Embed Size (px)
Citation preview

EXPERIMENTAL AND MOLECULAR PATHOLOGY 27, 68-80 ( 1977)
Hepatocellular Necrosis During Frog Virus 3-Induced Hepatitis of Mice: An Electron Microscopic Study1
ANNICK BINCEN AND ANDRE KIRN
Groupe de Recherches de 2’l.N.S.E.R.M. sur la Pathogdnie des Infections Virabs,
Laboratoire de Virologie de la Faculte’ de Mtklecine, Unioersitd Louis Pasteur, 3 rue Koeberld, 67000 Strasbourg, France
Receioed July 7, 1976, and in revised form February 18, 1977
The toxic degenerative hepatitis induced by Frog Virus 3 in mice is characterized by late cytoplasmic alterations of the hepatocytes as well as early nuclear lesions. These alterations vary according to the dose of virus used. The inoculation of one lethal dose is followed by an alteration of both rough and smooth endoplasmic reticula (appearance of fingerprints and hypertrophy of SER), an accumulation of lipids, and a final necrosis. With high doses of virus (20 LD) lesions of a much more drastic nature can be shown: The RER disrupts and even disappears. Mito- chondrial alterations can be seen just before the death of the animals, whereas steatosis and SER hypertrophy have never been observed.
Frog Virus 3 (FV,) inoculated intraperitoneally into mice produces an acute degenerative hepatitis characterized by severe pertubations of the macro- molecular metabolism of the liver [Biosynthesis of cellular RNA, DNA, and proteins is rapidly inhibited (Elharrar et al., 1973)] leading to the death of the animals within 18-24 hr ( Bingen-Brendel et al., 1972; Kirn et al., 1972). This hepatitis is of toxic origin since the virus, whose multiplication is a highly thermosensitive one, does not replicate in the mice. The toxicity of FV, is re- lated to structural proteins which can be solubilized from the virus particles ( Aubertin et al., 1973). Electron microscopic studies have shown that the earliest lesions of the hepatocytes are nuclear ones ( Bingen and Kirn, 1973), the cyto- plasm remaining free of alterations at the beginning of the infection. In this paper we describe the late cytoplasmic changes which appear at a time when the nuclear lesions are complete.
MATERIAL AND METHODS
The techniques of cell cultures, virus production, and titration have been previously described ( Bingen-Brendel et al., 1972). Inbred mice (IC strain), weighing from 15 to 20 g, were inoculated intraperitoneally with either one lethal dose (LD), corresponding to 2 x lo7 plaque-forming units (PFU), or 20 LD (4 x 10s PFU), depending on the experiment. Fifteen mice were used for each experiment. They were sacrificed by decapitation, 11 or 16 hr after the inoculation of 1 LD and 8 or 10 hr after the inoculation of 20 LD. Specimens
1 This work was supported in part by D.R.M.E. (Contract No. 75/123).
Copyright @ 1977 by Academic Press, Inc. All rights of reproduction in any form reserved. ISSN 0014-4500

FVrINDUCED NECROSIS OF MOUSE HEPATOCYTES 69
from different lobes of the liver were cut into pieces smaller than 1 mm3 with the aid of a razor blade and immediately fixed as already described (Bingen and Kirn, 1973). They were then sectioned wit,h an Ultramicrotome III LKB equipped with a diamond knife, and the sections were stained with uranyl acetate and lead citrate according to Reynolds (1963a). Bernhard’s technique (1968, 1969) for ethylenediaminetetraacetic acid treatment was applied to several specimens. Electron microscopic examinations were carried out in a Philips EM 300 electron microscope.
Adjacent thicker sections (0.5 to 1~) were cut with an Ultramicrotome III LKB equipped with glass knives and were stained with toluidine blue according to the method of Trump et al. (1961).
RESULTS
A. Nuclear Alterations of the Hepatocytes
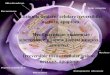
Figure 1, which represents a micrograph taken 12 hr after the inoculation of 1 LD, sums up the typical nuclear changes. The nucleus has an irregular, sinuous shape and shows an important condensation of the perinuclear chromatin. Numerous interchromatin granules are agglomerated in dense clusters at the center of the nucleoplasm, and, within them, near the nucleolus, two bundles of fibrillar material are visible. These particular nuclear bodies (Fig. la), when observed at higher magnification, are seen to be composed of numerous filaments ranging from 7 to 8 nm in diameter and arranged roughly parallel to one another.
