Embed Size (px)
Citation preview

Isoelectrofocusing Phenotype and Relative Concentrationof Transcobalamin II Isoproteins Related to the Codon259 Arg/Pro Polymorphism
Fares Namour, Muriel Guy, Isabelle Aimone-Gastin, Marie de Nonancourt,Nadir Mrabet, and Jean-Louis Gueant1
Laboratoire de Pathologie Cellulaire et Moleculaire en Nutrition, EP CNRS 616, Faculte de Medecine,B.P. 184, 54505 Vandoeuvre les Nancy Cedex
Received September 8, 1998
We investigated transcobalamin II (TC) isoelectrofo-cusing (IEF) phenotype and codon 259 polymorphism,in Caco-2 and HT-29 cells and in blood drawn from 39healthy Caucasians. Caco-2 cells expressed a single TCvariant (259-Arg), while HT-29 cells expressed TC witheither Arg or Pro at codon 259 and exhibited two iso-proteins in IEF with urea, but only one in IEF withouturea. Among the Caucasians, 7 subjects expressed theTC 259-Arg variant, 10 the 259-Pro variant, and 22were heterozygous. The TC 259-Pro isoprotein issuedfrom HT-29 cells and heterozygous caucasian sera,was, respectively, 2.4-fold and 1.6-fold higher than theTC 259-Arg isoprotein. Apo-TC and vitamin B12 serumconcentrations in 259-Pro homozygotes were, respec-tively, 1.7 and 1.4-fold higher than those in 259-Arghomozygotes (p<0.005 and p50.05). In conclusion, the259-Arg/Pro polymorphism yields two TC variantsonly titratable in denaturing conditions and affectsthe blood level of both Apo-TC and vitamin B12. © 1998
Academic Press
Vitamin B12 (cobalamin, cbl) binds to 3 proteins inmammals: intrinsic factor (IF), transcobalamin II (TC)and haptocorrin (Hc) (1). TC is a 43 kDa non-glycosylated protein that allows vitamin B12 to beinternalized into peripheral cells via receptor mediatedendocytosis (2,3). It also participates in vitamin B12transcytosis from the apical to the basal side of epithe-lial cells as demonstrated in Caco-2 and HT-29 cells, 2human colon adenocarcinoma cell lines with fetal en-terocyte features (4,5,6,7,8,9,10).
Comparison of cDNA clones issued from human um-bilical vein endothelial cells (HUVE), Caco-2 cells andhuman fibroblasts, reveals the existence of 5 single
point substitutions at codons 198, 219, 234, 259 and376 (11,12,13). Several TC isopeptides have been char-acterized in human plasma by polyacrylamide gel elec-trophoresis (PAGE) and isoelectric focusing (IEF) inagarose slab gels. They divide into 4 common pheno-types named X, S, M and F, and 4 rare phenotypesnamed DPAV, VMAR, KDEN and HPHI (14,15). HT-29cells have been shown to secrete a TC isoprotein with ameasured isoelectric point (pI) at 6.4 (8). The pI of TCexpressed in Caco-2 cells was not measured but thepredicted one was found at 6.85 (13).
The phenotypic heterogeneity of TC is attributed to 2mechanisms: a splicing of the signal peptide at alter-native sites (16) and/or a genetic polymorphism whichconsists in the replacement of an uncharged residue(Glu or Pro) by Arg (13). In the previous studies con-cerning TC secreted by established cell lines such asHUVE and Caco-2 cells, the pI values were predictedfrom the cDNA deduced aminoacid sequence (12,13).The present study investigates the relationship be-tween codon 259 polymorphism, IEF phenotyping andrelative concentrations of TC variants expressed inHT-29 and Caco-2 cells and in blood from healthyCaucasians.
MATERIAL AND METHODS
Cell culture and blood collection. Caco-2 and HT-29 cells wereroutinely grown in Dulbecco’s modified Eagle medium with 1% non-essential amino acids (for Caco-2 cells only) and decomplementedfetal calf serum (20% for Caco-2 and 10% for HT-29 cells) (LifeTechnologies, Cergy Pontoise, France), in an atmosphere of 10% CO2and at 37°C. The cells were lysed on day 15 and the supernatant wascollected for further experiments. Blood samples were collected onEDTA from 39 healthy caucasians individuals (age range 25-75years, median 51 years), after informed consent and approval by theethical committee of the university hospital.
