Embed Size (px)
Citation preview

Ig S U R F A C E RECEPTORS A N D ERYTHROPHAGOCYTIC ACTIVITY O F HISTIOCYTOSIS X CELLS I N VITRO
CHRISTIAN NEZELOF, NICOLE DIEBOLD AND
MARIE FRANCOISE ROUSSEAU-MERCK Croupe de Pathologie Pediatrique, Inserm U 77, Hopital des Enfants Malades, 149, rue de Shres,
75730 Paris CEDEX 15
PLATES XLVI AND XLVII
COMBINING three separately described diseases (Letterer-Siwe disease, Hand-Schiiller-Christian disease and eosinophilic granuloma), Lichtenstein (1953) coined the name " histiocytosis X ", to stress the probably histiocytic nature of the cellular proliferation and the mysterious origin of the disease.
Since this initial description, several new facts have come to light which are relevant to the identification of the principal proliferating cell of this disease. The most important contributions are: the discovery within the cytoplasm of most HX cells of a pentalaminar ultrastructural marker, the X body (Basset and Turiaf, 1965; Basset and Nezelof, 1966; De Man, 1968). Although similar, if not identical granules have been described in the Langerhans cells of the epidermis (Breathnach, 1969, comparable structures have rarely been observed in the commonly recognised histiocytic cells.
The classical concept of the reticulo-endothelial system has become un- satisfactory and has been replaced by the mononuclear phagocyte system (MPS), which has been delineated according to the origin, morphology, kinetics and immunological function of its constituent cells (Van Furth et al., 1972).
It is justified therefore to investigate the immunological behaviour of HX cells and to compare it with that of cells of the MPS. Since humoral-dependent avid phagocytic activity is regarded as the best functional criterion of MPS cells (Van Furth et al., 1972), it was decided to study the surface receptors of HX cells and to explore their phagocytic activity in tissue culture.
MATERIALS AND METHODS
The abbreviations used are: HX Histiocytosis X MPS Mononuclear phagocyte system HBSS Hank's balanced salt solution FITC Ig Fluorescein isothiocyanate conjugated goat
anti-human immunoglobulin FITC a, p , y Fluorescein isothiocyanate conjugated goat anti-
human immunoglobulin heavy chains FITC C3 Fluorescein isothiocyanate conjugated rabbit antLC3 RBC Red blood cells EAC Sheep erythrocytes coated with antibody and
SIg Surface immunoglobulin receptors Surface C3 receptors sc3
complement
-. -~ __ Received 10 Oct. 1916; accepted 3 Nov. 1976.
1. PATH.-VOL. 122 (1977) 105

106 CHRISTIAN NEZELOF, NICOLE DIEBOLD, MARIE F. ROUSSEAU-MERCK
The material was obtained from three cases of histologically confirmed eosinophilic granuloma of the skull. Pieces selected from the pink areas of the tumours removed in the operating room were gently minced with scissors. Some of the resulting fragments were suspended in HBSS and trypsinised (0.25 per cent. Trypsin, Flobio, Courbevoie, France) at 37°C for 10 min.
The cellular suspension was collected and washed twice in HBSS, then centrifuged at 800 r.p.m. and re-suspended in Eagle’s nutrient medium (Institut Pasteur, Paris) supplemented with 10 per cent. calf serum and antibiotics (penicillin, 200 unitslml; streptomycin, 0.1 mglml). The cells were seeded in Leighton tubes and allowed to settle and to grow. The nutrient medium was changed twice a week. A number of approximately 1 mm3 fragments were explanted in Leighton tubes and allowed to grow under the same conditions as the suspensions and used between the 5th and 12th days of culture.
The following studies were performed on the 5th, 6th and 8th days of culture.
