Embed Size (px)
Citation preview

Infection, Genetics and Evolution 4 (2004) 353–359
Geographic variation of metric properties within the neotropical sandflies
Jean-Pierre Dujardina,∗, François Le Pontba Institut de Recherche pour le Développement (IRD), Génétique et Evolution des Maladies Infectieuses (GEMI),
UMR 2724 CNRS/IRD, Avenue Agropolis 911, BP 64501, 34394 Montpellier, Franceb IRD La Paz, CP 9214 La Paz, Bolivia
Received 18 April 2004; received in revised form 30 April 2004; accepted 3 May 2004Available online 20 July 2004
Abstract
Traditional morphometrics remains a useful tool for differentiating sandflies species, particularly closely related taxa. Within a singlespecies, however, size variation among geographic populations might interfere with species distinction. In the past decades, a search for ametric property less prone to individual variations led to the use of ratios, and more recently to the use of size-free variables (after tentativesize extraction). While morphometrics is continuously enriched with new, more efficient techniques describing shape, the question remainsfor the morphologist whether an efficient removing of within-species size variation is sufficient to produce stable, specific characters.Is a single species metrically recognizable in spite of its geographical variation? To address this question the intraspecific variationin 10 neotropical sandflies was examined within and between large South American ecogeographic regions: the Amazonian region, theSub-Andean and the Chaco regions of Bolivia, and the Atlantic Coast of Brazil. Thus, the geographic stability of metric properties, as derivedfrom measurements between landmarks, was investigated 10 times allowing a total of 29 inter-regional and 13 intra-regional conspecificcomparisons. Metric proportions remained stable among conspecific populations of a single ecogeographic region. From one region toanother, however, size-independent changes were significant and their amount was correlated with altitude. This could reflect an importantrole of some environmental changes in shaping the insects, and should be taken into account when using morphometrics to identify sandflies.© 2004 Elsevier B.V. All rights reserved.
Keywords: Allometry; Altitude; Epidemiology; Medical entomology; South America; Taxonomy
1. Introduction
It is a current finding to observe among populations ofthe same sandfly species significant morphological (Youngand Duncan, 1994), genetic (Bonnefoy et al., 1986; Dujardinet al., 1997) or metric variations (Dujardin et al., 1997),some of them even questioning its taxonomic status (Caillardet al., 1986; Lanzaro et al., 1993; Dujardin et al., 1996).As sandflies have medical importance, particularly as leish-maniasis vectors, their correct taxonomic identification is acrucial component of any epidemiological study (Lanzaroand Warburg, 1995).
To understand the taxonomic significance of morpholog-ical variation in sandflies, genetic techniques are currentlyapplied (Kreutzer et al., 1990; Lanzaro et al., 1993; Dujardinet al., 1993, 1996). Morphometrics may be of valuable helptoo (Lane, 1988; Gebre-Michael and Lane, 1993), particu-larly when scaling for size variation (Dujardin and Le Pont,2000; Dujardin et al., 1999) or using geometric morphom-
∗ Corresponding author. Tel.:+33-4-67416237; fax:+33-4-67416299.E-mail address: [email protected] (J.-P. Dujardin).
etry (Dujardin et al., 2002). The main purpose of this studywas to evaluate the stability of traditional metric traits in theneotropical sandfly species (Lane and Ready, 1985; Lebbeet al., 1991; Añez et al., 1997; Gebre-Michael andMedhin, 1997; Marcondes et al., 1998), by testing on truedata the tenet of size-independent traits as a more sta-ble characters than size. The partition of metric variationinto “size” and “size-independent” variables is intended toprovide independent indicators of the way an organism’smorphology interacts with its immediate environment. Sizevariation is thought as having more environmental vari-ance than allometry-free differences, which refer to shapechanges being not merely a passive consequence of sizevariation. If this proposition is true, and as long as we con-sider a single species, the metric variation disclosed withgeography would be attributable to size differences due tosome growth inequality, and shape variation would only beits direct allometric effects. We explored here this questionby testing the null hypothesis of conspecific populationsbeing simply the allometric extension each one another(Klingenberg, 1996), comparing the geographic stability ofmetric properties within 10 species of neotropical sandflies.
1567-1348/$ – see front matter © 2004 Elsevier B.V. All rights reserved.doi:10.1016/j.meegid.2004.05.001

354 J.-P. Dujardin, F. Le Pont / Infection, Genetics and Evolution 4 (2004) 353–359
2. Materials and methods
2.1. Insects (Table 1)
Samples of the following ten New World species wereexamined: in the subgenusPsychodopygus: Lutzomyia ay-rozai (Barretto and Coutinho),L. geniculata (Mangabeira),L. hirsuta (Mangabeira),L. carrerai (Barretto); in the speciesgroupMigonei: L. cortelezzii (Brethes),L. lentioides (sensuForattini, 1971); in the subgenusNyssomyia: L. umbratilis(Ward and Fraha),L. whitmani (Antunes and Coutinho); inthe subgenusPsathyromyia: L. shannoni and in the sub-genusSciopemyia: L. sordelli (Table 1). These are quitecommon and widespread (except forL. cortelezzii) neotrop-ical species, four of them recognized as leishmaniasis vec-tors: L. ayrozai, L. carrerai, L. umbratilis andL. whitmani.Insects were male specimens except forL. carrerai andL.umbratilis. Species identification was performed on mor-phological criteria following the keys of Young and Duncan(1994).
