Embed Size (px)
Citation preview

Eur. J. Biochem. 133, 135- 139 (1983) i FEBS 1983
Inactivation-Reactivation Process and Repression of Permease Formation Regulate Several Ammonia-Sensitive Permeases in the Yeast Saccharomyces cerevisiae
Marcelle GRENSON
Laboratoire de Microbiologie, Faculte des Sciences, Universile libre de Bruxelles; and Institut de Recherches du Centre d’Enseignement et de Recherches des Industries Alimentaires et Chimiques, Bruxelles
(Received December 21, 1982) - ETB 6354
Two distinct regulatory mechanisms are responsible for the absence of general amino-acid perinease activity in cells of the wild-type strain Z1278b of Succhurornyces cerevisiue grown in the presence of ammonium ions.
One is a reversible inactivation process which progressively develops upon addition of ammonium ions to a proline-grown culture, and completely suppresses the permease activity within onc hour. This inactivation process is absent in mutants altered at the MUT2, MUT4, or PGR genetic loci.
I n these mutants, a repression of the formation of active permease may clearly be observed in the presence of ammonium ions. This second regulatory mechanism is absent in mutants affected at the GDHCR locus, which might code for a repressor molecule. I t is also relieved in the presence of a gln‘& mutation (which makes the glutamine synthetase thermosensitive) suggesting glutamine as an effector.
Two other ammonia-sensitive permeases, namely the proline permease and the ureidosuccinic-acid permease, seem to be subject to the same double regulation.
Mutations affecting the structural gene of the anabolic NADP-linked glutamate dehydrogenase (gdhA) seem to completely prevent repression of the general amino-acid permease, while they partially suppress its inactivation in the presence of ammonium ions.
Several uptake systems which transport potential nitrogen sources in the yeast Sacchuromyces cerevisiue are not active in cells grown in the presence of ammonium ions [l - 61. Based on the fact that they support the highest growth rates in minimal medium, ammonium ions and glutamine may be considered as the best nitrogen sources for this yeast. The mechanisms by which they prevent a number of other nitrogen compounds from being utilized have not been elucidated as yet. The same is true for other fungi, and even in bacterial cells many questions in the field of nitrogen catabolite repression remain obscure. The importance of nitrogen catabolite repression for the cell economy as well as for the understanding of fundamental regulatory mechanisms justifies new attempts at elucidating these problems. Conditions seem favourable in the case of the yeast general amino-acid permease, with the additional interest that constituents of the plasma membrane are concerned.
Several mutations are known to affect nitrogen catabolite inhibition or repression of permeases and enzymes in yeast. They have, however, provided little information about the mechanisms involved. The gdhA mutations, located in the structural gene for the anabolic NADP-linked glutamate dehydrogenase 171, decrease or suppress the sensitivity of these uptake systems [3,5,6], as well as that of nitrogen catabolic enzymes of S. cerevisiue [8 - 101, to the ‘ammonia effect’. The interest of these gdhA mutations is that their consequences do not seem to be simply linked to a decrease in glutamate production in several systems, namely the general amino-acid permease [3], arginase [8- 101, urea amidolyase and allan-
Emyme. Glutamate dehydrogenase (NAD’) (EC 1.4.1.2)
toinase [8], and one of the glutamic-acid permeases [5,11]. Hence they might help identify regulatory molecules or effec- tors of nitrogen catabolism. A possibility which has been considered in that field is that the anabolic glutamate- dehydrogenase molecule itself might act as a regulatory protein [3,9]. The effect of gdhA mutations on the regulation of nitrogen metabolism was studied in other fungi as well [12,13].
Mutations at the GDHCR locus of S. cerevisiue suppress nitrogen catabolite repression of all the nitrogen catabolic enzymes tested, namely the catabolic NAD-linked glutamate dehydrogenase [7,14], arginase and glutamine synthetase [I 51, allantoinase and urea amidolyase [lo], and proteinaseB [I6]. Furthermore, most of the ammonia-sensitive permeases are active in ammonia-grown g d K R mutants, including the ureidosuccinic-acid permease [4, 17, 181, the repressible glutamic-acid permease [I I], the proline permease (G. S olbu, C. Casteleyn, and M. Grenson, unpublished), and two methylamine/ammonia uptake systems [6]. The gdhCR mu- tations might affect a gene coding for a general ‘nitrogen repressor’ [I 51. The reasons why the general amino-acid permease is not active in ammonia-grown gdhCR mutants will be anaiysed in the present paper. I t must be mentioned that an additional effect of the g d K R mutation has been-observed, namely the suppression of a regulatory mechanism by which glutamine inactivates glutamine synthetase [16]. Although the relationship of this last effect with the suppression of the nitrogen catabolite repression by the gdhCR mutation has not been understood as yet, this observation must be kept in mind.