Challenge with serum and with fluorescein-conjugated antihuman immunoglobulin (FITC-Ig) The nutrient medium was removed and the cultures on the coverslips were rinsed three
times with HBSS for 10 min. Then the cells were incubated at room temperature for 30 min. with patient’s serum or AB serum obtained from the blood bank in a dilution of 10 per cent. The final volume of culture media was always 2 ml and the final concentration of the human serum 10 or 20 per cent. Control coverslips with cultures were incubated in HBSS under the same conditions. The incubated cells were washed three times with HBSS for 10 min. before being exposed at room temperature for 30 min. to the 1/10 and 1/4 diluted fluorescein- conjugated goat anti-human serums (FITC Ig, IgG, IgM, IgA) obtained from the Miles Laboratory, Kankakee, Ill. In one case, a fluorescein-conjugated rabbit anti C3 (FITC-Cd serum (Institut Pasteur, Paris) was also used.
After three further washings with HBSS, the cells were mounted in a mixture of phosphate buffered isotonic saline solution and glycerin on glass slides without fixation and were immediately examined under a Zeiss UV microscope using epi-illumination.
Challenge with human RBC andsheep erythrocytes coated with antibody and complement ( E A o After changing the nutrient medium, the coverslips with the cultured cells were incubated
at 37” for 2 hr with a 1 to 6 per cent. HBSS suspension of either anti RhD-treated Rht- human RBC or sheep RBC coated with rabbit anti-sheep RBC+complement components (EAC cells) obtained from the Institut Pasteur, Paris. After being washed with HBSS three times, the cultures were fixed either in aIcoholic Bouin’s solution for light-microscopic study or in a 3 per cent. buffered aqueous glutaraldehyde solution for electron-microscopic study. Control studies using non-opsonised human or sheep erythrocytes were undertaken using the same methodology as with the experimental material.
Challenge with latex particles 0.8 pm latex particles (Difco, Detroit, Mich.) were employed with or without opsonisation
with patient’s serum diluted to 10 per cent. in Eagle’s medium. After being washed with HBSS, the specimens were fixed in an alcoholic Bouin’s solution, stained with H t E and studied in the usual manner.
RESULTS
Tissue culture AS
described previously (Basset and Nezelof, 1969; Nezelof et al., 1973), it was heterogeneous and included :
The cellular growth was visible from the 4th-5th day of culture.

IMMUNOLOGICAL ACTIVITY OF H X CELLS 107
(a) A large number of round-shaped, adherent cells, ranging in diameter from 8 pm to 13 pm. These were regarded as the surviving HX cells since they had the same notched and scalloped nuclear outline, identical histochemical equipment and the same ultrastructural marker (fig. 1) as HX cells in vivo. They constituted the great majority of the attached cellular population at the early stage of culture.
(b) Large polykaryocytes measuring from 25 pm to 200 pm, having 5 to 100 nuclei. Although they did not contain any cytoplasmic X bodies, these polykaryons presumably resulted from the fusion of the above described HX cells (McAllister et al., 1963; Basset and Nezelof, 1969). Their proportion increased from the 6th or 7th day and decreased after the 12th day (Basset and Nezelof, 1969; Nezelof et al., 1974). Coarse lipochrome pigments were often present in the cytoplasm of these cells (Nezelof et al., 1974).
(c) Spindle-shaped cells ; these had the morphological appearance of fibroblasts and were thought to originate from the stroma of the eosinophilic granulomas. Their number increased with the duration of the culture and they provided a useful control for comparing the in-vitro behaviour of the HX cells with these cells.
(d) A few round-shaped cells; they had a smooth, oval nucleus. They were regarded as normal macrophages originating from the stroma of the tumours and were fairly easily distinguished from the HX cells even with the light microscope because of the deeply notched nucleus of the latter. No X bodies were seen in these cells with the electron microscope.