Extraction of RNA and genomic DNA from Caco-2, HT-29, andblood cells. Total RNA was prepared with guanidium thiocyanatefollowed by centrifugation in cesium chloride (17) and genomic DNA
1 To whom correspondence should be addressed. Fax: (33)3.83.59.27.98. E-mail: [email protected].
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 251, 769–774 (1998)ARTICLE NO. RC989463
769 0006-291X/98 $25.00Copyright © 1998 by Academic PressAll rights of reproduction in any form reserved.

was purified by the QIAamp spin column procedure (Quiagen,Hilden, Germany).
RT-PCR and amplification of genomic DNA. Ten matched senseand antisense oligonucleotides, localized in separate exons, wereused to amplify 5 overlapping DNA fragments, covering the entirecoding region of TC (1284 nucleotides) (Table 1). Briefly, RNA (4 mg)was reverse transcribed to cDNA after 1 h incubation at 42°C, in a 20mL reaction volume containing 10 mM DTT, 500 mM dNTP, 7 pmolspecific antisens primer and 200 U M-MLV reverse transcriptase(Life Technologies, Cergy Pontoise, France). An aliquot (10%) of thereaction mixture was subjected to amplification on the GeneAmpPCR System 2400 (Perkin Elmer, Norwalk CT, USA) in a reactionvolume of 50 mL containing 1.5 mM MgCl2, 200 mM dNTP, 15 pmolof each primer and 1.25 U of Taq polymerase (Perkin Elmer, RocheMolecular Systems, Branchburg NJ, USA). After an initial denatur-ation step at 94°C for 2 minutes, amplification was carried out for 30cycles, each cycle consisting of a 30-s denaturation time at 94°C, a30-s annealing time at a variable temperature and a 50-s extensiontime at 72°C, followed by a final step at 72°C for 10 minutes. The twogenomic DNA fragments corresponding to the TC regions containing
codons 259 and 341 were amplified with specific primers under thesama conditions (Table 1). The PCR products were visualized on an1.5% agarose gel.
DNA sequencing. DNA was directly subjected to double strandsequence analysis with AmpliTaq FS by the Dye deoxy terminatorcycle sequencing method (PE Applied Biosystems, Warrington,Great Britain). The reaction volume was 20 mL containing 12 ng ofDNA and 3.2 pmol of specific primer. Thermal cycling was carriedout for 25 cycles, each consisting of a step at 96°C for 10 s, a step at50°C for 5 s and a step at 60°C for 4 minutes. The amplificationproduct was then ethanol precipitated and loaded for an overnightmigration on a 6% polyacrylamide gel in the ABI 373 DNA sequencer(PE Applied Biosystems, Norwalk CT, USA). For each single frag-ment, the DNA sequenced was issued from at least 3 independentamplification reactions.
Gel filtration. A supernatant aliquot concentrated 10-fold (100mL for Caco-2 and 300 mL for HT-29 cells) or 300 mL of plasma, wasincubated with 20 mL (0.74 pmol) of cyano(57Co)cbl (Amersham, LesUlis, France) for 20 minutes under shaking at room temperature.
