Embed Size (px)
Citation preview

Fd C/WI. ‘hrc. Vol. 23. No. 3. PP. 363-366. 1985 Printed m Circa1 Briuain. All rights reserved
0278-6915/X5 L3.00 + 0.00 Copyright 0 1985 Pcrgamon Press Ltd
HYDROCARBON HYDROXYLATION SYSTEM IN LIVER MICROSOMES FROM FOUR ANIMAL SPECIES
E. F. PERDU-DURAND and J. E. TULLIEZ* Laboratoire de Recherches sur les Additifs Alimentaires, Institut National de la Recherche
Agronomique. B.P. 3 - 31931 -Toulouse cedex, France
(Rewind 26 ./II/~ 1983; Rrtrisiorl receioed I8 May 1984)
Abstract-The irl vitro metabolism of It-heptadecane was investigated using hepatic microsomes from Hubbard chickens, New Zealand rabbits. Wistar rats and rainbow trout. Incubations with the 14C- labelled alkane for I hr showed that the rate of oxidation varied between species; the rate (per mg protein) in chickens was roughly 20-fold greater than the rate in trout, and roughly IO-fold greater than the rates in rats and rabbits. On the basis of cytochromc P-450 content. the rate of heptadecane metabolism was roughly ZO-Told greater in the chicken than in the other species. Moreover, the I,,, Soret band of the reduced cytochrome P-450 CO-comples was observed at 452 nm in the chicken. Correlation between the rate of heptadecane metabolism and irl uiuo storage levels is discussed.
INTRODLICTION
Foods contain even and odd long-chain normal paraffins (C, z to CXs) resulting from plant bio- synthesis and food-chain transfer involving animals. In addition mineral oils and paraffins are used in food processing.
When hydrocarbons are administered orally to animals, the amount of absorption dirfers according to the species (Cravedi, Tulliez, Choubert & Luquet, 1983; Ladan, 1970; Tulliez& Bories. 1975b). Some of the absorbed hydrocarbon is stored unchanged as the parent compound, while the major portion under- goes lipid metabolism.
The biotransformation of alkanes has received more attention since McCarthy (1964) showed that mammals were able to oxidize n-paraffins to the corresponding fatty acids. III vim tissue-distribution studies with labelled alkanes in rats have located the oxidation products in the liver (Kolattukudy & Hankin, 1966; Tulliez & Bories, 1978). while in vitro studies have shown that the mixed-function oxidase catalysing the w-oxidation of rl-paraffins was present in the microsomal fraction of the liver in rats and mice (Durand & Tulliez, 1980; Kusunose, Ichihara & Kusunose, 1969) and of the intestine in rabbits and guinea-pigs (Ichihara, Ishihara, Kusunose & Kusunose, 1981; Mitchell & Hiibscher, 1968).
The aim of the present work was to compare the hydrocarbon hydroxylating system in the hepatic microsomes of chickens. rats, rabbits and trout.
EXPERIMENTAL
Chrwicals. [8-‘4C]0Heptadecane (specific activity 27 mCi/mmol) was synthesized at CEA (Saclay, France). Glucose-6-phosphate. glucose-6-phosphate dehydrogenase and NADP were obtained from Sigma (St Louis. MO, USA). Nicotinamide and magnesium chloride were purchased from E. Merck (Darmstadt. FRG). All the solvents used for extract-
*To whom correspondence should be addressed.
ion and thin-layer chromatography were obtained from commercial sources and purified by distillation.
AGnals. Male Wistar rats (I 50-200 g), male New Zealand rabbits (2.1-2.5 kg) and male Hubbard chickens (c. 2 kg) were used in this study. The animals were housed at 21°C + 1°C with a relative humidity of 60-70% and a 12-hr light/dark cycle, and were fed standard commercial diets (UAR Villemoisson S/Orge) and water ad lib. Rainbow trout (Saltno gairdtleri R., 150-200 g) were obtained from the pisciculture of INRA at Donzacq (Landes). The trout (male or female) were placed in fresh- water ponds at I8 f 2°C under natural lighting conditions.
Preparation of liver microsomes. Animals were killed by exsanguination and the livers were removed and immersed quickly in ice-cold 0.1 M-phosphate buffer (pH 7.4). weighed and then homogenized in a Potter-Elvehjem homogenizer fitted with a polytetra- fluoroethylene pestle in four vols of 0.1 M-phosphate buffer. The homogenates obtained were centrifuged at 10,000 g at 4°C for 20 min. Microsomes were sedimented by centrifuging the supematant fractions at 105,000 g at 4°C for 60 min.