Immunojluorescence studies After incubation in HBSS, no fluorescence could be detected on the surface
of the cells. However, some yellow autofluorescent granules were noticed within the cytoplasm of the HX cells. These spontaneously fluorescent granules were coarse and irregularly distributed in the cytoplasm of some of the large HX cells. These granules appeared to be the accumulated lipochrome pigment which is known to be abundant in these cells (Nezelof et al., 1974). As this pigment was absent from the fibroblasts it constituted a good indicator for the identification of HX cells in the membrane fluorescence studies.
After incubation in either patient’s serum or in AB serum, a fine spotty fluorescence was observed over the surface of the HX cells after reaction with FITC-Ig and FITC-IgG. This membrane fluorescence was seen on 20 per cent. to 40 per cent. of the HX cells and seemed to be unequally distributed over the cell surface (fig. 2). No surface fluorescence was detected on polykaryocytes and fibroblasts. FITC-IgA and FITC-IgM and, in the one case studied, FITC-C3 serum also produced negative results (table I).
The fluorescence of the surface immunoglobulins was similar, regardless whether the cultures were incubated with patient’s or with AB serum, or of the various dilutions used. Due to the shortage of pathological material the extinction reactions with unconjugated sera were not performed.

108 CHRISTIAN NEZELOF, NICOLE DIEBOLD, MARIE F. RO USSEAU-MERCK
Attempts to fix the cells in alcohol or acetone prior to incubation with FITC sera resulted in a diffuse, misleading fluorescence.
TABLE I Membrane fluorescence of H X cells and control fibroblasts
HBSS Patient's serum AB serum Incubation r A
FITC k @ Y Y & @ Y c3 HX cells ' , g ; K 6 + o o + + o K + o Fibroblasts 0 0 0 0 0 0 0 0 0 0 0 0 0
+ = fluorescence. 0 = no fluorescence.
Phagocytic studies When anti RhD-treated Rh+ human RBC and EAC cells were used an
intense erythrophagocytosis was readily demonstrated. The erythrophago- cytosis was preceded by conspicuous rosette formation (fig. 3). The erythro- phagocytic activity involved only the HX cells and especially mononucleate HX cells: some are able to engulf ten and even more RBC after 2 hr of incubation (fig. 4). The erythrophagocytic activity of the large HX cells appeared to be comparatively lower and the polykaryocytes did not ingest RBC. The number of normal " bystander " macrophages was very small and evidence of erythro- phagocytosis within them was not obvious.
The erythrophagocytic activity could be seen in 40 per cent. to 60 per cent. of the glass-adherent cells of good cultures. The omission of opsonin led to negative results in all the experiments.
The use of latex particles gave similar results. This material which, probably on account of its smaller size (0-8 pm), could be more easily engulfed, was present not only in the small HX cells but also in some of the large HX cells and in the polykaryocytes. The presence of opsonins was not a prerequisite to demonstrate phagocytosis of latex particles by the small and avid HX cells (table 11).
TABLE I1 Phagocytic activity of H X cells, polykaryocytes and control jibroblasts
ORh+ human RBC Latex particles &&
Ops. Not ops. Ops. Not ops. HX cells + 0 + + Polykaryocytes 0 0 + 0 Fibroblasts 0 0 0 0
Ops. = opsonised. Not ops. = not opsonised. + = phagocytosis. 0 = no phagocytosis.
DISCUSSION The aim of the present study was to investigate the immunological behaviour
of living HX cells and particularly to see whether the HX cells bear immuno- logical surface receptors and whether they are able to phagocytose foreign particles under precisely defined conditions.