TABLE 1
Oligonucleotide Primer Sequences Used in RT-PCR and Genomic DNA Amplification
Matched primers used in RT-PCR
Fragment length(pb)
59 primers and localizationin the gene
39 primers and localizationin the gene
Annealingtemperature
400 S1: 237 AS1: 343 60°CCCGATTCTTGCT GCCCCTGACCACTGCTCA 217 AAACTCACAGTT 36359 non-coding region Exon 3
404 S2: 286 AS2: 670 63°CTGCCAGGGCA GATCTCCTCTAGCCTTCCAT 305 cgcactgttct 690Exon 3 Exon 5
356 S3: 607 AS3: 943 64°CTTCACCTGTCT CTCAGCAGCTGAAGCGCTCA 627 GGTTCCAACAT 963Exon 5 Exon 7
347 S4: 883 AS4: 1210 62°CGTTCTGAACCA AGCAATACCCAAGACCTAC 903 TTGCAACAGTGG 1230Exon 6 Exons 8/9
179 S5: 1141 AS5: 914 60°CTTAACCTCCG CCTCGCACATGATGGGGAAA 1161 CTCCCTAGGCTT 1320Exon 8 39 non-coding region
Matched primers used in genomic DNA amplification
Codon number includedin the fragment
59 primers and localizationin the gene
39 primers and localizationin the gene
Annealingtemperature
259 S6: 700 AS6: 914 60°CAGAACAGTGC CTTCCCAGACGAGAGGAGATC 721 TGTCTGGCAC 934Exon 5 Exon 6
341 S7: 972 AS7: 1086 56°CATGTTGGAAC CCATGAGTTAGCTGCTGA 991 AGGAGGATTCA 1105Exon 7 Exon 7
Vol. 251, No. 3, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
770

The sample was then applied to a Sephacryl S300 HR column (Phar-macia, Uppsala, Sweden) and eluted with 20 mM Tris HCl buffer pH7.4 containing 0.7M NaCl, 0.01% Triton, 0.01% NaN3 at a flow rateof 0.3 mL/min. The fractions containing saturated TC (0.3 mL each)were collected for IEF. In one set of experiments, bovine serumalbumin was added to a final concentration of 0.02% (w/v) to thetubes in which TC was collected during gel filtration.
Isoelectrofocusing. IEF was performed in a 110 mL LKB column(Pharmacia, Uppsala Sweden) displaying a 5-50% sucrose gradientwith or without 5M urea. The samples were dialyzed overnight at4°C against 400 volumes distilled water with a 10 kDa cut offmembrane. Ampholines with pH 3-10 or 5-7 (Sigma, St Louis MO,USA) were added to a final dilution of 1:33. Focusing was run at 0.5kV for 48-h and refrigeration was provided by a continuous waterflow at room temperature. The pH values were measured with aSchott 2998 electrode on a Schott CG 825 pH meter (Schott-Gerate,Hofheim, Germany). The calculated pIs of TC were generated by thecomputer programme ExPASy (University of Geneva, Switzerland).
Radioisotope assays of serum vitamin B12 and TC. Vitamin B12was quantified by radioisotope dilution assay (18). Determination ofTC-(57Co)Cbl was performed as previously described (19,20), using asilica matrice (QUSO) to adsorb TC. Briefly, 3.75 g of QUSO wassuspended in 25 mL distilled water and 100 mL of the stirred sus-pension was added to 500 mL of plasma, incubated with (57Co)Cbland centrifuged at 2000 g for 15 minutes at 10°C. The tracer boundto HC was measured by g-counting in the supernatant. The tracerbound to TC was calculated from the difference between wholeplasma and supernatant values. The Mann-Whitney test was used tocompare serum Apo-TC and vitamin B12 concentrations in functionof TC-polymorphism.
RESULTS AND DISCUSSION
We have determined the phenotype of TC secreted bypost-confluent HT-29 and Caco-2 cells grown for 15days. The 10-fold concentrated culture medium waslabelled with (57Co)-cbl and submitted to gel filtrationon a pre-calibrated Sephacryl S-300 HR column, inorder to separate the TC-(57Co)cbl complex from Hc (8).In some experiments, albumin was added to to thetubes in which TC was collected during gel filtration(0.02%), for the purpose of counteracting aggregation(15). Liquid column IEF was chosen because it enablesto detect narrow pH ranges more efficiently than IEFin slab gels.
When IEF was performed under non-denaturing con-ditions (without urea), the TC-(57Co)cbl complex fromHT-29 cells was found to be more acidic than that fromCaco-2 cells, since the respective pIs were at 6.4 and6.6 (Table 2 and Fig.1). This result was confirmed afteranother IEF, performed with a mixture of TCs fromboth cell lines, showed two distinct isoproteins. Previ-ous data have already suggested that TC expressed inHT-29 and Caco-2 cells belong to different phenotypes(8,13).