The disappearance of the nucleoplasmic fibrillar material represents another striking modification in the hepatocyte nuclei during FV,-induced hepatitis and is even more evident after EDTA treatment. Figure 2 shows a mouse hepato- cyte nucleus after 4 hr of infection with 20 LD of FV,; the nucleoplasm seems to be completely emptied of its fibrillar matter. It should be emphasized that neither nucleolar segregation nor fragmentation have ever been observed. The only nucleolar modification consists of an increase in the fibrillar material fol- lowed by an annulation of the nucleoli later on during infection (Bingen, 1975). All these modifications appear in the hepatocyte nuclei of the infected mice in- dependent of the dose of virus administered, the changes being evident 1 hr after inoculation with 20 LD, whereas with 1 LD they take 4 to 6 hr to appear. Numerous observations of the livers of control mice inoculated with phosphate- buffered saline in place of the virus have never shown any nuclear alterations of the hepatocytes.
B. Cytoplasmic Alterations of the Hepatocytes
The cytoplasmic alterations differ considerably according to the dose of virus used.
(a) Cytoplasmic modifications observed after the inoculation of 1 LD. To begin with, thin sections were examined under a light microscope in order to establish the topography of the lesions. Until the 12t,h hr of infection, the ap- pearance of numerous vacuoles of steatosis constituted the only modification in the cytoplasm. Figure 3 (15 hr after infection) shows periporta1 necrotic foci surrounded by a layer of undamaged cells arranged around the centrolobular
70 BINGEN AND KIRN
FIGS. 1 and 2. Nuclear modifications. Fig. 1: The nucleus has an irregular shape and shows an important condensation of the perinuclear chromatin (Ch). In the nucleoplasm numerous interchromatin granules (Ig) are agglomerated in a large cluster. Near the nucleolus, essentially composed of its fibrillar material ( f ), one can observe two filamentous bodies ( Fi). X 16,000; 12 hr after inoculation of 1 LD (2 X 10’ PFU). Fig. la: A large bundle composed of fila- ments arranged in parallel, in longitudinal section. At right (arrow), ramifications of the bundle observed in transversal section. X37,500; 8 hr after inoculation of 1 LD (2 x 1Or PFU). Fig. 2: After EDTA treatment one can notice that the nucleoplasmic fibrillar ma- terial has disappeared. The chromatin (Ch) associated with the nuclear membrane and the nucleolus remains unstained. The nucleolus (Nu) and the interchromatin granules (Ig) are heavily stained. X 17,000; 4 hr after inoculation of 20 LD (4 X 10” PFU).

FVa-INDUCED NECROSIS OF MOUSE HEPATOCYTES 71
FIGS. 3 and 4. Light microscopy. Fig. 3: Pericentrolobular necrosis. A layer of cells around the centrolobular vein (C.L.V.) is undamaged. X420; 15 hr after inoculation of 1 LD. Fig. 4: Diffused necrosis from the portal vein (P.V.) to the centrolobular vein (C.L.V.), not in- volving some cells around the C.L.V. X350; 20 hr after inoculation of 1 LD.
vein. In the necrotic area the nuclei are pycnotic, the cytoplasm is vacuolized, and congestion of the sinusoids is evident. At this point during infection the most severely affected cells are located in the periportal region, Figure 4 (20 hr after infection) shows diffused necrosis of the hepatice lobule; however, peri- centrolobular cells are not involved in this necrosis.