NEZELOF. DIEBOLD AND ROUSSEAU-MERCK PLATE XLVI
IMMUNOLOGICAL ACTIVITY OF HX CELLS
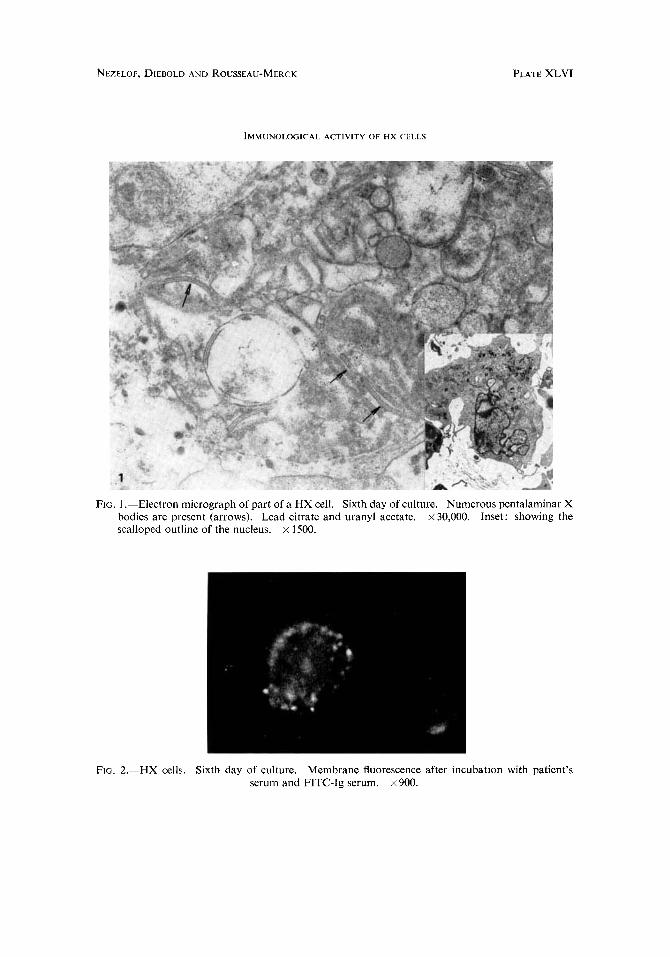
FIG. 1 .-Electron micrograph of part of a HX cell. Sixth day of culture. Numerous pentalaminar X bodies are present (arrows). Lead citrate and uranyl acetate. x 30,000. Inset: showing the scalloped outline of the nucleus. x 1500.
FIG. 2.-HX cells. Sixth day of culture. Membrane fluorescence after incubation with patient’s serum and FITC-Ig serum. x 900.
NEZELOF, DIEBOLD AND ROUSSEAU-MERCK
IMMUNOLOGICAL ACTIVITY OF HX CELLS
PLATE XLVII
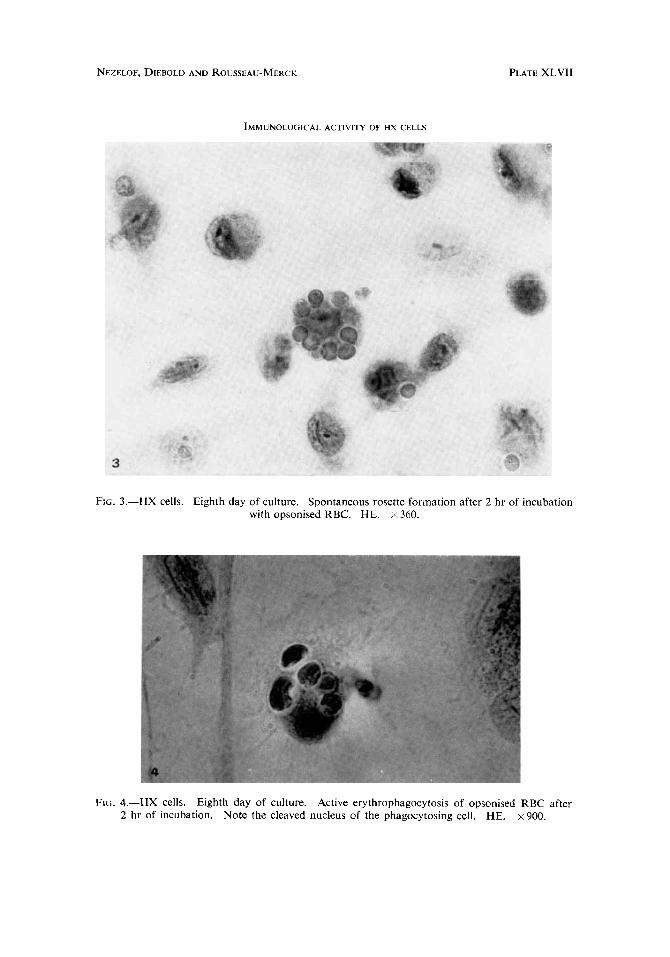
Frti. 3.-HX cells. Eighth day of culture. Spontaneous rosette formation after 2 hr of incubation with opsonised RBC. HE. x 360.