For each species, three to four geographic populationswere selected from at least two of the following regions: theAmazon basin (mainly French Guyana and North Beni orPando in Bolivia), the Rio de la Plata basin (mainly BolivianChaco), the Sub-Andean region from Bolivia (Yungas, AltoBeni and Alto Pilcomayo) and the Atlantic coast of Brazil(Sao Paulo) (seeFig. 1).
2.2. Characters
The following non-redundant structures (i.e. one measure-ment is not included in another one) were considered forpossible measurement: the third antennal segment (AIII),the labrum-epipharynx (EP), the height (HH), the interocu-lar distance (ID) or the third palpomere (P3) for the head;the width (WW), the length (WL), the “gamma” (GA) or the“alpha” (AL) dimensions for the wing; the genital filament(GF), the genital pump (GP), the lateral lobe (LL) or thestyle length (ST) for the genitalia. With one exception (L.hirsuta) at least one dimension was measured of each of thethree anatomical parts (head, wing, genitalia). In the two fe-male comparisons (L. carrerai, L. umbratilis) the measure-ments were limited to head and wings since female genitaliado not offer reliable landmarks between which to measuredistances. We used a maximum of seven characters, so thatthe ratio characters/sample within each group was alwaysless than 50% (Bookstein, 1996). Thus each species was de-fined by three to seven variables, while sample size was onaverage 14.5.
2.3. Numerical methods
Modern statistical techniques in morphometrics allow totest the null hypothesis of conspecific populations being sim-ply the allometric extension each one another, provided a
Fig. 1. Maps describing the ecogeographic areas of Latin America. Map1 (top) shows the northern and central parts of South America; map 2(bottom) shows Bolivia. Ecogeographic areas are: the Amazonian basin(see map 1, Amazonia, MOMELI; and map 2 RIB, RURTUM), theSub-Andean region (see map 2 of Bolivia, PAL, SUA, PAR, CAJ, UYU),the Chaco (CHO, SAN, ROB, MAR) and Atlantic coast (see map 2, SAOin the Atlantic forest of Brazil). CHO, Chochis (Chaco, Bolivia); MAR,Margarita (Bolivia); MONELI, Monsinery and St-Elie (Guyana); PAL,Palos Blancos (Bolivia); RIB, Riberalta (Bolivia); ROB, Robore (Bo-livia); RUR TUM, Rurrenabaque and Tumupasa (Bolivia); SAN, SantoCorazon (Bolivia); SAO, Sao Paulo (Brazil); SUAPAR CAJ, Suapi,Pararani and Cajuata, three Bolivian localities; UYU, Uyuni (Bolivia); km,kilometer.
common allometric trend is identifiable (for a review, seeKlingenberg, 1996). To remove a common allometric trendamong a set of measurements, we used the common prin-cipal components model (Airoldi and Flury, 1988). In thismodel, a single component is taken to account for shared al-lometric variation. In the space of the log-transformed mea-surements, conspecific individuals are expected to be foundalong the straight line defined by this single component. Met-
J.-P. Dujardin, F. Le Pont / Infection, Genetics and Evolution 4 (2004) 353–359 355
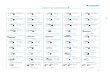
Table 1Insects
Year Loc Latitude Longitude ay ge hi ca co le um wh sh so
Sub-Andean region (Bolivia)1987 Tum – – – – – – 13 – – –1987 Rur 14◦71′S 67◦35′W – – – 15 – – – – – –1987 Sua 16◦18′S 67◦45′W 25 10 – – – 15 – – – –1987 Par 16◦18′S 67◦37′W – – – – – – – – 15 –1996 Caj 16◦43′S 67◦12′W – – – – – 15 – – 15 –1986 Pal 15◦27′S 67◦34′W – – 11 15 – – – – – –1987 Uyu 19◦15′S 64◦57′W – – – – 15 – – – – –
Chaco region (Bolivia)1987 Mar 21◦11′S 63◦42′W – – – – 15 – – – – –2000 Rob 21◦34′S 62◦59′W – – – – 15 – – – – –2000 San 17◦59′S 58◦50′W – – – – – 15 – 15 – 152000 Cho 17◦47′S 60◦25′W – – – – – – – 13 – –
Amazonian Basin (Bolivia, French Guyana)1988 Rib 11◦00′S 66◦05′W 27 10 8 15 – – 15 21 6 151979 Mon 4◦54′N 52◦30′W 23 7 11 – – – 15 – 15 151979 Eli 5◦12′N 53◦02′W – – 12 – – – – – – –
Atlantic coast region (Brazil)1985 Sao 23◦32′S 46◦38′W 11 10 – – – – – – – –
Total (488 insects) 86 37 42 45 45 45 43 49 51 45
Sample sizes of the different groups analyzed species by species. For each species (columns “ay” to “so”), the year of their capture (first column), thelocality of capture, its latitude and its longitude. The last row contains the sum of specimens by species (the grand total is 488). ay,ayrozai; ge,geniculata;hi, hirsuta; ca, carrerai (females); co,cortelezzii; le, lentioides; um, umbratilis (females); wh,whitmani; sh, shannoni; so, sordelli; Loc, localities; Caj,Cajuata; Cho, Chochis; Mar, Margarita; Pal, Palos Blancos; Par, Pararani; Rib, Riberalta (North Beni, Bolivia); Rob, Robore; Rur, Rurrenabaque; San,Santo Corazon; Sua, Suapi; Tum, Tumupasa; Uyu, Uyuni; Mon, Monsinery (French Guyana); Eli, Piste de St Elie (French Guyana); Sao, Sao Paulo(Brazil); S, south; W, west.