The first ultrastructural modifications observed consist of an accumulation of cytoplasmic lipid droplets and an alteration in the endoplasmic reticulum. As
72 BINGEN AND KIRN
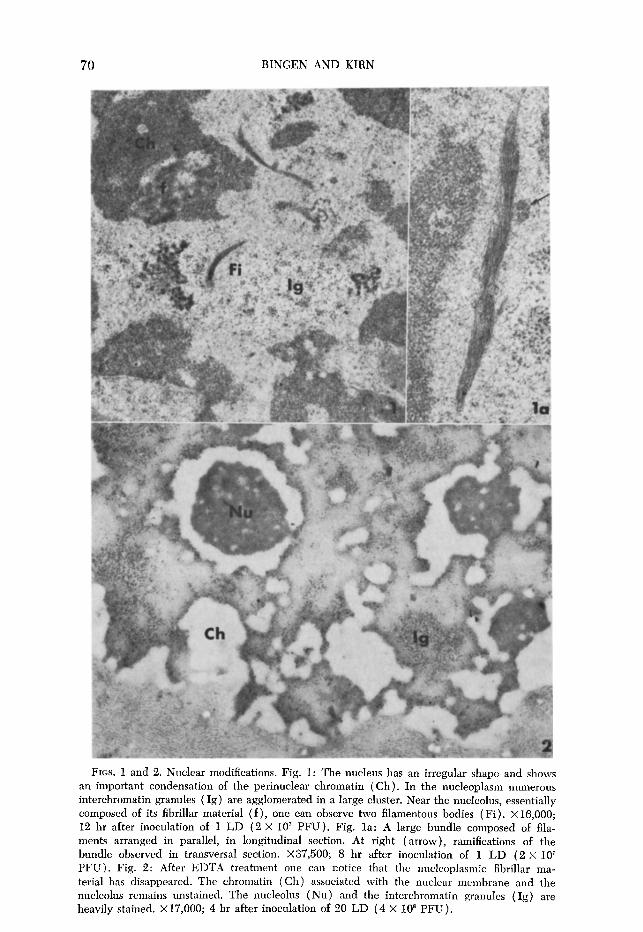
FIGS. 5-9. Cytoplasmic modifications observed after the inoculation of 1 LD (2 X 10’ PFU ). Fig. 5: The whole cytoplasmic area is invaded with fatty, lightly osmiophilic droplets. One can also note the presence of numerous glycogen particles. X8000; 12 hr after inocula- tion. Fig. 6: Concentric lamellar formations composed of 20 layers of paired membranes disposed concentrically around a mitochondrion. Short strips of these cisternae carry ribosomes (arrow). Peripheral lamellae are in close contact with the smooth-surfaced endoplasmic reticulum. Va:vacuole; X16,000; 15 hr after inoculation.
FVz-INDUCED NECROSIS OF MOUSE HEPATOCYTES ’ 73
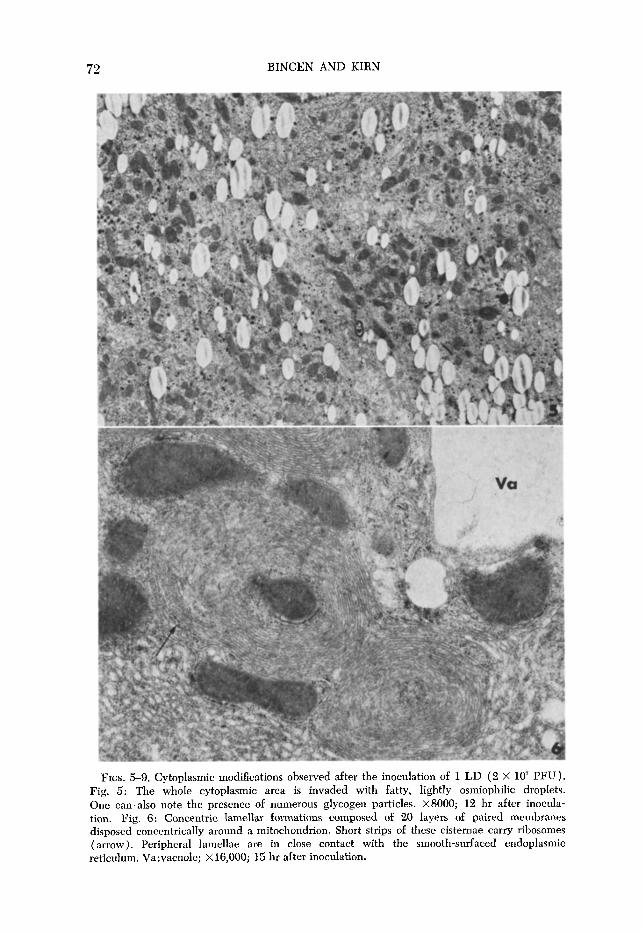
FIG. 7. Hypertrophy of the agranular reticulum. At the top left one can observe two small whorls of concentric lamellar structures (arrows). X10,000; 15 hr after inoculation.
FIG. 8. Involution of the smooth endoplasmic reticulum; Stage 1, the SER area is changed into a flaky material; stage 2, this material has lost its electron opacity; stage 3, represented by disintegration cavities surrounded by membranes. X6500; 15 hr after inoculation.