The TC microheterogeneity is not related to isopro-tein size (14), but can be explained either by a geneticpolymorphism responsible for the protein charge mod-ification (13), or a differential signal peptide cleavagesite on the nascent protein (16). Indeed, Li et al. pro-posed that residues Arg and Arg, Gln and Arg, and Glnand Pro present at positions 234 and 259, respectively,characterize the X, S and M phenotypes of TC in Caco-2cells and human fibroblasts (13). Quadros et al., afterproducing recombinant TC by using the baculovirusexpression system, emphasized the existence of 2 sig-nal peptidase recognition sequences on the leader pep-tide: one is the triplet alanine-leucine-threonine whichyields an isoprotein with glutamic acid and methionineas the first and second aminoacid and the other isthreonine-glutamic acid-methionine which yields, in-stead, an isoprotein with cysteine and methionine asthe first and second amino acid (16).
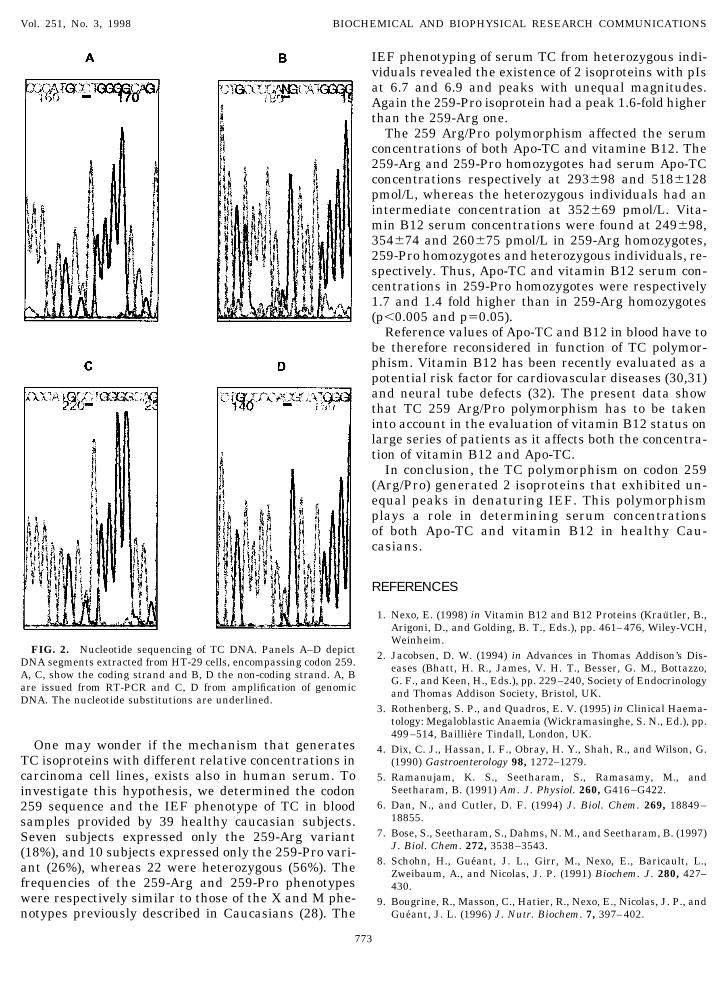
In order to recognize a putative genetic polymor-phism underlying the phenotypic microheterogeneityof TC, we amplified the cDNA from HT-29 and Caco-2cells by RT-PCR and determined the complete nucleo-tide sequence. Five overlapping fragments, encompass-ing the entire coding region, were amplified at least 3times by means of independent reactions, and yieldedthe expected sizes (400, 404, 356, 347 and 179 bp), asevidenced by electrophoresis in agarose gels. Eachfragment was double strand sequenced at least 3times. TCs expressed by HT-29 and Caco-2 cells hadthe same cDNA sequence except for codons 259 and341. We found that at codon 259, HT-29 cells are het-
TABLE 2
Genotypic and Phenotypic Polymorphisms of TC Secreted by Caco-2 and HT-29 Adenocarcinoma Cell Lines
Cell lines
Caco-2 HT-29
Codons 234 CAG (Gln) CAG (Gln)259 CGT (Arg) CGT (Arg) or CCT (Pro)341 CCA (Pro) CCG (Pro)
Measured pIs IEF without 5M urea 6.6 6.4IEF with 5M urea 6.7 6.6 and 6.8
Calculated pIs signal peptidase cleavage site: Ala-Leu-Thr 6.8 6.7 and 6.8signal peptidase cleavage site: Thr-Glu-Met 6.9 6.8 and 6.9
Vol. 251, No. 3, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
771
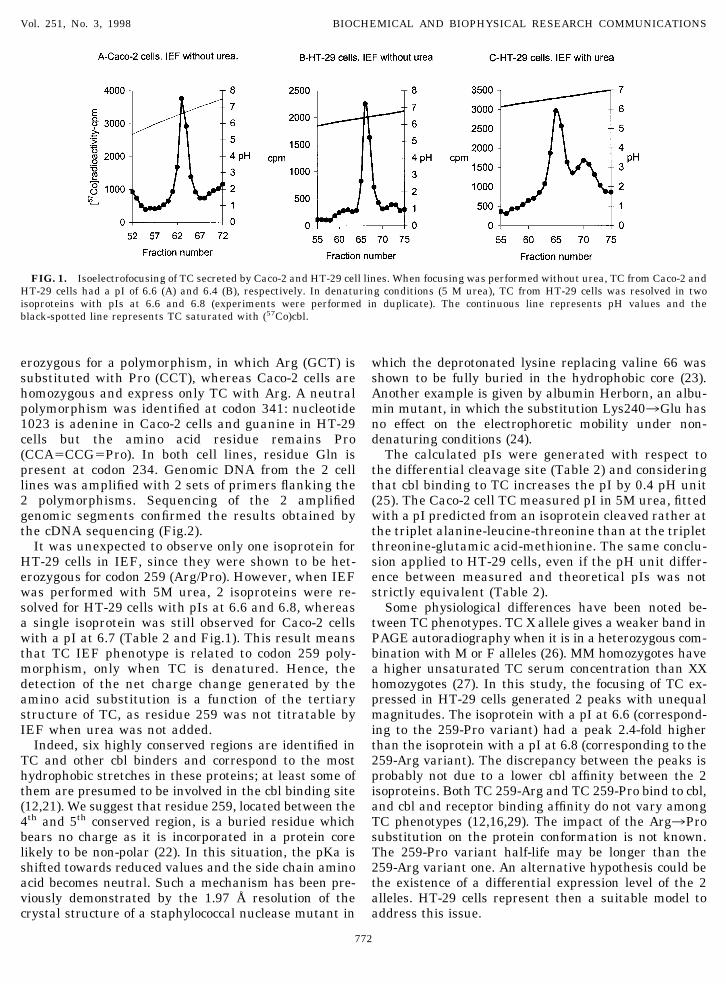