ric variation orthogonal to this direction is allometry-freeby construction, describing metric differences independentfrom size variation.
Using various sets of measurements described above, wefirst checked the hypothesis of a common allometric axis(i.e. the model of a common principal component). It wasfound in all cases, as expected for conspecific populations,but not for the same sets of characters (since we dealt withdifferent species). The influence of within-group allome-tries was then removed by using variables equivalent to anorthogonal projection of the data onto the first commonprincipal component (i.e. all the common principal compo-nents except the first one). The resulting “allometry-free”,or size-free, variables were submitted to a canonical vari-ate analysis (CVA), and Mahalanobis distances were derived(Mahalanobis, 1936).
2.4. Statistics on size-free and size-in variation
Size-free (or “allometry-free”) divergence was estimatedby the Mahalanobis distances derived from the various com-mon principal components except the first one. Size varia-tion was based on one variable only, i.e. the first commonprincipal component, so that we used the univariate equiva-lent of Mahalanobis distance known as the Pearson’s “Co-efficient of Racial Likeliness” (Nei, 1987; Pearson, 1926),to estimate its variation.
The independence of size-free and size-in distances wasexplored by a simple linear regression test between thecorresponding estimations. This relationship was examinedthree times: (i) on the sample of within-regions compar-isons, (ii) on the sample of between-regions comparisonsand (iii) on the total sample. The statistical significance ofrelationships of these distances with either geographic sep-aration or altitudinal variation was explored by parametricand non-parametric (Mantel) tests.
3. Results
We performed a total of 29 inter-regional and 13intra-regional comparisons (Fig. 1) on a total of 488specimens (Table 1). In the within-region and in thebetween-regions comparisons, as well as in the total sam-ple, no significant correlation could be found between sizeand size-free variation (detailed results not shown). Thissuggested we could indeed construct acceptable size-freevariables, statistically independent of size.
