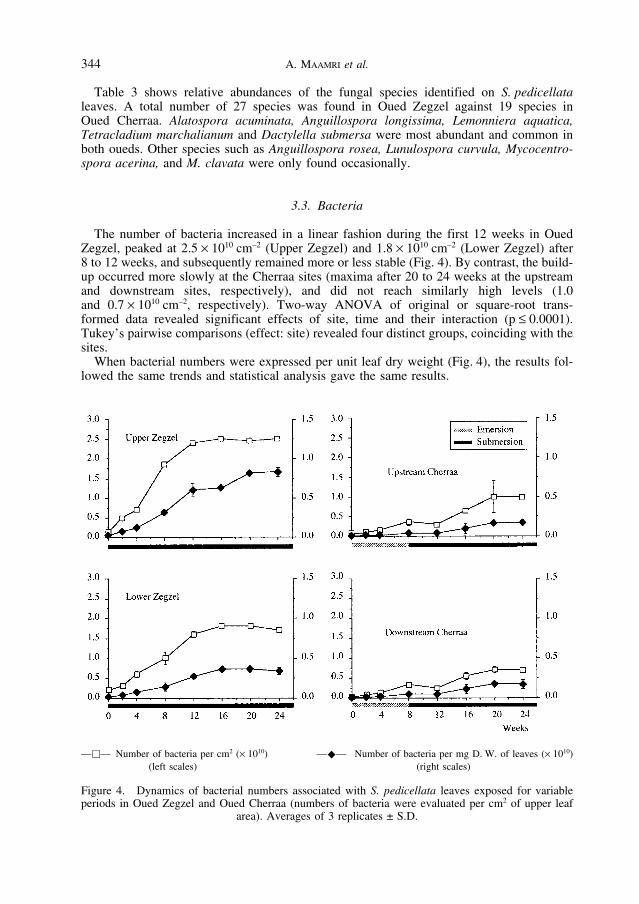
Table 3 shows relative abundances of the fungal species identified on S. pedicellataleaves. A total number of 27 species was found in Oued Zegzel against 19 species in Oued Cherraa. Alatospora acuminata, Anguillospora longissima, Lemonniera aquatica,Tetracladium marchalianum and Dactylella submersa were most abundant and common inboth oueds. Other species such as Anguillospora rosea, Lunulospora curvula, Mycocentro-spora acerina, and M. clavata were only found occasionally.
3.3. Bacteria
The number of bacteria increased in a linear fashion during the first 12 weeks in OuedZegzel, peaked at 2.5 × 1010 cm–2 (Upper Zegzel) and 1.8 × 1010 cm–2 (Lower Zegzel) after8 to 12 weeks, and subsequently remained more or less stable (Fig. 4). By contrast, the build-up occurred more slowly at the Cherraa sites (maxima after 20 to 24 weeks at the upstreamand downstream sites, respectively), and did not reach similarly high levels (1.0 and 0.7 × 1010 cm–2, respectively). Two-way ANOVA of original or square-root trans-formed data revealed significant effects of site, time and their interaction (p ≤ 0.0001).Tukey’s pairwise comparisons (effect: site) revealed four distinct groups, coinciding with thesites.
When bacterial numbers were expressed per unit leaf dry weight (Fig. 4), the results fol-lowed the same trends and statistical analysis gave the same results.
344 A. MAAMRI et al.
Figure 4. Dynamics of bacterial numbers associated with S. pedicellata leaves exposed for variableperiods in Oued Zegzel and Oued Cherraa (numbers of bacteria were evaluated per cm2 of upper leaf
area). Averages of 3 replicates ± S.D.