74 BINGEN AND KIRN
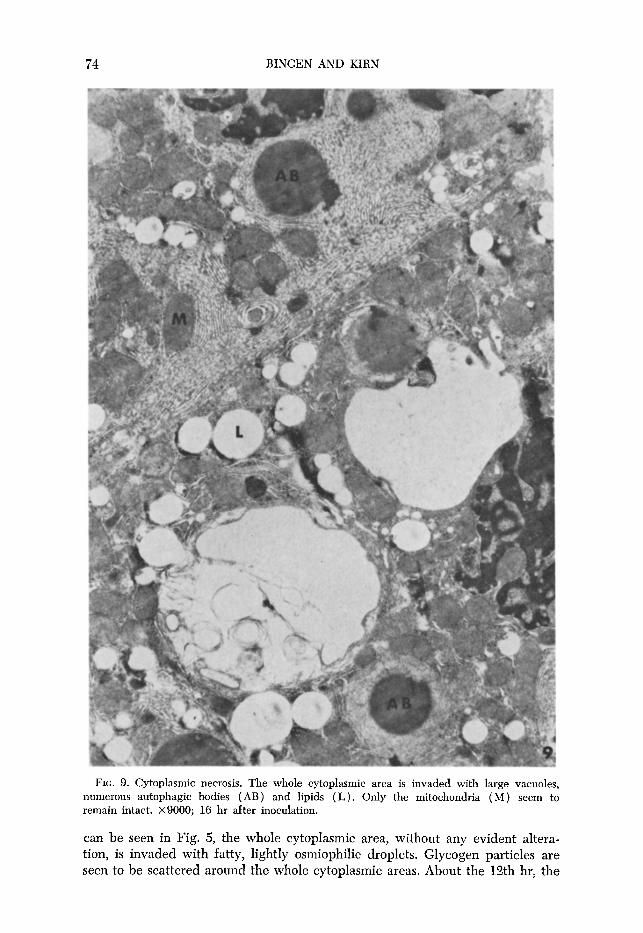
FIG. 9. Cytoplasmic necrosis. The whole cytoplasmic area is invaded with large vacuoles, numerous autophagic bodies ( AB) and lipids (L). Only the mitochondria ( M ) seem to remain intact. x9000; 16 hr after inoculation.
can be seen in Fig. 5, the whole cytoplasmic area, without any evident altera- tion, is invaded with fatty, lightly osmiophilic droplets. Glycogen particles are seen to be scattered around the whole cytoplasmic areas. About the 12th hr, the

FVrINDUCED NECROSIS OF MOUSE HEPATOCYTES 75
usual parallel array of rough endoplasmic reticulum (RER) is partially dis- organized, the long profiles being disrupted and curved around uninjured mito- chondria. After 14 to 15 hr of infection, concentric lamellar formations (Fig. 6)) which do not occur in normal hepatic parenchymal cells, can be observed. These structures, called fingerprints, consist of about 20 layers of paired mem- branes, devoid of ribosomes and disposed concentrically around a core, gen- erally a mitochondrion or a lipid droplet. Short strips of these cisternae carry ribosomes at the periphery of the whorls and around the encircled mitochondria or lipid droplets (Fig. 6). Generally, peripheral lamellae come into contact with the cytoplasm, joining either the membranes of another whorl or the tubular and vesicular elements of smooth-surfaced endoplasmic reticulum (SER) (bot- tom of Fig. 6). Besides the concentric lamellar formations, giant areas of smooth- surfaced membranes can also be seen. The expansion of SER, which forms a compact lattice (Fig. 7), occurs in an area free of other organelles. At this time, an apparent depletion of glycogen is noticeable in all of the hepatic parenchymal cells examined. The involution of smooth endoplasmic reticulum leads to the formation of vacuoles. One can postulate that this involution takes place as follows (Fig. 8) : The area of smooth-surfaced membranes is changed into a flaky material (stage 1) which disappears progressively (stage 2) leaving disintegration cavities surrounded by membranes ( stage 3). The intraperitoneal inoculation of low doses of FV, leads to cell necrosis after about 16 hr (Fig. 9). It is surprising that, at this time of complete cell necrosis, the mitochondria appear normal.
The cytoplasmic alterations of the hepatocytes described above were found in the livers of all 15 mice examined in our experiments, whereas they were never observed in the livers of control mice inoculated with phosphate-buffered saline.