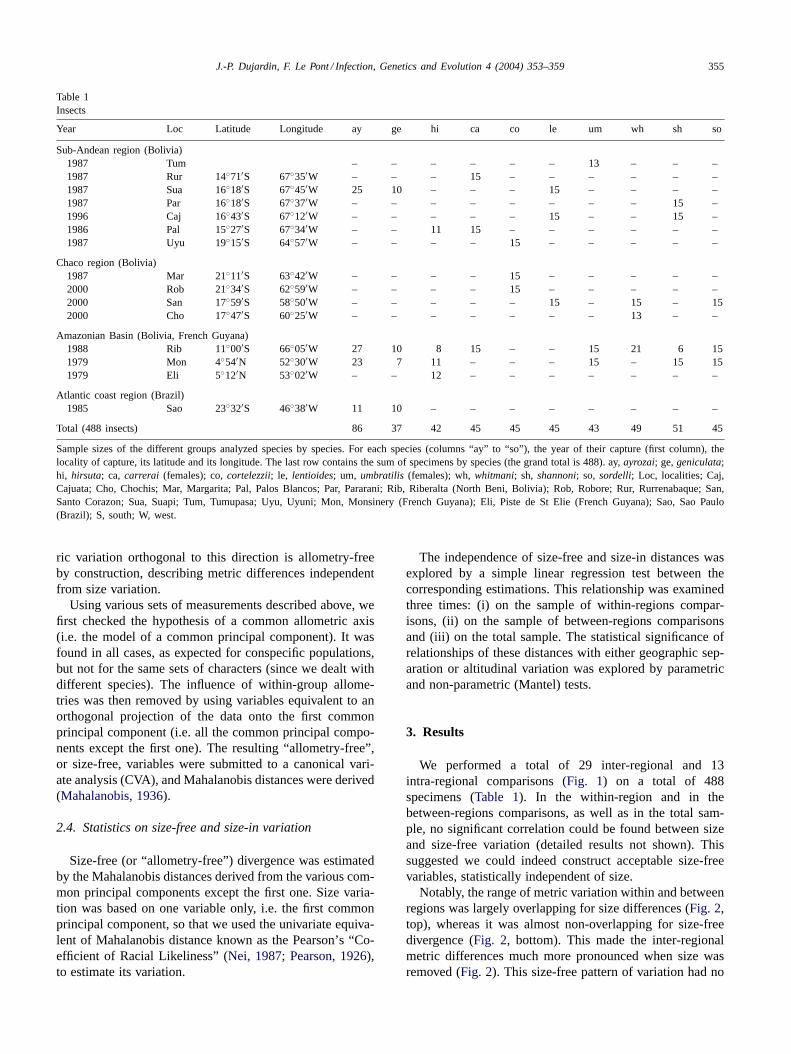
Notably, the range of metric variation within and betweenregions was largely overlapping for size differences (Fig. 2,top), whereas it was almost non-overlapping for size-freedivergence (Fig. 2, bottom). This made the inter-regionalmetric differences much more pronounced when size wasremoved (Fig. 2). This size-free pattern of variation had no
356 J.-P. Dujardin, F. Le Pont / Infection, Genetics and Evolution 4 (2004) 353–359
2.59 81.72Within
Between
Allometry-free variation
40.290.01
Between
Within
Size variation
Fig. 2. Quantile plots showing range of variation in size (top) orsize-independent (bottom) divergences between two populations, accord-ing to their belonging to the same eco-geographic region (W for withinregion) or to distinct ones (B for between regions). Allometry-free diver-gence is estimated by the Mahalanobis distance derived from discrimi-nant analysis on shape. Size variation is estimated by the CRL index ofPearson (1926)using the values of the first common principal component.Each box shows the group median as a vertical line across the middleand the quartiles (25th and 75th percentiles) as its ends. The 10th and90th quantiles are shown as lines on each sides the box.
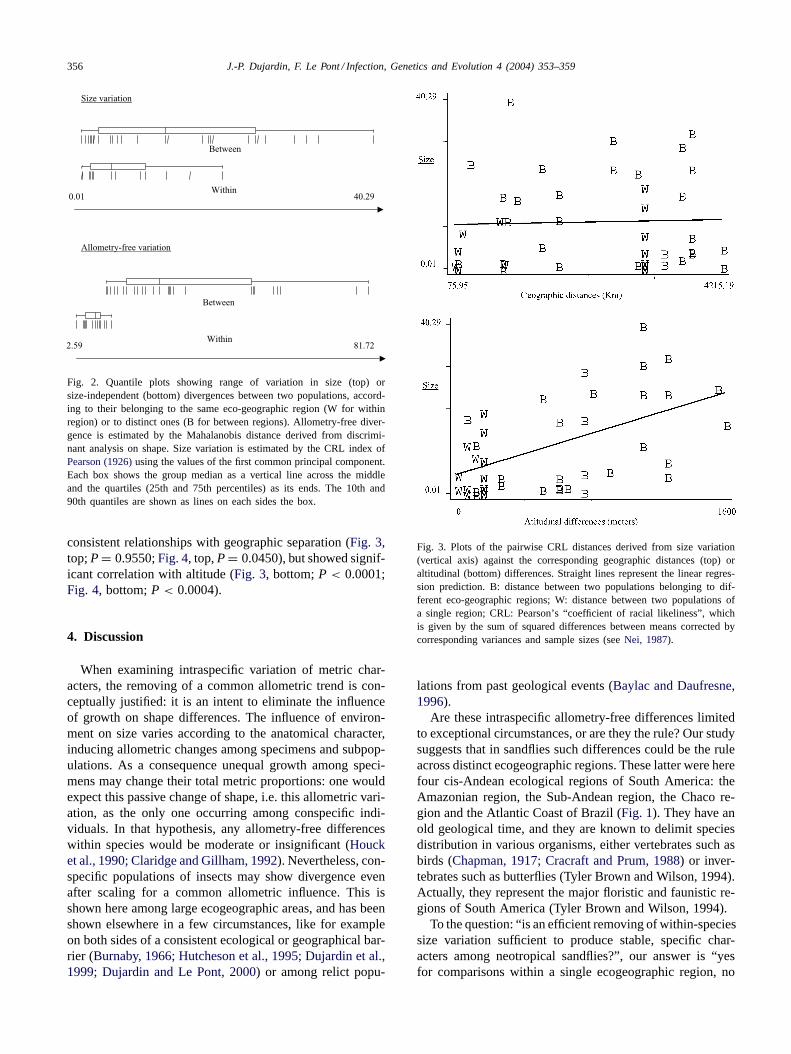
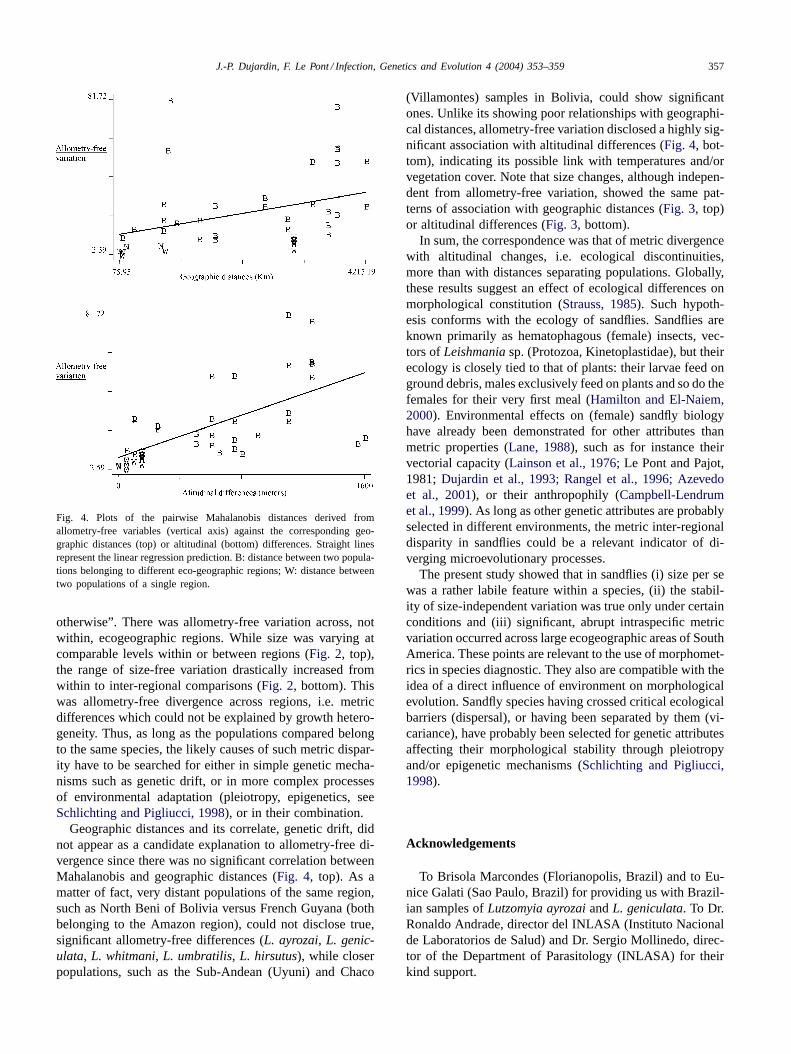
consistent relationships with geographic separation (Fig. 3,top;P = 0.9550;Fig. 4, top,P = 0.0450), but showed signif-icant correlation with altitude (Fig. 3, bottom;P < 0.0001;Fig. 4, bottom;P < 0.0004).
4. Discussion
When examining intraspecific variation of metric char-acters, the removing of a common allometric trend is con-ceptually justified: it is an intent to eliminate the influenceof growth on shape differences. The influence of environ-ment on size varies according to the anatomical character,inducing allometric changes among specimens and subpop-ulations. As a consequence unequal growth among speci-mens may change their