erozygous for a polymorphism, in which Arg (GCT) issubstituted with Pro (CCT), whereas Caco-2 cells arehomozygous and express only TC with Arg. A neutralpolymorphism was identified at codon 341: nucleotide1023 is adenine in Caco-2 cells and guanine in HT-29cells but the amino acid residue remains Pro(CCA5CCG5Pro). In both cell lines, residue Gln ispresent at codon 234. Genomic DNA from the 2 celllines was amplified with 2 sets of primers flanking the2 polymorphisms. Sequencing of the 2 amplifiedgenomic segments confirmed the results obtained bythe cDNA sequencing (Fig.2).
It was unexpected to observe only one isoprotein forHT-29 cells in IEF, since they were shown to be het-erozygous for codon 259 (Arg/Pro). However, when IEFwas performed with 5M urea, 2 isoproteins were re-solved for HT-29 cells with pIs at 6.6 and 6.8, whereasa single isoprotein was still observed for Caco-2 cellswith a pI at 6.7 (Table 2 and Fig.1). This result meansthat TC IEF phenotype is related to codon 259 poly-morphism, only when TC is denatured. Hence, thedetection of the net charge change generated by theamino acid substitution is a function of the tertiarystructure of TC, as residue 259 was not titratable byIEF when urea was not added.
Indeed, six highly conserved regions are identified inTC and other cbl binders and correspond to the mosthydrophobic stretches in these proteins; at least some ofthem are presumed to be involved in the cbl binding site(12,21). We suggest that residue 259, located between the4th and 5th conserved region, is a buried residue whichbears no charge as it is incorporated in a protein corelikely to be non-polar (22). In this situation, the pKa isshifted towards reduced values and the side chain aminoacid becomes neutral. Such a mechanism has been pre-viously demonstrated by the 1.97 Å resolution of thecrystal structure of a staphylococcal nuclease mutant in
which the deprotonated lysine replacing valine 66 wasshown to be fully buried in the hydrophobic core (23).Another example is given by albumin Herborn, an albu-min mutant, in which the substitution Lys2403Glu hasno effect on the electrophoretic mobility under non-denaturing conditions (24).