––�–– Number of bacteria per cm2 (× 1010) ––�–– Number of bacteria per mg D. W. of leaves (× 1010)(left scales) (right scales)

4. Discussion
In an earlier paper, MAAMRI et al. (1997a) showed that leaf decomposition was faster insections of the intermittent Oued Cherraa that retained some water. Similar observationswere made by HILL et al. (1988), and GURTZ and TATE (1988). The present study with wil-low leaves confirms and extends this pattern to the whole watercourse: the overall rate ofdecomposition declined from 0.009 d–1 in the uppermost permanent river section to 0.004 d–1
in the lowest intermittent section. Compared to earlier work by CHERGUI and PATTEE (1993)with the same species in the same stream, we found slightly lower decay rates. However,they used leaf packs compared to 2-mm mesh bags in the present study; the weight loss ofleaves is often somewhat slower in mesh bags (BÄRLOCHER and SCHWEIZER, 1983).
The overall breakdown coefficients of S. pedicellata in Oued Cherraa are among thelowest reported from intermittent rivers. For example, using essentially the same techniques,HILL et al. (1992) found k values between 0.004 and 0.017 d–1 for Carya illinoisensis andAcer negundo; for Ulmus americana, they varied between 0.0074 and 0.0077 d–1 (TATE andGURTZ, 1986). However, breakdown at the temporary sites clearly accelerated with the onsetof the wet period. For the first eight weeks (dry period), k was 0.0028 and 0.0008 (upstreamand downstream, respectively); it increased to 0.0446 and 0.0357 in the wet period, whichexceeded overall rates in the permanent Zegzel sites. This can probably be attributed to thedestruction of membranes and to leaching of antifungal substances from the leaves that haddried on the stream bed for eight weeks (BUNN, 1988; BÄRLOCHER, 1991). The leaves werefresh when first exposed, but dry by the time they were flooded in Oued Cherraa.
Hence breakdown in intermittent rivers clearly depends on the duration of submersion.During air exposure, breakdown in Oued Cherraa was mainly mechanical by cattle tramplingin open locations. Once the water flowed again, one may only speculate whether most dryleaves were carried downstream or whether they decayed on the spot (GASITH and RESH,1999). Unfortunately the first flow was not observed, but the retention of dry leaves may beestimated from former work at the Lower Zegzel site. This site is close to Upstream Cher-raa and its substratum (boulders, cobble and gravel) is similar to that of both Cherraa sites.The mean distance travelled by dry leaves of S. pedicellata and N. oleander thrown into theriver at the Lower Zegzel site varied between 23 and 333 m according to season and hencedischarge (MAAMRI et al., 1994b). In winter, extrapolation indicated that 99 % of the leaveswere trapped within 1150 m (N. oleander) or 1540 m (S. pedicellata). As Oued Cherraa isapproximately 18-km long, most leaf input seems to be processed in this river.
From their experiments in the Garonne River, GESSNER and CHAUVET (1994) concludedthat the rate of leaf decomposition was controlled by microscopic fungi, or, more specifi-cally, by aquatic hyphomycetes. In the present study, differences in the decay rates ofS. pedicellata at the four sites ran parallel to the numbers of fungal species: these numbersdecreased from a peak of 26 at Upper Zegzel to a low of 11 at Downstream Cherraa. Onthe other hand, RAVIRAJA et al. (1998) found no significantly slower leaf decomposition inan Indian stream, when organic pollution reduced the number of aquatic hyphomycete spe-cies by over 80%. This observation suggests that the ecological functions of many of thesespecies overlap to a considerable extent, or that they may be replaced by terrestrial fungi orbacteria. Indeed, ZEMEK et al. (1985) demonstrated that most of the 35 aquatic hyphomyce-te species that they screened degraded polysaccharides as well as other major constituentsof leaf litter.
Rather than with number of species, the rate of breakdown seems to be correlated withmaximum fungal biomass or with maximum fungal spore production (GESSNER and CHAUVET,1994; MAHARNING and BÄRLOCHER, 1996). The same relationship applies in the present studybetween log(conidia) and the rate of breakdown with a correlation coefficient of 0.93(p < 0.05). Conidium production per µg litter declined from 1.5 to 0.7 (Upper Zegzel toDownstream Cherraa), which falls between the values reported from a French stream
Microbial Colonisation of Decaying Willow Leaves in Morocco 345

(GESSNER and CHAUVET, 1994) and those from a Canadian stream (MAHARNING and BÄR-LOCHER, 1996).