total metric proportions: one wouldexpect this passive change of shape, i.e. this allometric vari-ation, as the only one occurring among conspecific indi-viduals. In that hypothesis, any allometry-free differenceswithin species would be moderate or insignificant (Houcket al., 1990; Claridge and Gillham, 1992). Nevertheless, con-specific populations of insects may show divergence evenafter scaling for a common allometric influence. This isshown here among large ecogeographic areas, and has beenshown elsewhere in a few circumstances, like for exampleon both sides of a consistent ecological or geographical bar-rier (Burnaby, 1966; Hutcheson et al., 1995; Dujardin et al.,1999; Dujardin and Le Pont, 2000) or among relict popu-
Fig. 3. Plots of the pairwise CRL distances derived from size variation(vertical axis) against the corresponding geographic distances (top) oraltitudinal (bottom) differences. Straight lines represent the linear regres-sion prediction. B: distance between two populations belonging to dif-ferent eco-geographic regions; W: distance between two populations ofa single region; CRL: Pearson’s “coefficient of racial likeliness”, whichis given by the sum of squared differences between means corrected bycorresponding variances and sample sizes (seeNei, 1987).
lations from past geological events (Baylac and Daufresne,1996).
Are these intraspecific allometry-free differences limitedto exceptional circumstances, or are they the rule? Our studysuggests that in sandflies such differences could be the ruleacross distinct ecogeographic regions. These latter were herefour cis-Andean ecological regions of South America: theAmazonian region, the Sub-Andean region, the Chaco re-gion and the Atlantic Coast of Brazil (Fig. 1). They have anold geological time, and they are known to delimit speciesdistribution in various organisms, either vertebrates such asbirds (Chapman, 1917; Cracraft and Prum, 1988) or inver-tebrates such as butterflies (Tyler Brown and Wilson, 1994).Actually, they represent the major floristic and faunistic re-gions of South America (Tyler Brown and Wilson, 1994).