FIG. 4.-HX cells. Eighth day of culture. Active erythrophagocytosis of opsonised RBC after 2 hr of incubation. Note the cleaved nucleus of the phagocytosing cell. HE. x 900.

IMMUNOLOGICAL ACTIVITY OF H X CELLS 109
Tissue culture morphology As has already been shown HX cells can be maintained in tissue culture for
several weeks (McAllister et al., 1963; Basset and Nezelof, 1969; Nezelof et al., 1974). Although they do not divide in vitro under the conditions employed in those studies, the HX cells retain their shape, their notched or scalloped nucleus, their cytochemical equipment and their ultrastructural marker (Nezelof et al., 1973). The use of 5-7-day-old tissue cultures of eosinophilic granuloma offers especially valuable material in this respect in the study of living HX cells. At this stage the eosinophil leucocytes, the plasma cells and the lymphocytes have disappeared and only some fibroblasts and macrophages are growing among the HX cells. The fibroblasts, incidentally, provide useful controls for comparing the morphological, kinetic and immunological characteristics of these two categories of cells (Nezelof et al., 1973). In addition, large polykaryocytes, probably resulting from the fusion of HX cells, are an almost constant finding at this stage of the culture.
In the present study, the majority of the growing cells retained the main features of HX cells. Due to the tight adherence of the HX cells to the glass coverslips, difficulties were encountered in attempting to harvest intact phago- cytising HX cells for proper electron-microscopic studies. Nevertheless we have demonstrated the presence of the pentalaminar X body in these cells in electron- microscopic studies of phagocytosing cultures (fig. 1). The X bodies were only found in cytoplasmic areas where no evidence of phagocytic activity was seen, suggesting that phagocytosis is associated with the disappearance of these membrane related ultrastructural markers (Basset and Nezelof, 1969). This does not disprove the true HX nature of the phagocytic cell since it is known that the X body is a provisional structure which is present only in the early stages of the differentiation of this type of cell and which disappears in the older forms, especially in polykaryocytes (Nezelof et al., 1973). In the absence of this peculiar cytoplasmic marker, the deeply indented or scalloped nucleus allowed us in most cases to recognise the true nature of the individual phagocytic cell and to distinguish it from stromal " bystander "macrophages.
Cytoplasmic autofluorescent lipochrome pigment was most abundant in old cells, especially in polykaryocytes. Such material, although in lesser quantities, was visible in the HX cells as well as ordinary macrophages. Thus the presence of this pigment, except for its absence in fibroblasts, was of limited use in differentiating the various cell types within the cultures.
Immunological surface receptors It is now well documented that monocytes and macrophages carry numerous
surface receptors and are able, under certain conditions, to bind IgG and the C3 fraction of complement on to receptors (SIg or SC3) on their surface (Berken and Benacerraf, 1966; Huber and Fudenberg, 1967; Huber and Fudenberg, 1968 ; Lay and Nussenzweig, 1968). It is also well established that the phagocytic activity of these cells is closely dependent on the co-operation of certain serum

110 CHRISTIAN NEZELOF, NICOLE DIEBOLD, MARIE F. ROUSSEAU-MERCK
factors (Ig, complement, etc) and, for this reason, is regarded as an immuno- phagocytosis. This distinguishes “ professional phagocytes ” from non- professional or facultative phagocytes that can ingest particles at low rates without dependence on immunoglobulins and complement (Rabinovitch, 1967; Rabinovitch, 1968 ; Van Furth and Thompson, 1971). Accurate evaluation of such functional capacities of various cell types have provided valuable criteria in determining whether they belong to the MPS or not.