Another mutation which makes the general amino-acid permease partially active is ammonia-grown cells is mepl [6].

136
This is not surprising since the mepl mutation slows down ammonia uptake by inactivating the high capacity methyl- amine/ammonia uptake system [6].
Recently, several other mutations have been isolated which affect the regulation of the general amino-acid permease [I 9,201. Their effect will be analysed in the present paper and in the accompanying study [21]. Evidence will be provided for the existence of two distinct regulatory mechanisms each of which very efficiently prevents the permease from functioning in ammonia-grown cells.
MATERIALS AND METHODS
The yeast strains used are all isogenic to the wild-type strain C 1278b of Saccharomyces cerevisiae [22] except for the mu- tations mentioned. The strains 4324d (gdhA-1), MG1694 (gdhA-I, amal ) and 12597a (gdhCR-I) have been described previously [7], as well as thegapl-1 mutant strain 2512c [2]. The mutant strain 17251a (glnts) has a thermosensitive glutamine synthetase; at 36°C it is unable to grow in the absence of glutamine, while it grows normally at 22°C; at 29"C, its doubling time in ammonia-minimal medium is 2 h 30 min instead of 2 h for the wild-type strain. This mutant has been used in a study of the methylamine/amnionia uptake systems [6]. The isolation of the mut2, mut l , andpgr mutations has been described [20]. These mutations have been utilized previously
Media and growth conditions were those described pre- viously [23]; the growth temperature was 29 "C ; glucose was the carbon source; the nitrogen source was either ammonia as 0.01 M (NH4)$04 (ammonia minimal medium, previously named M. am medium) or 1 mg x ml-' L-proline (proline minimal medium, previously named M. pro medium).
The initial uptake rate of amino acids or ureidosuccinic acid was measured as previously [24], with 0.02 mM carbamoyl-~-['~C]citrulline, 0.02 mM uniformly 14C-labelled L-proline, or 0.05 mM DL-ureidosuccinic acid. Citrulline and proline were purchased from the Radiochemical Centre, Amersham, and ureidosuccinic acid was a kind gift from Dr F. Lacroute.
The activity of NAD-glutamate dehydrogenase was mea- sured according to [25].
The pools of free amino acids were extracted from washed mid-log phase cells by ice-cold 5 trichloracetic acid for 20min; they were analysed with a Beckman amino acid analyzer (model 120c).
~ 9 1 .
RESULTS
N o general amino-acid permease activity is detectable in ammonia-grown cells of the wild-type strain C1278b of Saccharomyces cerevisiae. On the contrary, maximal activity of this permease is observed in the same yeast strain growing exponentially in proline-minimal medium [2].
Reversible Inactivation of the General Amino-Acid Permease upon Addition of Ammonium Ions to Proline-Grown Cells
The effect of adding ammonium ions to proline-grown cells is shown on Fig. 1. As observed earlier [2], upon addition of ammonia, the pre-existing general amino-acid permease ac- tivity progressively disappears so as to be completely lost within 1 h. Fig. 1 shows that this inactivation is reversible upon return to proline medium; the reactivation is rapid and
w Y Q
0 3 c
0 0.1 0 2 0.3 O.L 0.5
Absorbance
Fig. 1. Reversible inacrivution of the generul amino-acid permease upon udditiun uf ammonium ions to proline-grown wild-rype cells. The general amino-acid permease activity was estimated by measuring the initial uptake rate of 0.02 mM ~-['~C]citrulline. Wild-type cells of strain C 1278b growing exponentially in proline minimal medium (o----o). At the time shown by the arrow (i), ammonium ions [as 0.01 M (NH4),S04 final concentration] were added to a portion of the culture (H --*). After growth in the presence of ammonium ions, the cells were transferred to proline minimal medium (r), and the restoration of citrulline uptake activity was followed: (0- - recovery after 1 h in the presence of ammonium ions; (L,----L>) recovery after 3 h in the presence of ammonium ions. Abscissa: absorbance at 660 nm, as a measure of cell mass/ml in the growing culture
complete. After a longer period of growth in the presence of ammonia (3 h), it can be seen that only the permease activity present prior to ammonia addition is recovered rapidly. Further increase in general amino-acid permease activity occurs more slowly.