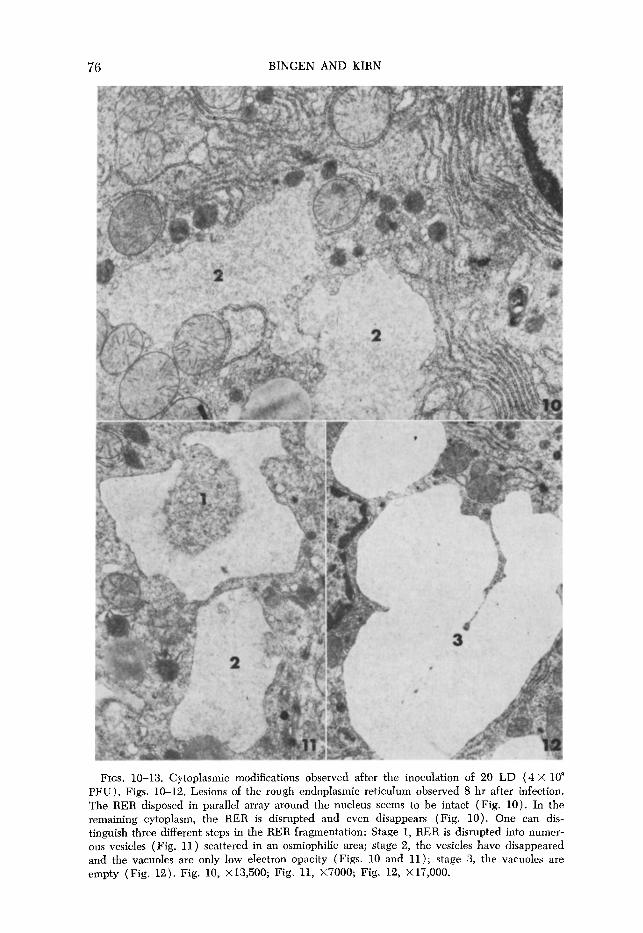
(b) Cytoplasmic modifications observed after the inoculation of 20 lethal doses (4~10~ PFU). Th e 1 esions observed after the inoculation of high doses of virus are different from those described above. At the 8th hr, more drastic alterations occur, such as the sudden disruption and even dissolution of RER membrane together with the simultaneous formation of vacuoles (Fig. lo), some of them containing numerous vesicles scattered in an osmiophilic area (Fig. 11). These vesicles, which carry ribosomes, may proceed from the frag- mentation of the RER (stage 1). In some vacuoles of only moderate electron opacity, like those shown in Figs. 10 and 11, the vesicles have disappeared (stage 2), and empty vacuoles may be considered to represent the final phase of RER disintegration (Fig. 12). From the morphological point of view, no apparent differences were noted in the chondrium of animals inoculated with FV, when compared to that of control animals. The chondrium structure is preserved for a long time, and alterations are observed only later on during the course of the infection, just before the death of the mice. At this point, few mitochondria appear normal, many showing irreversible alterations (Fig. 13). They tend to become swollen, their internal cristae are rather indistinct or even missing, and the mitochondrial matrix is fragmented; in other organelles the granular matrix is rather patchy and vacuolized.
It should be emphasized that, with high doses of FV,, steatosis or SER hy-
76 BINGEN AND KIRN
FIGS. 10-13. Cytoplasmic modifications observed after the inoculation of 20 LD (4 X 10” PFU). Figs. 10-12. Lesions of the rough endoplasmic reticulum observed 8 hr after infection. The RER disposed in parallel array around the nucleus seems to be intact (Fig. 10). In the remaining cytoplasm, the RER is disrupted and even disappears (Fig. 10). One can dis- tinguish three different steps in the RER fragmentation: Stage 1, RER is disrupted into numer- ous vesicles (Fig. 11) scattered in an osmiophilic area; stage 2, the vesicles have disappeared and the vacuoles are only low electron opacity (Figs. 10 and 11); stage 3, the vacuoles are empty (Fig. 12). Fig. 10, X 13,500; Fig. 11, X7000; Fig. 12, X 17,000.

FVs-INDUCED NECROSIS OF MOUSE HEPATOCYTES 77
FIG. 13. Mitochondria alterations, 9 hr 40 min after infection. Late in the course of in- fection, the mitochondria shbw irreversible alterations. They tend to become swollen, their internal cristae are indistinct or even missing, and the mitochondrial matrix is fragmented or vacuolized. X 15,500.
pertrophy have never been observed, whereas glycogen depletion occurs as early as 8 hr after infection. These drastic cytoplasmic alterations could be seen in all 15 mice used in those experiments involving a high virus dose.