Not surprisingly, the dynamics of conidium production differed among the four sites. Itpeaked after four weeks of immersion at the two permanent sites, which agrees with severalother studies (e.g., BÄRLOCHER, 1982; GESSNER and CHAUVET, 1994; BALDY et al., 1995). Atthe intermittent Oued Cherraa sites, maxima occurred eight weeks after the start of the wet period, or 16 weeks after placing the leaves in the intermittent river. However, someconidia were released after 2 and 4 weeks (Upstream and Downstream Cherraa, respectively).Apparently, aquatic hyphomycetes were able to survive on or in the dry stream bed and colo-nise the leaves. This survival may have been facilitated by the relatively high air humidity(between 70 and 82% in 1994/95, MAAMRI et al., 1997a) and the light showers that pre-ceded the heavy rain in January. Lower night temperatures may also have induced dew for-mation, and the presence of a film of water can be instrumental in lateral dispersal of aqua-tic hyphomycete conidia (BANDONI and KOSKE, 1974). Nevertheless, fungal colonisationclearly increased with the onset of permanent water flow. This increase is not surprisingbecause in addition to providing more favourable conditions for the growth of aquatichyphomycetes, spore production is also stimulated by water current, which will increase the likelihood of successful colonisation (SANDERS and WEBSTER, 1980; BÄRLOCHER, 1991;MAAMRI et al., 1998).
All aquatic hyphomycetes identified during the study are listed in Table 3. More specieswere found in the permanent sections; nevertheless, two species (Clavatospora longibra-chiata and Geniculospora inflata) were restricted to Oued Cherraa. Ten species are reportedfor the first time from the Zegzel/Cherraa system.
In addition to fungi, bacteria often play an important role in litter decomposition (e.g. TAM
et al., 1983; MAAMRI et al., 1999). Like fungal species richness and spores, bacterial num-bers were significantly higher on leaves exposed in the permanent Zegzel sites (p < 0.001).We found considerably higher numbers than MAAMRI et al. (1999) on the same leaf species.This difference could be due to differences in counting technique, and/or in nutrient or dis-solved organic matter concentrations in the water. MAAMRI (in press) found bacterial num-bers between 107 (sonication) and 1010 (grinding) on the same batches of willow and oleander leaves according to the technique used. Like SRIDHAR and BÄRLOCHER (1993), wecounted bacteria directly on the leaf while other authors detached the bacteria from the leaves through ultrasonication (VELJI and ALBRIGHT, 1986; NEWELL et al., 1996; MAAMRI
et al., 1999), with a stomacher (FRY et al., 1985), or by grinding (BALDY et al., 1995). Counting the bacteria directly on decaying leaves eliminates potential errors due to incomplete detachment, cell breakage or the difficulty of identifying bacteria among leafdebris.
5. Conclusion
Overall breakdown of S. pedicellata leaves proceeded faster at the permanently flowingOued Zegzel sites than at the intermittent Oued Cherraa sites. The difference can be attributed primarily to the very slow breakdown during air exposure. But breakdown wasalso faster in the Upper than in the Lower Zegzel (both permanent sites). The same gradientfrom highest (Upper Zegzel) to lowest (Downstream Cherraa) values was found in speciesrichness of aquatic hyphomycetes, their spore production from recovered leaves and thenumber of bacteria present on decaying leaves. Nevertheless, breakdown rate and fungalsporulation greatly increased in intermittent Oued Cherraa after flooding, suggesting thatsome inconspicuous preparation had taken place during the dry period. Surprisingly, somecolonisation by aquatic hyphomycetes occurred even during this period.
346 A. MAAMRI et al.

6. Acknowledgements
This project was made possible by financial support from Societas Internationalis Limnologiae.
7. References
BALDY, V., M. O. GESSNER, and E. CHAUVET, 1995: Bacteria, fungi and the breakdown of leaf litter ina large river. – Oikos 74: 93–102.