Absence of Inactivation Process in Mutants Altered at the MUT2, MUT4, or PGR Loci
Upon addition of ammonium ions to a mut2, mut4, or pgr mutant strain growing in proline-minimal medium, no in- activation is observed (Fig. 2a - c). The pre-existing permease activity is maintained, whereas further increase is prevented.
On the contrary, normal inactivation of the general amino- acid permease does occur in a gdhCR mutant strain, as well as in a glnts mutant strain (Fig. 2d).
Repression of Synthesis of the General Amino-Acid Permease
Repression of a constituent of the general amino-acid permease is manifest in mutants which have lost the in- activation mechanism, namely the mut2, mut4, and pgr strains (Fig. 2a-c).
Loss of Ammonia Repression in gdhCR and glntS Mutant Strains
While the activity of the general amino-acid permease is low or negligible in ammonia-grown single mut2, or mut4, or pgr mutant strains, it is high in double mutants which contain a gdhCR or a gln'" mutation (Table 1). Hence the gdhCR and the glnfA mutations provoke derepression of the general amino-acid permease. The absence of cumulative effect of other pairs of mutations is also shown in Table 1 .
Suppression qf the 'Ammonia Ej'ect' on Other Ammonia-Sensitive Permeases
Table1 shows that the same pairs of mutations which relieve the 'ammonia effect' on the general amino-acid per-
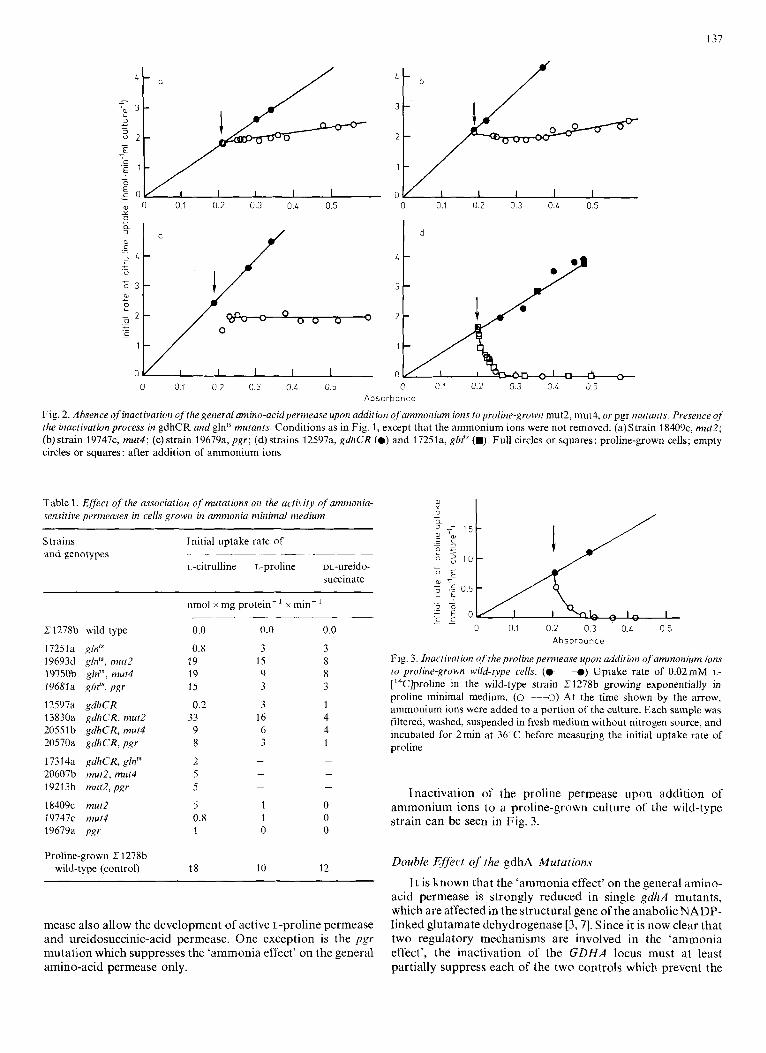
137
0 0.1 0.2 0.3 0.L 0.5 0 0.1 0.2 0.3 0.L 0.5
I d
0 0.1 0.2 0.3 0.1 0.5 0 0 1 0.2 0 3 0.L 0.5 Absorbance
Fig. 2. Absence of inactivation ojrhe general amino-acidpermeuse upon addition ofammonium ions toproline-grown mut2, mut4, or pgr mutants. Presence of the inactivation process in gdhCR and glntS mutants. Conditions as in Fig. 1, except that the ammonium ions were not removed. (a)Strain 18409c, mut2; (b) strain 19747c, m u t l ; (c) strain 19679~1, pgr; (d) strains 12597a, gdhCR (0) and 17251a, glnth (m). Full circles or squares: proline-grown cells; empty circles or squares: after addition of ammonium ions
m Y 0
D
Table 1. Efyect of the association of mutations on the activity of ammonia- .sensirive permeuses in cells grown in ammonia minimal medium c /
Strains Initial uptake rate of and genotypes _ _ _ . ~ _ _ _ .