DISCUSSION
Most of the nuclear lesions induced by FV, in the hepatocytes are similar to those observed after the inoculation of drugs which depress RNA synthesis, such as a-amanitin (Fiume et al., 1969; Meyer-Schulz and Porte, 1971). Con- cerning the nucleolar changes, however, some fundamental differences were noticed. Nucleolar fragmentation or segregation were never observed in FV,- induced hepatitis, the main alteration of the nucleolus being characterized by the appearance of ring-shaped nucleoli made up essentially of fibrillar material (Bingen, 1975). This demonstrates that an inhibition of RNA synthesis does not necessarily result in nucleolar segregation. A similar result was obtained by Derenzini and Bonetti (1975) who showed that the administration of large doses of cycloheximide, which led to a halt in RNA synthesis in the rat liver, did not produce any typical nucleolar lesion of the hepatocyte nuclei. The presence of fibrillar bodies in the hepatocyte nucleoplasm seems to be specific to FV3 infection, but its significance is completely unknown (Bingen and Kirn, 1975). It is interesting to notice that, in the hepatitis produced with FV,- solubilized proteins, the same nuclear bodies could be observed ( Aubertin et al., 1977). The cytoplasmic alterations of the hepatocytes appear long after the

78 BINGEN AND KIRN
inoculation of FV,, while, by way of comparison, the nuclear lesions occur as early as 1 hr after infection (Bingen-Brendel et al., 1972; Bingen and Kim, 1973). On the other hand, the cytoplasmic lesions differ according to the viral input, whereas the nuclear changes, with the exception of their intensity, do not (Bingen, 1975). The considerable alterations of RER, including cisternal dis- ruption and fragmentation and even membrane dissolution are observed after the inoculation of 20 LD. Similar changes have already been described after the administration of diverse hepatotoxic compounds such as CCI, (Bassi, 1960; Reynolds, 196313)) phosphorus (Jezequel, 1958)) ally1 formate ( Rouiller, 1964)) and 2-methyl-4-dimethylaminoazobenzene (Lafontaine and Allard, 1964). The endoplasmic reticulum alterations observed after the inoculation of 1 LD are characterized by concentric lamellar structures and by SER hypertrophy. Finger- print configurations have also been reported after the administration of aflatoxine B1 ( Svoboda et al., 1966), ethionine ( Meldolesi et al., 1967)) thioacetamide ( Salomon, 1962)) a-naphtylisothiocyanate ( Steiner and Baglio, 1963)) and CC14 (Stenger, 1966). Investigations into the effects of various toxic agents on the liver (Steiner and Baglio, 1963; Steiner et nl., 1964) have concluded that these configurations represent degenerative manifestations of the ergastoplasm. How- ever, Stenger (1966) has observed these structures in regenerative nodules, and Benedetti and Emmelot (1966) have regarded hepatic fingerprint configura- tions as evidence of ergastoplasm regeneration. Since, in liver regeneration, the in vivo rate of translation is at least the same as under nonregenerative condi- tions (Scornik, 1975), it would be surprising if, in FVa hepatitis, where the macromolecular metabolism (including the protein synthesis of the liver) is severely depressed ( Elharrar et al., 1973)) these figures represented regeneration. Proliferation of the smooth-surfaced elements may have something to do with a detoxication reaction. Such a hypertrophy of the smooth reticulum has been implicated in drug detoxication after the injection of several hepatotoxic sub- stances. Since no modification of the smooth-surfaced elements occurs after FV3 poisoning, it may be postulated that it is the properties of this system which are affected.
In the mice inoculated with 1 LD of FV, there is a considerable accumulation of lipids within the hepatocytes. It has been claimed that one important pro- tein thought to be manufactured by the hepatic RER is a lipid-acceptor protein (Stenger, 1970). A n insufficiency of this protein, with which the lipids may be complexed for transport from the liver to the plasma, may be responsible for fatty livers. It is likely that the steatosis in FV3 hepatitis is induced by the disorganization of the hepatic RER accompanied by a defect in liver protein synthesis (Elharrar et al., 1973). Such a mechanism has been invoked to ex- plain the hepatotoxic effect of yellow P (Fiume et al., 1969), thioacetamide (Barker et al., 1963), dimethylnitrosamine ( Rees and Shotlander, 1963) and ethionine (Harris and Robinson, 1961). Glycogen begins to disappear 8 hr after inoculation with a high virus dose and 12 hr after inoculation with 1 LD. This observation corresponds closely to the severe hypoglycemia present at about the same time in the mice (Imler et al., 1975).
With low doses of virus the structures of the mitochondria are well preserved, even at a time when the cells show necrosis. With high doses of virus, how- ever, severe mitochondrial alterations occur: The mitochondrial matrix is frag- mented, and the mitochondria are vacuolized just before the death of the

FVa-INDUCED NECROSIS OF MOUSE HEPATOCYTES 79
animals. These ultrastructural observations bear out biochemical studies which show a 40% inhibition of the succinic dehydrogenase activity at 10 hr post- infection (Elharrar, personal communication). As with CC4 ( Smuckler et al., 1962), alterations in mitochondrial enzyme activity appear later than the period during which altered protein synthesis was first shown.