BANDONI, R. J. and R. E. KOSKE, 1974: Monolayers and microbial dispersal. – Science 183: 1079–1081.BÄRLOCHER, F., 1982: Conidium production from leaves and needles in four streams. – Canadian J. Bot.
60: 1487–1494.BÄRLOCHER, F., 1991: Fungal colonisation of fresh and dried leaves in the river Teign. – Nova Hedwi-
gia 52: 349–357.BÄRLOCHER, F. and M. SCHWEIZER, 1983: Effects of leaf size and decay rate on colonisation by aquatic
hyphomycetes. – Oikos 41: 205–210.BUNN, S. E., 1988: Processing of leaf litter in a northern jarrah forest stream, Western Australia: I. Sea-
sonal differences. – Hydrobiologia 162: 201–210.CHAUVET, E., 1990: Hyphomycètes aquatiques du sud-est de la France. – Gaussenia 6: 3–31.CHERGUI, H. and E. PATTEE, 1992: Processing of fresh and dry Salix leaves in a Moroccan river system.
– Acta oecol. 13: 291–298.CHERGUI, H. and E. PATTEE, 1993: Fungal and invertebrate colonization of Salix fresh and dry leaves
in a Moroccan river system. – Arch. Hydrobiol. 127: 57–72.FINDLAY, S. E. G. and T. L. ARSUFFI, 1989: Microbial growth and detritus transformation during decom-
position of leaf litter in a stream. – Freshwat. Biol. 21: 261–269.FRY, J. C., R. GOULDER, and C. A. RIMES, 1985: A note on the efficiency of stomaching for the quanti-
tative removal of epiphytic bacteria from submerged aquatic plants. – J. appl. Bacteriol. 58: 113–115.GASITH, A. and V. H. RESH, 1999: Streams in Mediterranean climate regions: abiotic influences and bio-
tic responses to predictable seasonal events. – Annu. Ecol. Rev. Syst. 30: 51–81.GESSNER, M. O. and E. CHAUVET, 1994: Importance of stream microfungi in controlling breakdown rates
of leaf litter. – Ecology 75: 1807–1817.GURTZ, M. E. and C. M. TATE, 1988: Hydrologic influences on leaf decomposition in a channel and
adjacent bank of a gallery forest stream. – Amer. Midl. Nat. 120: 11–21.HILL, B. H., T. J. GARDNER, and O. F. EKISOLA, 1988: Breakdown of gallery forest leaf litter in inter-
mittent and perennial prairie streams. – Southw. Nat. 33: 323–331.HILL, B. H., T. J. GARDNER, O. F. EKISOLA, and G. M. HENEBRY, 1992: Microbial use of leaf litter in
prairie streams. – J. N-Amer. Benthol. Soc. 11: 11–19.INGOLD, C. T., 1975: Guide to aquatic hyphomycetes. – Freshwater Biological Association, Sci. Publ.
N° 30. Ambleside, Cumbria, England, 96 p.MAAMRI, A., H. CHERGUI, and E. PATTEE, 1994a: Allochthonous input of coarse organic matter to a
Moroccan mountain stream. – Acta oecol. 15: 495–508.MAAMRI, A., H. CHERGUI, and E. PATTEE, 1994b: La rétention des feuilles mortes dans un cours d’eau
de montagne au Maroc: variations saisonnières. – Annls Limnol. 30: 323–332.MAAMRI, A., H. CHERGUI, and E. PATTEE, 1995: Impact des apports végétaux allochtones sur la distri-
bution de la faune benthique dans un cours d’eau montagnard marocain, l’Oued Zegzel. – Ecologie26: 81–94.
MAAMRI, A., H. CHERGUI, and E. PATTEE, 1996: Dynamics of fine particulate and dissolved organic mat-ter in a Moroccan mountain stream: effects of stream flow and litter quality. – Pol. Archwm Hydro-biol. 43: 401–410.
MAAMRI, A., H. CHERGUI, and E. PATTEE, 1997a: Leaf litter processing in a temporary NortheasternMoroccan river. – Arch. Hydrobiol. 140: 513–531.