To the question: “is an efficient removing of within-speciessize variation sufficient to produce stable, specific char-acters among neotropical sandflies?”, our answer is “yesfor comparisons within a single ecogeographic region, no
J.-P. Dujardin, F. Le Pont / Infection, Genetics and Evolution 4 (2004) 353–359 357
Fig. 4. Plots of the pairwise Mahalanobis distances derived fromallometry-free variables (vertical axis) against the corresponding geo-graphic distances (top) or altitudinal (bottom) differences. Straight linesrepresent the linear regression prediction. B: distance between two popula-tions belonging to different eco-geographic regions; W: distance betweentwo populations of a single region.
otherwise”. There was allometry-free variation across, notwithin, ecogeographic regions. While size was varying atcomparable levels within or between regions (Fig. 2, top),the range of size-free variation drastically increased fromwithin to inter-regional comparisons (Fig. 2, bottom). Thiswas allometry-free divergence across regions, i.e. metricdifferences which could not be explained by growth hetero-geneity. Thus, as long as the populations compared belongto the same species, the likely causes of such metric dispar-ity have to be searched for either in simple genetic mecha-nisms such as genetic drift, or in more complex processesof environmental adaptation (pleiotropy, epigenetics, seeSchlichting and Pigliucci, 1998), or in their combination.
Geographic distances and its correlate, genetic drift, didnot appear as a candidate explanation to allometry-free di-vergence since there was no significant correlation betweenMahalanobis and geographic distances (Fig. 4, top). As amatter of fact, very distant populations of the same region,such as North Beni of Bolivia versus French Guyana (bothbelonging to the Amazon region), could not disclose true,significant allometry-free differences (L. ayrozai, L. genic-ulata, L. whitmani, L. umbratilis, L. hirsutus), while closerpopulations, such as the Sub-Andean (Uyuni) and Chaco
(Villamontes) samples in Bolivia, could show significantones. Unlike its showing poor relationships with geographi-cal distances, allometry-free variation disclosed a highly sig-nificant association with altitudinal differences (Fig. 4, bot-tom), indicating its possible link with temperatures and/orvegetation cover. Note that size changes, although indepen-dent from allometry-free variation, showed the same pat-terns of association with geographic distances (Fig. 3, top)or altitudinal differences (Fig. 3, bottom).
In sum, the correspondence was that of metric divergencewith altitudinal changes, i.e. ecological discontinuities,more than with distances separating populations. Globally,these results suggest an effect of ecological differences onmorphological constitution (Strauss, 1985). Such hypoth-esis conforms with the ecology of sandflies. Sandflies areknown primarily as hematophagous (female) insects, vec-tors ofLeishmania sp. (Protozoa, Kinetoplastidae), but theirecology is closely tied to that of plants: their larvae feed onground debris, males exclusively feed on plants and so do thefemales for their very first meal (Hamilton and El-Naiem,2000). Environmental effects on (female) sandfly biologyhave already been demonstrated for other attributes thanmetric properties (Lane, 1988), such as for instance theirvectorial capacity (Lainson et al., 1976; Le Pont and Pajot,1981; Dujardin et al., 1993; Rangel et al., 1996; Azevedoet al., 2001), or their anthropophily (Campbell-Lendrumet al., 1999). As long as other genetic attributes are probablyselected in different environments, the metric inter-regionaldisparity in sandflies could be a relevant indicator of di-verging microevolutionary processes.
The present study showed that in sandflies (i) size per sewas a rather labile feature within a species, (ii) the stabil-ity of size-independent variation was true only under certainconditions and (iii) significant, abrupt intraspecific metricvariation occurred across large ecogeographic areas of SouthAmerica. These points are relevant to the use of morphomet-rics in species diagnostic. They also are compatible with theidea of a direct influence of environment on morphologicalevolution. Sandfly species having crossed critical ecologicalbarriers (dispersal), or having been separated by them (vi-cariance), have probably been selected for genetic attributesaffecting their morphological stability through pleiotropyand/or epigenetic mechanisms (Schlichting and Pigliucci,1998).
Acknowledgements
To Brisola Marcondes (Florianopolis, Brazil) and to Eu-nice Galati (Sao Paulo, Brazil) for providing us with Brazil-ian samples ofLutzomyia ayrozai andL. geniculata. To Dr.Ronaldo Andrade, director del INLASA (Instituto Nacionalde Laboratorios de Salud) and Dr. Sergio Mollinedo, direc-tor of the Department of Parasitology (INLASA) for theirkind support.

358 J.-P. Dujardin, F. Le Pont / Infection, Genetics and Evolution 4 (2004) 353–359
References
Airoldi, J.P., Flury, B.K., 1988. An application of common principalcomponent analysis to cranial morphometry ofMicrotus californicusandM. ochrogaster (Mammalia Rodentia). J. Zool. Lond. 216, 21–36.
Añez, N., Valenta, D.T., Cazorla, D., Quicke, D.J., Feliciangeli, D., 1997.Multivariate analysis to discriminate species of phlebotomine sand flies(Diptera: Psychodidae):Lutzomyia townsendi, L. spinicrassa, and L.youngi. J. Med. Entomol. 34, 312–316.
Azevedo, A.C.R., Souza, N.A., Meneses, C.R.V., Costa, W.A., Costa,S.M., Lima, J.B., Rangel, E.F., 2001. Ecology of the sandflies (Diptera:Psychodidae: Phlebotominae) in the North of the State of Mato Grosso,Brazil. Mem. Inst. Oswaldo Cruz 97, 459–464.