The calculated pIs were generated with respect tothe differential cleavage site (Table 2) and consideringthat cbl binding to TC increases the pI by 0.4 pH unit(25). The Caco-2 cell TC measured pI in 5M urea, fittedwith a pI predicted from an isoprotein cleaved rather atthe triplet alanine-leucine-threonine than at the tripletthreonine-glutamic acid-methionine. The same conclu-sion applied to HT-29 cells, even if the pH unit differ-ence between measured and theoretical pIs was notstrictly equivalent (Table 2).
Some physiological differences have been noted be-tween TC phenotypes. TC X allele gives a weaker band inPAGE autoradiography when it is in a heterozygous com-bination with M or F alleles (26). MM homozygotes havea higher unsaturated TC serum concentration than XXhomozygotes (27). In this study, the focusing of TC ex-pressed in HT-29 cells generated 2 peaks with unequalmagnitudes. The isoprotein with a pI at 6.6 (correspond-ing to the 259-Pro variant) had a peak 2.4-fold higherthan the isoprotein with a pI at 6.8 (corresponding to the259-Arg variant). The discrepancy between the peaks isprobably not due to a lower cbl affinity between the 2isoproteins. Both TC 259-Arg and TC 259-Pro bind to cbl,and cbl and receptor binding affinity do not vary amongTC phenotypes (12,16,29). The impact of the Arg3Prosubstitution on the protein conformation is not known.The 259-Pro variant half-life may be longer than the259-Arg variant one. An alternative hypothesis could bethe existence of a differential expression level of the 2alleles. HT-29 cells represent then a suitable model toaddress this issue.
FIG. 1. Isoelectrofocusing of TC secreted by Caco-2 and HT-29 cell lines. When focusing was performed without urea, TC from Caco-2 andHT-29 cells had a pI of 6.6 (A) and 6.4 (B), respectively. In denaturing conditions (5 M urea), TC from HT-29 cells was resolved in twoisoproteins with pIs at 6.6 and 6.8 (experiments were performed in duplicate). The continuous line represents pH values and theblack-spotted line represents TC saturated with (57Co)cbl.
Vol. 251, No. 3, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
772
One may wonder if the mechanism that generatesTC isoproteins with different relative concentrations incarcinoma cell lines, exists also in human serum. Toinvestigate this hypothesis, we determined the codon259 sequence and the IEF phenotype of TC in bloodsamples provided by 39 healthy caucasian subjects.Seven subjects expressed only the 259-Arg variant(18%), and 10 subjects expressed only the 259-Pro vari-ant (26%), whereas 22 were heterozygous (56%). Thefrequencies of the 259-Arg and 259-Pro phenotypeswere respectively similar to those of the X and M phe-notypes previously described in Caucasians (28). The
IEF phenotyping of serum TC from heterozygous indi-viduals revealed the existence of 2 isoproteins with pIsat 6.7 and 6.9 and peaks with unequal magnitudes.Again the 259-Pro isoprotein had a peak 1.6-fold higherthan the 259-Arg one.
The 259 Arg/Pro polymorphism affected the serumconcentrations of both Apo-TC and vitamine B12. The259-Arg and 259-Pro homozygotes had serum Apo-TCconcentrations respectively at 293698 and 5186128pmol/L, whereas the heterozygous individuals had anintermediate concentration at 352669 pmol/L. Vita-min B12 serum concentrations were found at 249698,354674 and 260675 pmol/L in 259-Arg homozygotes,259-Pro homozygotes and heterozygous individuals, re-spectively. Thus, Apo-TC and vitamin B12 serum con-centrations in 259-Pro homozygotes were respectively1.7 and 1.4 fold higher than in 259-Arg homozygotes(p,0.005 and p50.05).