The microsomal pellet was suspended with gentle homogenization in ten vols of 0.1 M-phosphate buffer to obtain a protein concentration of approx- imately I mg/ml. The protein content of the micro- somal suspension was determined by the method of Lowry. Rosebrough, Farr & Randall (,I951) with bovine serum albumin as standard. Cytochrome P- 450 was measured by the CO difference spectra with a spectrophotometer (Uvikon 810, Kontron), using the method of Omura & Sato (1964).
Heptadecme hydrmuylatior~. The metabolism of heptadecane by hepatic microsomal suspension was measured in an incubation medium that consisted of 2 ml of microsomal suspension and 3 ml of phos- phate buffer (pH 7.4), containing 3 pmol NADP, 7.5 pmol glucose-6-phosphate, 25 pmol nicotin- amide, 2.6 units glucose-6-phosphate dehydrogenase and 20 pmol magnesium chloride. After an initial 5-min incubation, the reactions were begun by the
363
364 E. F. PERDU-DURAND and J. E. TULLW
addition of [‘4CJheptadecane in 50 ~1 methanol. The concentration of substrate was adjusted by the addition of the unlabelled hydrocarbon. Incubations were carried out at 17°C for trout and at 37°C for the other animal species. Reactions were stopped after I hr, unless otherwise stated, by the addition of I ml 10 y0 (w/v) trichloroacetic acid.
,!%.utraction o/hydrocarbon. The heptadecane and its hydroxylation products were extracted with I5 ml ethyl acetate. After filtration through anhydrous sodium sulphate, the organic solvent was evaporated to dryness and the residue was redissolved in 1 ml methanol.
Thit&yer chromatography. A sample of the methanolic solution obtained was applied to a thin- layer plate of Silica gel G (Merck) and separation was achieved using hexane-ether (9:1, v/v) as the developing solvent. Although it was possible to separate the two main metabolites, i.e. fatty alcohol and fatty acid, as previously described (Kusunose ef al. 1969), in this experiment it was more advan- tageous just to separate the substrate from the oxidation products.
0 10 20 30 40 50 60
Time (min) Heplodecone concentrollon IpM)
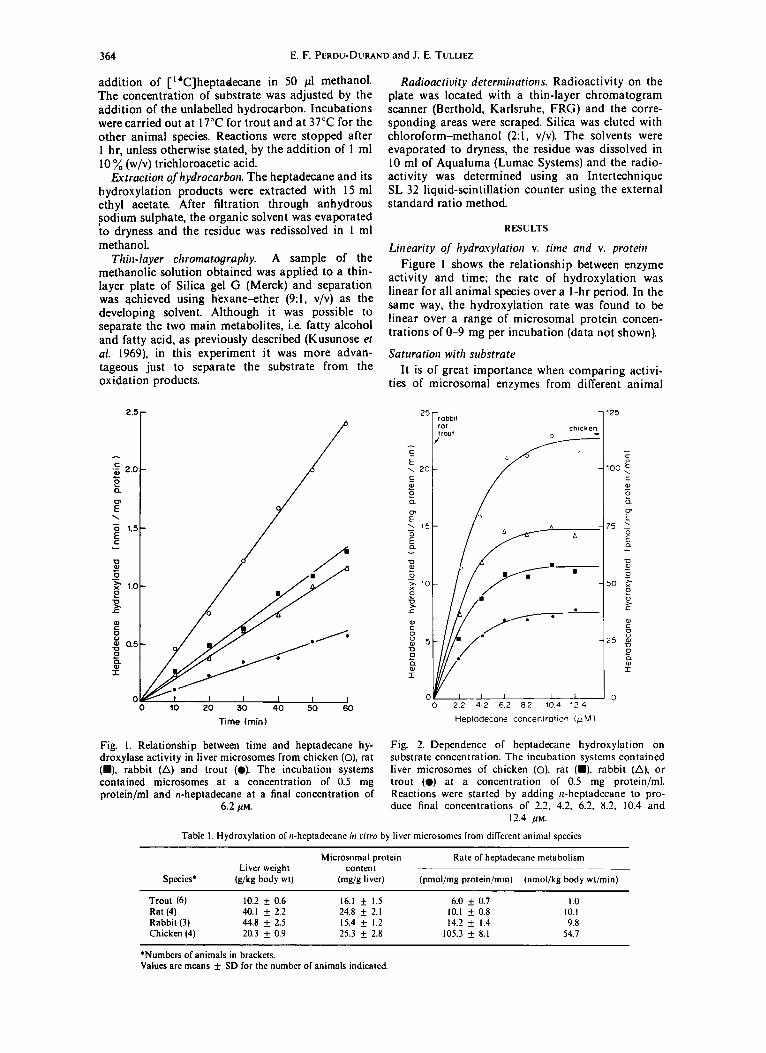
Fig. I. Relationship between time and heptadecane hy- droxylase activity in liver microsomes from chicken (0), rat (W), rabbit (A) and trout (a). The incubation systems contained microsomes at a concentration of 0.5 mg protein/ml and n-heptadecane at a final concentration of
6.2 pu.