The question of the relationships between the nuclear and the cytoplasmic alterations is somewhat complex. Although the nuclear lesions occur first, it cannot be concluded that the cytoplasmic alterations are the consequence of the nuclear picnosis, as the necrosis of the cytoplasm could be produced by another viral component different from the one acting at nuclear level. The inhibition of the transcription and the translation produced by FVs-soluble proteins in tissue cultures seems to have two independent effects ( Aubertin and Kirn, 1975). On the other hand, it may be admitted that the cytoplasmic lesions are secondary to the lesions of the sinusoidal cells produced by the virus which enters and destroys the Kupffer celIs (Bingen, 1975). Fiume ( 1975) has demonstrated that the administration of p-amanitin to the rat led to the death of the animals by nephritis; in this case the hepatocytes showed drastic nuclear lesions but no cytoplasmic alterations. However, when a p-amanitin-albumin conjugate was inoculated, it produced an acute hepatitis, and both lesions of the sinusoidal cells and hepatocyte necrosis could be demonstrated.
ACKNOWLEDGMENTS
We wish to express our gratitude to Mrs. Michelle Valle, Genevieve Reinhart, and Mr. Francis Lecerf for their technical assistance.
REFERENCES
AUBERTIN, A. M., HIRTH, C., TRAVO, C., NONNENMACHER, H., and Knw, A. (1973). Prep- aration and properties of an inhibitory extract from frog virus 3 particles. I. Viral. 11, 6947701.
AIJBERTIN, A. M., et KIRN, A. ( 1975). I n “Les Colloques de l’Institut national de la Sante et de la recherche medical,” Vol. 47, pp. 347-354. INSERM.
AURERTIN, A. M., ANTON, M., BINGEN, A., ELHARFCAR, M., and KIRN, A. (1977). Solubilized viral proteins produce a fatal hepatitis in mice. Nature (London) 265, 456-467.
BASSI, M. (1960). Electron microscopy of rat liver after carbon tetrachoride poisoning. Exp. Cell Rcs. 20, 313-324.
BARKER, E. A., SMUCKLER, E. A., and BENDITT, E. P. (1963). Effects of thioacetamide and yellow phosphorus poisoning on protein synthesis in vivo. Lab. Invest. 12, 955-960.
BENEDETTI, E. L., and EMMELOT, P. (1960). In “Proceedings European Regional Conference on Electron Microscopy, Del&” (A. L. Houwink and B. J. Spit, Eds.,) Vol. 2, pp. 875-878. Nederlandse Vereniging Voor Electronen Microscopy.
BERNHARD, W. (1968). Une methode de coloration regressive a l’usage de la microscopic Blectronique. C.R. Acad. Sci. P&s 267, 2170-2173.
BERNHARD, W. ( 1969). A new staining procedure for electron microscopical cytology. J. Uhzstruct. Res. 27, 259-265.
BINGEN-BFLENDEL, A., BATZENSCHLAGER, A., GUT, J. P., HIRTH, C., VETTER, J. M., et KIN, A. ( 1972). Etude histologique et virologique de l’hepatite degenerative aigue provoquee par le FV3 (Frog Virus 3) chez la souris. Ann. Inst. Pasteur 122, 125-142.
BINGEN, A., et KIRN, A. ( 1973). Modifications ultrastructurales precoces des noyaux des hepato- cytes de souris au tours de l’hepatite deg&rerative aigue provoquee par le FVs (Frog virus 3). J. Ultrustrwt. Res. 45, 343-355.
BINGEN, A. (1975). Hepatotoxicite du Frog Virus 3 (FVs) chez la souris. Etude ultra- structurale des lesions des hepatocytes. Thbe de doctorat en Biologie Humaine, Uni- versite Louis Pasteur de Strasbourg, no. 1.

80 BINGEN AND KIRN
BINCEN, A., and KIHN, A. (1975). Fibrillar bod’ res in hepatocyte nuclei during the course of the toxic hepatitis produced by Frog Virus 3 in mice. J. Ultrastnrct. Res. 50, 167-173.
DERENZINI, M., and BONETTI, E. ( 1975). Cycloheximide-induced ultrastructural changes in hepatocyte nuclei in partially hepatectomized rats. Virchows Arch. B. Cell. Pathol. 19, 115-125.
ELHARRAR, M., HIRTH, C., BLANC, J., et KIRN, A. ( 1973). Pathogenic de l’hkpatite toxique de la souris provoquee par le FV, (Frog Virus 3) : Inhibition de la synthese des macro- molecules du foie. B&him. Biophys. Acta 319, 91-102.
FIUME, L., MAAINOZZI, V., and NARDI, F. (1969). The effects of amanitin poisoning on
mouse kidney. Brit. J. Exp. PathoZ. 50, 270-276. FIUME, L. ( 1975). In “Pathogenesis and Mechanisms of Liver Cell Necrosis” (D. Keppler,
Ed.), pp. 57-67. University Park Press, Baltimore Maryland. HARRIS, P. M., and ROBINSON, D. S. ( 1961). Eth ionine administration in the rat. 1. Effects
on the liver and plasma lipids and on the disposal of dietary fat. Biochem. J. 80, 352-360.