Baylac, M., Daufresne, T., 1996. Wing venation variability inMonarthropalpus buxi (Diptera, Cecidomyiidae) and the quaternarycoevolution of Box (Buxus sempervirens L.) and its midge. Ageometrical morphometric analysis. In: Marcus, L.F., Corti, M., Loy,A., Naylor, G.J.P., Slice, D. (Eds.), Advances in Morphometrics.Proceedings of the 1993 NATO-ASI on Morphometrics NATO ASI, Ser.A, Life Sciences (587 pp.). Plenum Publishers, New York, pp. 285–301.
Bonnefoy, S., Tibayrenc, M., Le Pont, F., Dujardin, J.P., Desjeux, P.,Ayala, F.J., 1986. An isozymic study ofLutzomyia longipalpis (Diptera,Psychodidae), the vector of visceral leishmaniasis in the Yungas(Bolivia). Cah. O.R.S.T.O.M., sér. Ent. méd et Parasitol XXIV (3),213–217.
Bookstein, F.L., 1996. Combining the tools of geometric morphometrics.In: Marcus, L.F., Corti, M., Loy, A., Naylor, G.J.P., Slice, D. (Eds.),Advances in Morphometrics. Proceedings of the 1993 NATO-ASI onMorphometrics, NATO ASI, Ser. A, Life Sciences (587 pp.). PlenumPublishers, New York, pp. 131–151.
Burnaby, T.P., 1966. Growth-invariant discriminant functions andgeneralized distances. Biometrics 22, 96–110.
Caillard, T., Tibayrenc, M., Lepont, F., Dujardin, J.P., Desjeux, P., Ayala,F.J., 1986. Diagonosis by isozyme methods of 2 cryptic species,Psychodopygus carrerai and Psychodopygus yucumensis (Diptera,Psychodidae). J. Med. Entomol. 23 (5), 489–492.
Campbell-Lendrum, D.H., Pinto, M.C., Brandao Filho, S.P., Souza,A.A., Ready, P.D., Davies, C.R., 1999. Experimental comparisonof anthropophily between geographically dispersed populations ofLutzomyia whitmani (Diptera: psychodidae). Med. Vet. Entomol. 13,299–309.
Chapman, F.M., 1917. The distribution of bird-life in Colombia: acontribution to a biological survey of South America. Bull. Am. Mus.Nat. Hist. 36, 1–729.
Claridge, M.F., Gillham, M.C., 1992. Variation in populations ofleafhoppers and planthoppers (Auchenorrhyncha): biotypes andbiological species. In: Foottit, R.G., Sorensen, J.T. (Eds.), Ordinationin the Study of Morphology, Evolution and Systematics of Insects:Applications and Quantitative Genetic Rationales (418 pp.). Elsevier,New York, pp. 241–259.
Cracraft, J., Prum, R.O., 1988. Patterns and processes of diversification:speciation and historical congruence in some neotropical birds.Evolution 42, 603–620.
Dujardin, J.C., Llanos-Cuentas, A., Caceres, A., Arana, M., Dujardin,J.P., Guerrini, F., Gomez, J., Arroyo, J., De Doncker, S., Jacquet, D.,Hamers, R., Guerra, H., Le Ray, D., Arevalo, J., 1993. Molecularkaryotype variation ofLeishmania (Viannia) peruviana: indicationof geographical populations in Peru along a North-South cline. A.Trop. Med. Parasitol. 87, 335–347.
Dujardin, J.P., Le Pont, F., 2000. Morphometrics of a sandfly subspecies:L. carrerai thula. Comptes Rendus de l’Académie des Sciences, Paris.Life Sci. 323, 1–7.
Dujardin, J.P., Le Pont, F., Baylac, M., 2002. Geographic versusinterspecific differentiation of sandflies: a landmark data analysis. Bull.Entomol. Res. 93, 87–90.
Dujardin, J.P., Le Pont, F., Martinez, E., 1999. Quantitative morphologicalevidence for incipient species withinLutzomyia quinquefer (Diptera:Psychodidae). Mem. Inst. Oswaldo Cruz 94, 829–836.
Dujardin, J.P., Torrez, E.M., Le Pont, F., Hervas, D., Sossa, D., 1997.Isozymic and metric variation in theLutzomyia longipalpis complex.Med. Vet. Entomol. 11, 394–400.
Dujardin, J.P., Le Pont, F., Cruz, M., Leon, R., Tarrieu, F., Guderian, R.,Echeverria, R., Tibayrenc, M., 1996. Cryptic speciation inLutzomyia(nyssomyia) trapidoi (Fairchild and Hertig) (Diptera: Psychodidae)detected by Multilocus Enzyme Electrophoresis. Am. J. Trop. Med.Hyg. 54 (1), 42–45.
Gebre-Michael, T., Medhin, G., 1997. Morphometric separation ofFemales ofPhlebotomus (Phlebotomus) duboscqi andP. (P.) bergeroti(Diptera: Psychodidae). J. Med. Entomol. 34 (4), 383–386.
Hamilton, J.G.C., El-Naiem, D.A., 2000. Sugars in the gut of the sandflyPhlebotomus orientalis from Dinder National Park, Eastern Sudan.Med. Vet. Entomol. 14, 64–70.