MAAMRI, A., H. CHERGUI, and E. PATTEE, 1997b: Dynamique des apports de litière végétale et de lafaune invertébrée dans une rivière méditerranéenne temporaire: l’Oued Cherraa, au Maroc. – Ecolo-gie 28: 251–264.
MAAMRI, A., E. CHAUVET, H. CHERGUI, F. GOURBIERE, and E. PATTEE, 1998: Microbial dynamics ondead leaves in a temporary Moroccan river. I-Fungi. – Arch. Hydrobiol. 144: 41–59.
MAAMRI, A., E. PATTEE, X. GAYTE, and H. CHERGUI, 1999: Microbial dynamics on dead leaves in atemporary Moroccan river. II-Bacteria. – Arch. Hydrobiol. 144: 157–175.
Microbial Colonisation of Decaying Willow Leaves in Morocco 347

MAAMRI, A., in press: A critical comparison of techniques for estimating bacterial abundance on leaflitter. – Annls Limnol.
MAHARNING, A. R. and F. BÄRLOCHER, 1996: Growth and reproduction in aquatic hyphomycetes. –Mycologia 88: 80–88.
MARVANOVÁ, L., 1997: Freshwater hyphomycetes: a survey with remarks on tropical taxa. – In: JANARD-HANAN, K. K., C. RAJENDRAN, K. NATARAJAN and D. L. HAWKSWORTH (eds). Tropical Mycology.Science Publishers, U.S.A.: 169–226.
MOTULSKY, H. J. and L. A. RANSNAS, 1987: Fitting curves to data using nonlinear regression: a prac-tical and nonmathematical review. – FASEB Journal 1: 365–374.
NEWELL, S. Y., T. L. ARSUFFI, and L. A. PALM, 1996: Misting and nitrogen fertilization of shoots of asaltmarsh grass: effects upon fungal decay of leaf blades. – Oecologia 108: 495–502.
NEWELL, S. Y., R. D. FALLON, and J. D. MILLER, 1989: Decomposition and microbial dynamics for stand-ing, naturally positioned leaves of the salt-marsh grass Spartina alterniflora. – Marine Biology (Berlin)101: 471–481.
RAVIRAJA, N. S., K. R. SRIDHAR, and F. BÄRLOCHER, 1998: Breakdown of Ficus and Eucalyptus leavesin an organically polluted river in India: fungal diversity and ecological functions. – Freshwat. Biol.39: 537–545.
SANDERS, P. F. and J. WEBSTER, 1980: Sporulation responses of some aquatic hyphomycetes in flowingwater. – Trans. Br. Mycol. Soc. 74: 601–605.
SRIDHAR, K. R. and F. BÄRLOCHER, 1993: Seasonal changes in microbial colonisation of fresh and driedleaves. – Arch. Hydrobiol. 128: 1–12.
TAM, T. Y., C. I. MAYFIEL, and W. E. INNISS, 1983: Microbial decomposition of leaf material at 0 °C. –Microb. Ecol. 9: 355–362.
TATE, C. M. and M. E. GURTZ, 1986: Comparison of mass loss, nutrients, and invertebrates associatedwith elm leaf litter decomposition in perennial and intermittent reaches of tallgrass prairie streams. –Southw. Nat. 31: 511–520.
VELJI, M. I. and L. J. ALBRIGHT, 1986: Microscopic enumeration of attached marine bacteria of seawater,marine sediment, fecal matter and kelp blade samples following pyrophosphate and ultrasound treat-ments. – Can. J. Microbiol. 32: 121–126.
WILKINSON, L., 1989: SYSTAT: the system for statistics. – SYSTAT Inc., Evanston, IL., USA.ZEMEK, J., L. MARVANOVÁ, L. KUNIAK, and B. KADLECIKOVA, 1985: Hydrolytic enzymes in aquatic
hyphomycetes. – Folia Microbiol. 30: 363–372.
Manuscript received April 20th, 2000; revised October 2nd, 2000; accepted November 25th, 2000
348 A. MAAMRI et al.





