Fig. 2. Dependence of heptadecane hydroxylation on substrate concentration. The incubation systems contained liver microsomes of chicken (0). rat (m). rabbit (A), or trout (0) at a concentration of 0.5 mg protein/ml. Reactions were started by adding rl-heptadecane to pro- duce final concentrations of 2.2, 4.2. 6.2. 8.2, 10.4 and
12.4 pu.
Radioactioity deteminatior~s. Radioactivity on the plate was located with a thin-layer chromatogram scanner (Berthold, Karlsruhe, FRG) and the corre- sponding areas were scraped. Silica was eluted with chloroform-methanol (2:1, v/v). The solvents were evaporated to dryness, the residue was dissolved in 10 ml of Aqualuma (Lumac Systems) and the radio- activity was determined using an Intertechnique SL 32 liquid-scintillation counter using the external standard ratio method.
RESULTS
Lhearity of hydro.yylatiorl v. the md v. proteirl
Figure 1 shows the relationship between enzyme activity and time; the rate of hydroxylation was linear for all animal species over a I-hr period. In the same way, the hydroxylation rate was found to be linear over a range of microsomal protein concen- trations of O-9 mg per incubation (data not shown).
Saturatiorl with substrate
It is of great importance when comparing activi- ties of microsomal enzymes from different animal
I I I I I I 22 42 62 62 10.4 12 4
Table I. Hydroxylation of~heptadecane br vim by liver microsomcs from direrent animal species
Species* Liver weight
(g/kg body WI)
Microsomal protein Rate of heptadecane metabolism content
(mg/g liver) (pmol/mg protein/min) (nmol/kg body wt/min)
Trout (6) 10.2 t 0.6 16.1 + 1.5 6.0 f 0.7 1.0 Rat (4) 40.1 + 2.2 24.8 f 2.1 10.1 + 0.8 10.1 Rabbit (3) 44.8 f 2.5 15.4 + 1.2 14.2 + 1.4 9.8 Chicken (4) 20.3 f 0.9 25.3 + 2.8 105.3 f 8.1 54.7
*Numbers of animals in brackets. Values are means + SD for the number or animals indicated.

Paralfin hydroxylation in liver microsomes 365
Table 2. Hepalic cytochrome P-450 dependent melabolism ol n-hepladecane
Species
Rate or heptadecane Cyrochrome P-450 corm* Apparenl i.,, metabolism
(nmol/mg protein) Inm) (pmol/nmol cyt. P-450/min)
Trout 0.19 f 0.04 450 31.74 Rat 0.52 f 0.13 450 19.48 Rabbit 0.95 + 0.21 450 14.91 Chicken 0.23 f 0.04 452 456.52
*Values are means f SD for numbers of animals indicated in Table I.
species, to use a saturating substrate concentration but not one that is so high as to decrease the precision of the comparison. Thus the dependence of heptadecane hydroxylase on substrate concentration was determined (Fig. 2).
Species differences irl [’ 4CJ/teptadecatle llydrosylatiorl rate
Using 6.2 ph+[‘4C]heptadecane, hydroxylation rates were compared between species (Table 1). It appears that great variations occur, with the chicken showing the highest ability to oxidize rl-heptadecane, on whichever basis the results are expressed (micro- somal protein or body weight).
As indicated in Table 2, the hepatic cytochrome P-450 concentration in the rat was 2.3-fold greater than in the chicken. Moreover, the I,,, of the reduced cytochrome P-450 CO-complex from chicken was at 452 nm, whereas for the other species the cytochrome P-450 A,,,,, was at 450 nm.
The results concerning heptadecane hydroxylation were recalculated and expressed in pmol heptadecane hydroxylated per nmol cytochrome P-450. The values obtained (Table 2) for trout, rat or rabbit liver were of the same order of magnitude, whereas the value for chicken was 20-fold higher than the mean for the other three species.