In our study, incubation of HX cells either in patients’ serum or in AB serum with the simultaneous use of FITC-IgG revealed SIg on the plasma membrane of the HX cells (fig. 2). These SIg appear as a continuous or discontinuous punctuate ring similar, if not identical, to the S-immunofluorescence pattern observed in the case of monocytes and macrophages after exposure to various blood serums (Unanue, 1972). An irregular distribution of SIg on the surface of the HX cells is frequently observed and may be related to the fact that these cells are adherent to glass and are mobile. SIg can be detected on only 20 to 40 per cent. of HX cells. These cells are usually small and round-shaped. Few SIg are visible on the larger cells and none on the polykaryocytes. The reasons for this peculiar distribution are not clear, although one may assume that the large HX cells and, even more likely, the polykaryocytes represent inactive, old elements (Papadimitriou, 1973 ; Mariano and Spector, 1974). Incubation with either patient’s serum or with AB serum produces similar results. Only receptors for IgG can be demonstrated. The use of specific FITC-IgA and FITC-IgM, under the same conditions, fails to reveal any receptor for those kinds of immuno- globulins. It is known, indeed, that only IgG (and more specifically IgGl and IgG3) are cytophilic for the human macrophage (Huber and Fudenberg, 1968).
No C3 receptor has been observed on the surface of HX cells in direct immunofluorescence, whereas C3 receptors are known to be present on the surface of monocytes and macrophages (Huber and Fudenberg, 1968; Lay and Nussenzweig, 1968). This negative result remains questionable since this investigation has been carried out only once and under difficult technical conditions. The rosettes obtained with the use of human RBC coated with antigen-antibody complexes or EAC cells, including activated fractions of complement, lead one to think that HX cells also bear receptors for the C3 fraction of complement.
Phagocytic activity Much more than the presence of SIgG and SC3 avid phagocytic activity
and especially humoral-dependent immunophagocytic activity have been considered as the most reliable functional criteria to identify cells belonging to the MPS (Rabinovitch, 1967; Van Furth, 1970).
In our experiments, the phagocytosis of heterologous and opsonised RBC is closely related to the presence of antigen-antibody complexes on the surface of RBC, since erythrophagocytosis is suppressed if the RBC are not sensitised by antibodies prior to performing the experiment (table 11). It is important to stress that the small HX cells show a greater erythrophagocytic activity than the large ones. Some of these small cells can engulf six to ten and sometimes more

IMMUNOLOGICAL ACTIVITY OF H X CELLS 111
RBC (fig. 4). However, the polykaryocytes, like the fibroblasts, are not able to take up any RBC into their cytoplasm. No rosette formation can be seen around these multinucleated cells, suggesting that these elements are poorly active and carry few membrane-receptors. Nevertheless, the use of smaller foreign particles, i.e., 0.8 pm latex particles, makes it possible to observe the potential phagocytic activity of some polykaryocytes. For this latter type of phagocytosis opsonins provided by preliminary incubation in pooled human serum are not required since some small HX cells are also able to engulf unopsonised latex particles.
Under the conditions of this study the number of cells which showed erythrophagocytic activity was greater than the number which showed mem- brane immunofluorescence. We have no explanation for the quantitative discrepancy between these results. The phagocytosis obtained in HX cultures can be readily correlated with the frequent evidence of phagocytosis (RBC, cellular debris or eosinophilic granules) observed on routine histological examination of eosinophilic granulomas.