IMLER, M., SCHLIENGER, J. L., et KIRN, A. ( 1975). Hyperammoniemie et hypoglycemic au course de l’hepatile d&generative aigue provoquee chez la souris par le Frog Virus 3. Med. Chirg. Dig. 2, 9-12.
JEZEQUEL, A. M. ( 1958). Les elfetes de l’intoxication aigue au phosphore sur le foie de rat. Ann. Anat. Pathol. 3, 512-537.
KIFLN, A., GUT, J. P., BINGEN, A., et HIRTH, C. (1972). Acute hepatitis produced by Frog Virus 3 in mice. Arch. Gesamte Virusforsch. 36, 394497.
LAFONTAINE, J, C., and ALLARD, C. (1964). A light and electron microscope study of mor- phological changes induced in rat liver cells by the azo dye 2 Me-DAB. J. Cell. BioZ. 22, 143-172.
MELDOLESI, J., CLEMENTI, F., CHIESARA, E., CONTI, F., and FANTI, A. (1967). Cytoplasmic changes in rat liver after prolonged treatment with low doses of ethionine and adenine. Lab. Invest. 17, 267-275.
MEYER-SCHTJLIZ, F., et PORTE, A. ( 1971). S ur les modifications nucleaires provoquees par 1’ or-amanitine dans les hkpatocytes de souris. Cytobiologie (Band 3, Heft 3), 387400.
REES, K. R., and SHOTLANDER, V. L. (1963). Fat accmnulation in acute liver injury. Ann. N.Y. Acad. Sci. 104, 905-915.
REYNOLDS, E. S. (1963a). The use of lead citrate at high pH as an electron opaque stain in electron microscopy. J. Cell. Biol. 17, 209-212.
REYNOLDS, E. S. (196313). Liver parenchymal cell injury. I. Initial alterations of the cell following poisoning with carbon tetrachloride. J. Cell. Biol. 19, 139-157.
ROUILLER, C. (1964). In “The Liver” (C. Rouiller, ed.), Vol. 2, pp. 335-476. Academic Press, New York.
SALOMON, J. C. ( 1962). Modifications des cellules du parenchyme hepatique du rat sous Peffet de la thioacetamide. Etude au microscope electronique des lesions observkes B la phase tardive dune intoxication chronique. J. Ultrastruct. Res. 7, 293307.
SCORNIK, 0. A. (1975). In “Liver Regeneration after Experimental Injury” (R. Lesch and W. Reutter, eds. pp. 156-168 Stratton Intercontinental Medical Book, New York.
SMUCKLER, E. A., ISERI, 0. A., and BENDITT, E. P. ( 1962). An intracellular defect in pro-
tein synthesis induced by carbon tetrachloride. J. Exp. Med. 116, 55-71.
STEINER, J. W., and BAGLIO, G. M. (1963). Electron microscopy of the cytoplasm of parenchymal liver cells in or-naphtyl-isothiocyanate-induced cirrhosis. Lab. Invest. 12, 765-790.
STEINER, J. W., MIYAI, K., and PHILLJPS, M. J. (1964). Electron microscopy of membrane- particle arrays in liver cells of ethionin-intoxicated rats. Amer. J. PuthoZ. 44, 169-213.
STENGER, R. J. ( 1966). Concentric lamellar formations in hepatic paren&ymal cells of carbon
tetrachloride-treated rats. J. UZtrastruct. Res. 14, 240-253. STENGER, R. J. ( 1970). Organelle pathology of the liver. The en&plasmic reticulum. as-
troenterology 58, 554-574.
SVOBODA, D., GRADY, H. J., and HIGGINSON, J. (1966). Aflatoxin B1 injury in rat and monkey liver. Amer. J. PathoZ. 49, 1023-1051.
TRUMP, B. F., SMUCKLER, E. A., and BENDITT, E. P. ( 1961). A method for staining epoxy sections for light microscopy. J. UZtrastruct. Res. 5, 343.





![Atelier Inaugural - Datacraft · 2020. 9. 30. · (2013) Machine learning approach for automated screening of malaria parasite using light microscopic images [7] Gopakumar et al](https://img.pdfslide.fr/doc/110x75/60fbf4d22cec0a3b3e4d5e47/atelier-inaugural-datacraft-2020-9-30-2013-machine-learning-approach-for.jpg)