DISCUSSION
The metabolism of saturated hydrocarbons has been described in various animal species. The first step of the biotransformations, namely hydroxy- lation, leads to fatty alcohol. This is subsequently converted to fatty acid, which then undergoes B-oxidation. It has been established that paraffin hydroxylation is catalysed by a mixed-function oxidase involving cytochrome P-450, as shown using
the microsomal fraction of mouse liver (Kusunose et al. 1969) and of rabbit intestinal mucosa (Ichihara et al. 1981).
In this paper, we have reported on the relative ability of different animal species to oxidize a paraffin hydrocarbon. Large variations in the oxi- dation rate of n-heptadecane by liver microsomes were observed (Table 1). This is consistent with previous long-term irl uiuo studies in our laboratory. Table 3 summarizes some of these data, which are from experiments on rats (Tulliez & Bories, 1975b). trout (Cravedi & Tulliez, 1983) and chickens (J. E. Tulliez, unpublished data, 1975) fed diets in which normal paraffins were incorporated In this table, although results for the rat refer to an experiment conducted with eicosane, we have previously shown that heptadecane and eicosane show similar absorp tion and tissue retention when given to rats as single oral doses (Tulliez & Bories, 1975a). The results for the rat are from a chronic feeding study, but it has been observed that for chronic feeding of paraffins in the C,5-C24 range, the shorter chain lengths lead to higher residue levels in rat tissues (J. E. Tulliez, unpublished data, 1975). Thus, if results for the rat in Table 3 were for heptadecane, the hydrocarbon levels in the carcass or adipose tissue would have been higher.
Whereas hydrocarbon residues were only present in minute quantities in chickens, more hydrocarbon was stored in the other species and may have represented up to 6-8 y0 of the total amount ingested during the experimental period. The lower levels of hydrocarbon stored in the chicken may be related to its greater ability to oxidize hydrocarbon (Table 1).
Ichihara, Kusunose & Kusunose (1969) have shown that chick embryo microsomes have a high level of aliphatic hydrocarbon hydroxylation activity for the shorter-chain paraffin, decane. However,
Table 3. Qorage ofn-alkanes in trout. rats and chickens after long-term ingestion in the diet*
Species Alkane
Dietary Residue Residue in level in cwzass adipose tissue
(“, dry weight) (@g/g wet weight) (ppig wet weight)
Troult Hcpladccanc 0. I 1 I41 I474 RM$ Eicosenc 0.10 373 I029 Chickens Hcptadecane 0.10 Traces 24
*Feeding of lest diets in these srudies continued until the concentration of hydrocarbon in adipose tissue reached a plateau.
tSee Cravcdi & Tulliez (1983) for rull details. Dam presenred above are from the end or7 months or leeding rainbow trout 1.0”. mixed n-alkanes in the diet.
fSee Tulliez & Bories (1975b) for l’ull details. Data presented above are Tar male Wistar rats fed for 3 months with eicosane only in the diet.
(jJ. E. Tulliez. unpublished dala (1975).

366 E. F. PERDU-DURAND and J. E. TULLIEZ
Kusunose et al. (1969) found differences between the hydroxylation systems of rr-decane and II- hexadecane. Indeed, lung and kidney microsomes from mice could oxidize lower alkanes into the corresponding alcohols and fatty acids, but did not transform rt-hexadecane, whereas liver microsomes oxidized this parafin readily to palmitic acid.
Heptadecane hydroxylation has been found to remain unchanged after pretreatment with the cyto- chrome P-450 inducers phenobarbital or 3-methyl- oholanthrene (Durand & Tulliez, 1980), whereas Ichihara et al. (1969) found that decane hydroxy- lation was enhanced five- to ten-fold in rats pre- treated with phenobarbital, thus confirming the difference between the hydroxylating systems of short- and long-chain alkanes.
Moreover, the cytochrome P-450 concentrations determined in the present study are in agreement with those found in studies where enzymatic activi- ties from rat and chicken liver microsomes were compared (Lasker, Graham & Abou-Donia, 1982). They underline interesting differences in the mixed- function oxidase systems of mammalian and avian species. Firstly, the levels of constitutive cytochrome P-450 found in the chickens in our study were 2.3- and 4.1-fold less than those found in rats and rabbits, respectively. Secondly, the nature of the haemo- proteins was different since the Soret maximum of the reduced-CO binding complex of chicken liver microsomes was at 452 nm while that of rat and rabbit liver microsomes was at 450 nm.