The results obtained in this study corroborate the data obtained in our previous studies, in which glass adherence, mobility, tendency to form poly- karyocytes, absence of DNA synthesis in vitro and the type of cytochemical equipment (i.e., acid phosphatase and esterase) of the HX cells were similar to those found by studying other " professional " mononuclear phagocytes (Basset and Nezelof, 1969; Nezelof et al., 1974). Our results and the data obtained by others provide a consistent group of arguments strongly suggesting that the HX cell represents the pathological (probably neoplastic) variant of a cell which normally belongs to the MPS and, aposteriovi, justifies the name " histiocytosis " given 20 yr ago to this disease.
Since the HX cell shares with the epidermal Langerhans cell numerous morphologic features: size, scalloped nucleus and the presence of an ultra- structural marker in their cytoplasm called either X body (Nezelof et al., 1973) or Langerhans Body (Breathnach, 1965), one may assume that the poorly understood, clear, dendritic Langerhans cells constantly present in the epidermis (Breathnach, 1965) and those occasionally found in lymph nodes (Kondo, 1969) or in the thymus (Hoshino, 1970) also belong to the mononuclear phagocyte system, and may constitute the stem cell of histiocytosis X (Nezelof et al., 1973).
SUMMARY
Membrane receptors for Ig and IgG and receptors for immune complexes (C3) have been identified on the surface of histiocytosis X cells by direct immuno- fluorescence and by rosette formation. Histiocytosis X cells also avidly phagocytose human RBC and latex particles. The cells studied originated from three eosinophilic granulomas of bone and were maintained in culture for 12 days.
These immunological characteristics suggest that histiocytosis X cells represent a neoplastic variant of a cellular subpopulation (Langerhans lineage) which probably belongs to the mononuclear phagocyte system.

112 CHRISTIAN NEZELOF, NICOLE DIEBOLD, MARIE F. ROUSSEAU-MERCK
RBsmB Des rkcepteurs de membrane pour les Ig, les IgG, ainsi que pour les com-
plexes immuns (C,) ont CtC identifiCs a la surface de cellules d'histiocytose X, provenant de 3 granulomes Cosinophiles osseux et maintenus en culture. Confronttes a des particules de latex et A des hCmaties humaines opsonisies, ces cellules ont Cgalement montrC une intense activiti phagocytaire.
Ce comportement immunologique, joint aux particularitks ultrastructurales suggttre que les cellules de l'histiocytose X reprksentent la dkviation tumorale d'une lignCe cellulaire originale (lignCe langerhansienne) laquelle appartient probablement au systttme phagocytaire mononuclCC.
Key words : histiocytosis X; Langerhans cell; mononuclear phagocyte system; erythro- phagocytosis; IgG membrane receptors.
The authors express their gratitude to Dr F. Basset and Dr R. Van Furth for their help and guidance. We wish also to thank P. Bolo for providing opsonised RBC, J. J. Prat and H. Mouly for their skilful technical assistance and Mrs Grimal for preparation of the manuscript.
REFERENCES BASSET, F., AND TURIAF, J. 1965. Identification par la microscopie Clectronique de particules
de nature probablement virale dam les lksions granulomateuses d'une Histiocytose X pulmonaire. C.R. Acad. Sci (Paris), 261, 3701-3703.
Presence en microscopie electronique de structures filamenteuses originales dans les lesions pulmonaires et osseuses de l'histiocytose X. Etat actuel de la question. Bull. Mem. SOC. Med. Hap. Paris, 117, 413426.
BASSET, F., ET NEZELOF, C. 1969. L'Histiocytose X. Microscopie Clectronique, culture in vitro et histoenzymologie. Discussion A propos de 21 cas. Rev. Franc. Etud. Clin. Biol., 14, 31-45.
BERKEN, A., AND BENACERRAF, B. 1966. Properties of antibodies cytophilic for macrophages. J. Exp. Med., 123, 119-144.