Houck, M.A., Gauthier, J.A., Strauss, R.E., 1990. Allometric scaling inthe Earliest Fossil Bird,Archaeopteryx lithographica. Sciences 247,195–198.
Hutcheson, H.J., Oliver, J.H., Houck, M.A., Strauss, R.E., 1995.Multivariate morphometric discrimination of nymphal and adult formsof the blacklegged tick (Acari: Ixodidae), a principal vector of theagent of lyme disease in Eastern North America. J. Med. Entomol.32, 827–842.
Klingenberg, C.P., 1996. Multivariate allometry. In: Marcus, L.F.,Corti, M., Loy, A., Naylor, G.J.P., Slice, D., (Eds.), Advancesin Morphometrics. Proceedings of the 1993 NATO-ASI onMorphometrics, NATO ASI, Ser. A, Life Sciences (587 pp.). PlenumPublishers, New York, pp. 23–49.
Kreutzer, R.D., Palau, M.T., Morales, A., Ferro, C., Felicianceli, D.,Young, D.G., 1990. Genetic relationships among phlebotomine sandflies (Diptera: Psychodidae) in theverrucarum species group. J. Med.Entomol. 27 (1), 8.
Lainson, R., Ward, R.D., Shaw, J.J., 1976. Cutaneous leishmaniasis inNorth Brazil: Lutzomyia anduzei as a major vector. Trans. R. Soc.Trop. Med. Hyg. 70, 171–172.
Lane, R.P., 1988. Geographic variation in Old World phlebotominesandflies. In: Service, M.W. (Ed.), Biosystematics of HaematophagousInsects. Systematics Association Special Volume No. 37. OxfordUniversity Press Oxford, UK, pp. 77–90, ISBN 0-19-857709-5.
Lane, R.P., Ready, P.D., 1985. Multivariate discrimination betweenLutzomyia wellcomei, a vector of mucocutaneous leishmaniasis, andLu. complexus (Diptera: Phlebotominae). A. Trop. Med. Parasitol. 79,469–472.
Lanzaro, G.C., Warburg, A., 1995. Genetic variability in phlebotominesandflies: possible implications for leishmaniasis epidemiology.Parasitol. Today 11 (4), 151–154.
Lanzaro, G.C., Ostrovska, K., Herrero, M.V., Lawyer, P.G., Warburg, A.,1993.Lutzomyia longipalpis is a species complex: genetic divergenceand interspecific hybrid sterility among three populations. Am. J.Trop. Med. Hyg. 48 (6), 839–847.
Lebbe, J., Torrez, M.E., Vignes, R., Dedet, J.P., 1991. Morphométrie alaireet caractérisation spécifique des phlébotomes. Parassitologia 33 (Suppl.1), 365–372.
Mahalanobis, P.C., 1936. On the generalized distance in statistics. Proc.Natl. Inst. Sci. India 2, 49–55.
Marcondes, C.B., Lozovei, A.L., Galati, E.A.B., Taniguchi, H.H., 1998.The usefulness of Bergmann’s rule for the distinction of membersof Lutzomyia intermedia species complex (Diptera, Psychodidae,Phlebotominae). Mem. Inst. Oswaldo Cruz RJ 93, 363–364.
Nei, M., 1987. Molecular Evolutionary Genetics. Chapter IX. GeneticDistance Between Populations. Columbia University Press, NY, 512pp.
Pearson, K., 1926. On the coefficient of racial likeliness. Biometrika 18,337–343.
Rangel, E.F., Lainson, R., Souza, A.A., Ready, P., Azevedo, A.,1996. Variation between geographical populations ofLutzomyia(nyssomyia) whitmani (Antunes and Coutinho, 1939) sensu lato(Diptera: Psychodidae: Phiebotominae) in Brasil. Mem. Inst. Oswaldocruz, Rio de Janeiro 97 (1), 43–50.

J.-P. Dujardin, F. Le Pont / Infection, Genetics and Evolution 4 (2004) 353–359 359
Schlichting, C.D., Pigliucci, M., 1998. Phenotypic Evolution: A ReactionNorm Perpective, Sinauer Associates, Inc., Sunderland, MA, pp. 387.
Strauss, R.E., 1985. Static allometry and variation in body form in theSouth American catfish genusCorydoras (callichthyidae). Syst. Zool.34, 381–396.
Young, D.G., Duncan, M.A., 1994. Guide to the Identification andGeographic Distribution ofLutzomyia Sand Flies in Mexico, the WestIndies, Central and South America (Diptera: Psychodidae), AssociatePublishers, Gainesville, Fl, pp. 1059.



![Cégep de Saint-Félicien · 2016. 9. 12. · -- National Geographic Society (É.-U.) Gennevilliers] : National Geographic France, 2012. Bibliothèque — Collection générale 910.202](https://img.pdfslide.fr/doc/110x75/6026bbbcab9d3b67cb2a6ea4/cgep-de-saint-f-2016-9-12-national-geographic-society-u-gennevilliers.jpg)