These differences along with a 20-fold higher heptadecane hydroxylase activity in chickens than in rats or rabbits, calculated on the basis of cytochrome P-450 content, suggest that a specialized form of haemoprotein is responsible for the hydroxylation of long-chain hydrocarbons. The minor form of cyto- chrome P-450, not inducible by phenobarbital or 3- methylcholanthrene, metabolizing heptadecane in rats may be the major form in the chicken. The existence of such functionally specialized cyto- chromes has already been mentioned by Ichihara and his colleagues (Ichihara, Yamakawa, Kusunose & Kusunose, 1979; Ichihara ef al. 1981) who pointed out that alkanes, aromatic hydrocarbons and fatty acids were hydroxylated in rabbit intestinal mucosa by different cytochrome P-450 isozymes. Whether there is a true native cytochrome P-450 in chickens or a mixture of cytochromes that gives rise to the peak at 452 nm is to be determined in future investigations by us using purification procedures and reconstituted systems.
REFERENCES
Cravedi J. P. & Tulliez J. E. (1983). Hydrocarbons disposition, lipid content, and fatty acid composition in trout after long-term dietary exposure to n-alkanes. Ewir. Rcs. 32,398.
Cravedi J. P.. Tulliez J.. Choubert G. & Luquet P. (1983). Digestibilite des hydrocarbures saturis chez la Truite. Dtoir. Polltrr. Ser. A. 32, 39.
Durand E. & Tulliez J. (1980). Microsomal oxidation of linear, branched and cyclic paraffins in rat liver. Ann. Nutr. Aliment. 34. 491.
Ichihara K.. Ishihara K.. Kusunose E. & Kusunose M. (1981). Some properties of a hexadecane hydroxylation system in rabbit intestinal mucosa microsomes. J. Biachem. 89, I82 I.
Ichihara K.. Kusunose E. & Kusunose M. (1969). Micro- somal hydroxylation of decane. Biochirn. biophys. Acra 176, 713.
Ichihara K., Yamakawa I.. Kusunose E. & Kusunose M. (1979). Fatty acid w- and (m-1) hydroxylation in rabbit intestinal mucosa microsomes J. Biochetn. 86, 139.
Kolattukudy P. E. & Hankin L. (1966). Metabolism of a plant wax paraffin (n-nonacosane) in the rat. J. Nufr. 90, 167.
Kusunose M., Ichihara K. & Kusunose E. (1969). Oxidation of n-hexadecane by mouse liver microsomal fraction. Bioch. biophys. Acta 176. 679.
Ladan P. E. (1970). Intake of feed hydrocarbons and purified paraffins in chicks. Dokl. Vses. Akad. Sel’Skokho; Nauk. 4. 36.
Lasker J. M.. Graham D. G.. JL Abou-Donia M. B. (1982). Differential metabolism of O-ethyl O-4 nitrophenyl phenylphosphonothioate by rat and chicken hepatic microsomes. Biochern. Phwac. 31, 196 I.
Lowry 0. H., Rosebrough N. J., Farr A. L. & Randall R. J. (1951). Protein measurement with the Folin phenol reagent. J. biol. Chern. 193, 265.
McCarthy R. D. (1964). Mammalian metabolism of straight-chain saturated hydrocarbons. Biochim hiophys. Acta 84.74.
Mitchell M. P. & Hiibscher G. (1968). Oxidation of II- hexadecane by subcellular preparations of guinea pig small intestine. Eur. J. Biochem. 7,90.
Omura T. & Sato R. (1964). The carbon monoxide binding pigment of liver microsomes. I. Evidence for its hemo- protein nature. J. hiol. Ghan. 239,237O.
Tulliez J. & Bories G. (1975a). Metabolisme des hydro- carbures paraffiniques et naphtbniques chez les animaux suptrieurs. I. RCtenlion des paraffines (normal, cycle et ramif&) chez le rat. Am. Nufr. Alimerlf. 29, 201.
Tulliez J.-E. & Bories G.-F. (1975b). Mttabolisme des hydrocarbures paraffmiques et naphtbniques chez les animaux superieurs. II. Accumulation et mobilisation chez le rat. AWL Nrm. Aliment. 29, 213.
Tulliez J. E. & Bories G. F. (1978). Metabolism of a n- paraffin. heptadecane, in rats. Lipids 13, 1 IO.







![Multi-tissue RNA-seq and transcriptome …...where squalamine, a compound discovered in the liver of this shark species with antimicrobial activity [31, 32] has been more recently](https://img.pdfslide.fr/doc/110x75/5e665011985b9a59ee449497/multi-tissue-rna-seq-and-transcriptome-where-squalamine-a-compound-discovered.jpg)





