BREATHNACH, A. S. 1965. The cell of Langerhans. International Review ofcytology, 18,l-28. DE MAN, J. C. H. 1968. Rod like tubular structures in the cytoplasm of the histiocytes in
histiocytosis X. J. Path. Bact., 95, 123-126. HOSHINO, T., KUKITA, A., AND SATO, S. 1970. Cells containing Birbeck granules (Langerhans
cell granules) in the human thymus. J. Elect. Microsc., 19, 271-276. HUBER, H., AND FUDENBERG, H. H. 1967. In vitro interaction of human monocytes with
IgG and red cells sensitised with different IgG antibodies. Blood, 30, 885-886. HUBER, H., AND FUDENBERG, H. H. 1968. Receptor sites of human monocytes for IgG.
In?. Archs. Allergy, App. Immun., 34, 18-31. KONDO, Y. 1969. Macrophages containing Langerhans cell granules in normal lymph nodes
in rabbit. Z . Zellfousch, 98, 506-51 1 . LAY, V. H., AND NUSSENZWEIG, V. 1968. Receptors for complement on leukocytes. J. Exp.
Med., 128, 991-1007. LICHTENSTEIN, L. 1953. Histiocytosis X. Integration of eosinophilic granuloma of bone,
Letterer Siwe disease and Hand Schuller Christian disease as related manifestations of a single nosologic entity. Arch. Path., 56, 84-102.
MCALLISTER, R. M., KIKENAS, M., SCHAW, R. M., AND LANDING, B. 1963. Cytologic and virologic studies of culture derived from neoplastic and non-neoplastic tissues in children. Lab. Invest., 12, 343-354.
MARIANO, M., AND SPECTOR, W. G. 1974. The formation and properties of macrophage polykaryons (inflammatory giant cells). J. Path., 113, 1-19.
BASSET, F., ET NEZELOF, C. 1966.

IMMUNOLOGICAL ACTIVITY OF HX CELLS 113
NEZELOF, C., BASSET, F., AND ROUSSEAU, M. F. 1973. Histiocytosis X. Histogenetic arguments for a Langerhans cell origin. Biomedicine, 18, 365-371.
NEZELOF, C., DESPRES, S., BARBEY, S., ET ROUSSEAU, M. F. 1974. Histiocytose X. Elbments histochimiques et autoradiographiques en faveur d’une synthese lipidique en culture in vitro. Biomedicine, 20, 414-424.
PAPADIMITRIOU, J. M. 1973. Detection of macrophage receptors for heterologous IgG by scanning and transmission electron microscopy. J. Path., 110, 213-219.
RABINOVITCH, M. 1967. Non-professional and professional phagocytosis : Particle uptake by L cells and by macrophages. J. Cell Biol., 35, 1108a (abstract).
RABINOVITCH, M. 1968. Phagocytosis: the engulfment stage. Sem. in Hemat., 5, no 2, 134-155.
RABINOVITCH, M. 1970. Phagocytic recognition. Zn Mononuclear phagocytes, edited by R. Van Furth, Blackwell, Oxford, p. 299.
UNANUE, E. R. 1972. The regulatory role of macrophages in antigenic stimulation. I n Advances in immunology, 15, 134-165.
VAN FURTH, R. 1970. The origin and turnover of promonocytes, monocytes and macrophages in normal mice. In Mononuclear phagocytes, edited by R. Van Furth, Blackwell, Oxford, p. 151.
VAN FURTH, R., COHN, Z. A., HIRSCH, J. G., HUMPHREY, J. H., SPECTOR, W. G., AND LANGEWORT, H. L. 1972. Le systeme phagocytaire mononuclbaire: nouvelle classifica- tion des macrophages, des monocytes et de leurs cellules souches. Bull. Org. Mond. SantP, 47, 651-655.
VAN FURTH, R., AND THOMPSON, J. 1971. Review of the origin and kinetics of the promono- cytes, monocytes and macrophages and a brief discussion of the mononuclear phagocyte system. Ann. Znst. Pasteur, 120, 337-355.