L-citruhe r.-proline oL-ureido- succinate
nmol x mg protein-' x min-' I -- -
Cl278b wild type 0.0 0.0 0.0 0 0.1 0.2 0.3 0.L 0.5 ..
17251a glntS 19693d gin", mut2 19750b g W , mutl 19681 a ghP, pgr
12597a gdhCR 13830a gdhCR, mur2 20551b gdhCR, mut4 20570a gdhCR, pgr
0.8 19 I9 15
0.2 33 9 8
3 15 9 3
3 16 6 3
Absorbonce
Fig. 3. Inactivation of the proline permease upon addition oJammonium ions ro proline-grown wild-rype cells. (.---a) Uptake rate of 0.02 mM L-
['4C]proline in the wild-type strain ,E 127% growing exponentially in proline minimal medium. (o---*) At the time shown by the arrow, ammonium ions were added to a portion of the culture. Each sample was filtered, washed, suspended in fresh medium without nitrogen source, and incubated for 2min at 36'C before measuring the initial uptake rate of proline
- - 173 14a gdhCR, gln'" 2 20607b mut2, mut4 5
Inactivation of the proline permease upon addition of 192 1 3 b muf2, pgr 5
18409c mut2 5 1 0 ammonium ions to a proline-grown culture of the wild-type 0 19747c mutl 0.8 1
29679a pgr 1 0 0
- -
- -
strain can be seen in Fig. 3.
Proline-grown C1278b wild-type (control) I8 10 12 Double Effect of' the gdhA Mututions
I t is known that the 'ammonia effect' on the general amino- acid permease is strongly reduced in single gdhA mutants, which are affected in the structural gene of the anabolicNADP- linked glutamate dehydrogenase [3,7]. Since it is now clear that two regulatory mechanisms are involved in the 'ammonia effect', the inactivation of the G D H A locus must at least partially suppress each of the two controls which prevent the
mease also allow the development of active L-proline permease and ureidosuccinic-acid permease. One exception is the pgv mutation which suppresses the 'ammonia effect' on the general amino-acid permease only.
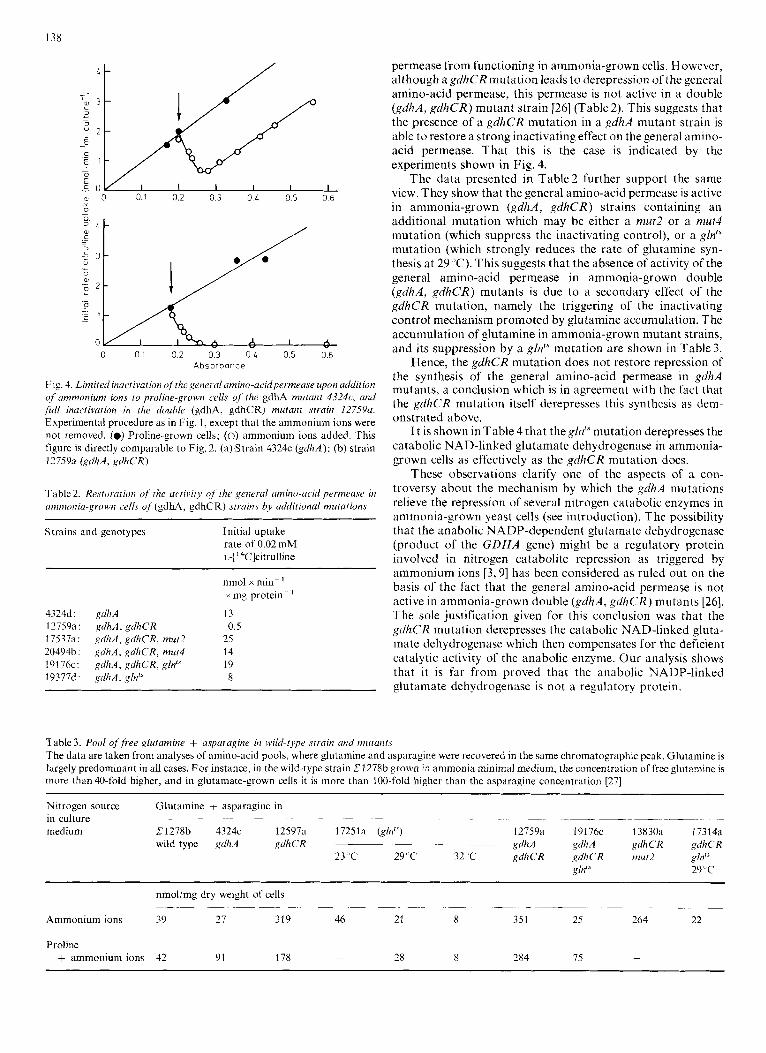
13x
L - permease from functioning in ammonia-grown cells. However, although a gdhCR mutation leads to derepression of the general amino-acid permease, this permease is not active in a double (gdhA, gdhCR) mutant strain [26] (Table 2). This suggests that the presence of a gdhCR mutation in a gdhA mutant strain is able t o restore a strong inactivating effect on the general amino- acid permease. That this is the case is indicated by the experiments shown in Fig. 4.
The data presented in Table2 further support the same a, 0 0.1 0.2 0.3 0 L 0.5 0.6 view. They show that the general amino-acid permease is active
additional mutation which may be either a mur2 or a mut4 mutation (which suppress the inactivating control), or a gln'"
thesis at 29°C). This suggests that the absence of activity of the general amino-acid permease in ammonia-grown double (gdhA, gdhCR) mutants is due to a secondary effect of the gdhCR mutation, namely the triggering of the inactivating control mechanism promoted by glutamine accumulation. The accumulation of glutamine in ammonia-grown mutant strains, and its suppression by a glrt'" mutation are shown in Table 3.
Hence, the gdhCR mutation does not restore repression of
-
I
0 in ammonia-grown (gdhA, gdhCR) strains containing an
mutation (which strongly reduces the rate of glutamine syn-
Y - Q
.-
& 3 -
- 3
L
A
0 0.1 0.2 0.3 O.I 0.5 0.6 Absorbance
Fig. 4. Linzited inactivation ofthe general amino-acidpermease upon addition of ammonium ions to proliiie-grow cells of' ihe gdhA mutatit 4324c, and full inactivation in [he double (gdhA, gdhCR) mutant strain 12759~. Experimental procedure as in Fig. 1, except that the ammonium ions were not removed. (0) Proline-grown cells; (0) ammonium ions added. This figure is directly comparable to Fig. 2. (a) Strain 4324c (gdhA); (b) strain 1275% (gdh.4, gdhCR)
Table 2. Restoration of the acriviiy of the general umbio-acid permease irz ai??nioizia-grol.~n cells of (gdhA, gdhCR) strains by additional mutations
Strains and genotypes Initial uptake rate of 0.02 mM ~-['~C]citrulline
nmol x min- x mg protein- '
4324d: gdhA 13 12759a: gdhA, gdliCR 0.5 17537a: XrfhA, gdhCR, mut2 25 20494b: gdhA, gdhCR, mu14 14
19377d: gdhA, gbi" 8 19176~: gdhA, gdhCR, gln" 19
the synthesis of the general amino-acid permease in gdhA mutants, a conclusion which is in agreement with the fact that the gdhCR mutation itself derepresses this synthesis as dem- onstrated above.
I t is shown in Table 4 that the gln'" mutation derepresses the catabolic NAD-linked glutamate dehydrogenase in ammonia- grown cells as effectively as the gdhCR mutation does.
These observations clarify one of the aspects of a con- troversy about the mechanism by which the gdhA mutations relieve the repression of several nitrogen catabolic enzymes in ammonia-grown yeast cells (see introduction). The possibility that the anabolic NADP-dependent glutamate dehydrogenase (product of the GDHA gene) might be a regulatory protein involved in nitrogen catabolite repression as triggered by ammonium ions [3,9] has been considered as ruled out on the basis of the fact that the general amino-acid permease is not active in ammonia-grown double (gdhA, gdhCR) mutants [26]. The sole justification given for this conclusion was that the gdhCR mutation derepresses the catabolic NAD-linked gluta- mate dehydrogenase which then compensates for the deficient catalytic activity of the anabolic enzyme. Our analysis shows that it is far from proved that the anabolic NADP-linked glutamate dehydrogenase is not a regulatory protein.
Table 3 . Pool of free glutamiiie + aspuragine in tvild-type srrain arid niuiaiits The data are taken from analyses of amino-acid pools, where glutamine and asparagine were recovered in the same chromatographic peak. Glutamine is largely predominant in all cases. For instance, in the wild-type strain Z 1278b grown in ammonia minimal medium, the concentration of free glutamine is more than 40-fold higher, and in glutamate-grown cells it is more than 100-fold higher than the asparagine concentration [27]
Nitrogen source Glutamine + asparagine in in culture medium
- -~ -
C1278b 4124c 12597a 1 7 2 5 1 ~ (gld ' ) wild type gdhA gdhCR _ _ _ _ ~
23 C 29 C 72 c
~ ~
127596 19176~ 13830a 17314a gtlhA d h . 4 gdhCR gdhCR gdliCR gdhCR nzur2 gln"
gln" 29 c nmol/mg dry weight of cells
~ -~ -~ ~ ~ ~-~ ~ - - _ _ Ammonium ions 39 27 319 46 21 8 151 25 264 22
Proline + ammonium ions 42 91 178 28 8 284 75

139
Table 4. Propcr/ies (:f'gln'' cnrd gdhCR rnutunt .vfruins The cells were grown in ammonia minimal medium. Measurements were made in mid-log phase of growth. NAD-linked glutamate dehydrogenase (catabolic) activitv was measured at 30°C
Strains and genotypes
-
Mean Activity of cgeneration glutamate time dehydrogenase
"AD)
h pmol x mg protein-' x min-' 4324d: ,qcIhA 5 2 21278h: wild type 2 3 17251a: Rln" 2.5 146 12597a: gtl/iCR 2 150
DISC USS 1 O N
The data presented here show that upon addition of ammonium ions to a proline-grown culture of a wild-type strain of yeast, the general amino-acid permease is pro- gressively and completely inactivated. Simultaneously, further synthesis of the permease is prevented by repression. That these processes are linked to two distinct regulatory mechanisms is shown by the fact that they are lost separately as a result of mutations. Mutants affected at the MUT2, MUT4, or PGR loci have lost the inactivation mechanism of the general amino-acid permease, whereas gdhCR and gln" mutants have lost the repression of the synthesis of the permease in the presence of ammonium ions. The general amino-acid permease is fully active in ammonia-grown double mutants bearing one mu- tation of each of these two groups.
Also the proline permease and the ureidosuccinic-acid permease are fully active in the same ammonia-grown double mutants. The effect of the pgr mutation, however, is strictly limited to the general amino-acid permease. This is in keeping with the fact that thepgr mutation is located in the GAP1 locus which is the putative structural gene of the general amino-acid permease [20].
The results of the genetic analysis of the pgr mutation [20] were compatible with two possibilities, namely that the mu- tation affects either a promoter-like structure, or a regulatory site which is transcribed and possibly translated. In any case, based on the fact that the semidominant pgr mutation has the same effect on the general amino-acid permease as the recessive mut2 and mut4 mutations, it was concluded that the PGR region determines a receptor site for the negative control mediated by the products of the MUT2 and the MUT4 sites. It seems now clear that these mutations affect the regulatory mechanism which inactivates the general amino-acid permease when ammonium ions are added to the culture medium, and hence that the PGR site must be translated.
The fact that the association of mutations of the same two groups (namely, a mut2 or a mut4 mutation with a gdhCR or a glrP mutation) also completely suppresses the 'ammonia effect' on the proline permease and the ureidosuccinic-acid perinease indicates that these permeases are also regulated both by inactivation and by repression. Inactivation of the proline permease upon addition of ammonium ions has been observed directly.
The addition of ammonium ions to a proline-grown gdhA mutant strain does not seem to provoke repression of the
general amino-acid permease, but the pre-existing activity partially disappears. The persistance of an inactivating process in a gdhA mutant seems to be responsible for the fact that the general amino-acid permease is not active in a double (gc/hA, gdizCR) mutant strain grown in the presence of ammonium ions, as developed under Results.
A n important point about the perinease-inactivating pro- cess is its reversibility, a useful character for a regulatory mechanism. We have observed that the general amino-acid permease is rapidly reactivated when aminonia is removed before repression has developed. That this reactivation seems to depend on the product of the N P R l gene is supported by the results presented in the accompanying paper [21].
1 gratefully acknowledge thc skilful tcchnical assistance of F. Muyldermans-Verhaegen, and 1 thank J . 2 . ten Have who performed the amino-acid analyses. This work was supported by grant 2.4929/79 from the Fonds dr lu Recherche Fonciumentale Collectiw
REFERENCES
1. Schwenckc, J . & Magana-Schwencke, N . (1969) Binchin?. Biophys.
2. Grenson, M., Hou, C . & Crabeel, M. (1970) J . Bacreriol. 103, 770-
3. Grenson, M. & Hou, C. (1972) Binchrrn. Binphys. Res. Commun. 48,
Acts, 173, 302 - 3 12.
777.
4. 5.
6. 7.
8.
9.
10.
1 1 . 12.
13. 14.
15.
16.
17. 18. 19. 20. 21. 22.
23.
24.
2s.
26.
27.
749 - 756. Drillien, R. & Lacroute, F. (1972) J . Bucteriol. 109, 203-208. Darte, C. & Grenson, M. (2975) Biochem. Biophys. Rex Cnmmun. 67,
Duhois, E. & Grenson, M. (1979) Mol. Gen. Genet. 175, 67-76. Grenson, M., Dubois, E., Piotrowska, M., Drillien, R. & Aigle, M.
Dubois, E., Grenson, M. & Wiame, J.-M. (1973) Biochem. Biophys.
Duhois, E., Grenson, M. & Wiame, J.-M. (1974) Eur. J. Biochem. 48,
Duhois, E., Vissers, S., Grenson, M. & Wiame, J.-M. (1977) Biocheni.
Darte, C. (1976) Thesis, University of Brussels. Arst, H. N. & Mac Donald, D. W. (1973) Mol. Grn. Genet. 122,261 -
Kinghorn,J. R. &Pateman,J.A.(1973)J.Gen. Microbiol. 78,39-46. Drillien, R., Aigle, M. & Lacroute, F . (1973) Biochem. Biophys. Res.
Duhois, E. &Grenson, M. (1974) Biochem. Biophys. Res. Commun. 60,
Legrain, C., Vissers, S., Dubois, E., Legrain. M. & Wiame. J.-M. (1982)
Lacroute, F. (1966) Thesis, University of Paris. Grenson, M. (1969) Eur. J . Biochem. 11, 249-260. Grenson, M. & Dubois, E. (1982) Eur. J . Biochem. 121, 643-647. Grenson, M. & Achcroy, 8. (1982) Mol. Gtw. Genet. 188, 261 - 265. Grcnson, M. (1983) Eur. J . Bioc,hm. 133, 142 - 144. Bechet, J., Grenson, M. & Wiame, J . M. (1970) Eur. J . Biochem. 12,
Jacobs, P., Jauniaux, J . C. & Grenson, M. (1980) J . Mol. B id . 139,
Grenson, M., Moussct, M., Wiame, J.-M. & Bechet, J. (1966) Biodirn.
Boehringer, C. F. (1968) Bioclrimicu Curnlo,que, Biochimicd, Mann-
Roon, R. J ., Even, H. L. & Larimore, F. (1974) J . Buctrriol. 118, 89-
Ramos, F. & Wiame, J.-M. (1979) Eur. J . Biochem. 94, 409-417.
1028- 1033.
(1974) Mol. Gar. Genet. 128, 73-85.
Res. Commun. 50, 967- 972.
603 - 616.
Biophys. Res. Commun. 75, 233 - 239.
265.
Cornnrun. 53, 367 - 372.
150-157.
Eur. J . Biochem. i23, 61 1 - 616.
31 -39.
691 - 704.
Biophys. Acta, 127, 325 - 338.
heim.
95.
M. Grenson, Laboratoire de Microbiologie, Faculte des Sciences de I' Universite Libre- de Bruxelles, c/o lnstitut de Recherches du Centre d'Eiiseignement et de Recherches des Industries Alimentaires et Chimiques. Avenue Emile-Gryzon 1, B-1070 Bruxelles, Belgium





