Reference values of Apo-TC and B12 in blood have tobe therefore reconsidered in function of TC polymor-phism. Vitamin B12 has been recently evaluated as apotential risk factor for cardiovascular diseases (30,31)and neural tube defects (32). The present data showthat TC 259 Arg/Pro polymorphism has to be takeninto account in the evaluation of vitamin B12 status onlarge series of patients as it affects both the concentra-tion of vitamin B12 and Apo-TC.
In conclusion, the TC polymorphism on codon 259(Arg/Pro) generated 2 isoproteins that exhibited un-equal peaks in denaturing IEF. This polymorphismplays a role in determining serum concentrationsof both Apo-TC and vitamin B12 in healthy Cau-casians.
REFERENCES
1. Nexo, E. (1998) in Vitamin B12 and B12 Proteins (Krautler, B.,Arigoni, D., and Golding, B. T., Eds.), pp. 461–476, Wiley-VCH,Weinheim.
2. Jacobsen, D. W. (1994) in Advances in Thomas Addison’s Dis-eases (Bhatt, H. R., James, V. H. T., Besser, G. M., Bottazzo,G. F., and Keen, H., Eds.), pp. 229–240, Society of Endocrinologyand Thomas Addison Society, Bristol, UK.
3. Rothenberg, S. P., and Quadros, E. V. (1995) in Clinical Haema-tology: Megaloblastic Anaemia (Wickramasinghe, S. N., Ed.), pp.499–514, Bailliere Tindall, London, UK.
4. Dix, C. J., Hassan, I. F., Obray, H. Y., Shah, R., and Wilson, G.(1990) Gastroenterology 98, 1272–1279.
5. Ramanujam, K. S., Seetharam, S., Ramasamy, M., andSeetharam, B. (1991) Am. J. Physiol. 260, G416–G422.
6. Dan, N., and Cutler, D. F. (1994) J. Biol. Chem. 269, 18849–18855.
7. Bose, S., Seetharam, S., Dahms, N. M., and Seetharam, B. (1997)J. Biol. Chem. 272, 3538–3543.
8. Schohn, H., Gueant, J. L., Girr, M., Nexo, E., Baricault, L.,Zweibaum, A., and Nicolas, J. P. (1991) Biochem. J. 280, 427–430.
9. Bougrine, R., Masson, C., Hatier, R., Nexo, E., Nicolas, J. P., andGueant, J. L. (1996) J. Nutr. Biochem. 7, 397–402.
FIG. 2. Nucleotide sequencing of TC DNA. Panels A–D depictDNA segments extracted from HT-29 cells, encompassing codon 259.A, C, show the coding strand and B, D the non-coding strand. A, Bare issued from RT-PCR and C, D from amplification of genomicDNA. The nucleotide substitutions are underlined.
Vol. 251, No. 3, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
773

10. Gueant, J. L., Schohn, H., Bougrine, R., Masson, C., Gastin, I.,Nicolas, J. P., Leheup, B., Grignon, G., Chantret, I., Rousset, M.,and Nexo, E. (1994) in Advances in Thomas Addison’s Diseases(Bhatt, H. R., James, V. H. T., Besser, G. M., Bottazzo, G. F., andKeen, H., Eds.), pp. 229–240, Society of Endocrinology andThomas Addison Society, Bristol, UK.
11. Platica, O., Janeckzo, R., Quadros, E. V., Regec, A., Romain, R.,and Rothenberg, S. P. (1991) J. Biol. Chem. 266, 7860–7863.
12. Li, N., Seetharam, S., Lindemans, J., Alpers, D. H., Arwert,F., and Seetharam, B. (1993) Biochem. Biophys. Acta 1172,21–30.
13. Li, N., Sood, G. K., Seetharam, S., and Seetharam, B. (1994)Biochem. Biophys. Acta 1219, 515–520.
14. Frater-Schroder, M. (1983) Mol. Cell. Biochem. 56, 5–31.15. Hansen, M., and Frater-Schroder, M. (1987) Electrophoresis 8,
221–223.16. Quadros, E. V., Sai, P., and Rothenberg, S. P. (1993) Blood 81,
1239–1245.17. Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) in Molecu-
lar Cloning, A Laboratory Manual, 2nd Edition., pp. 7.19–7.22,Cold Spring Harbor, New York.
18. Djalali, M., Gueant, J. L., Lambert, D., El Kholty, S., Saunier,M., and Nicolas, J. P. (1990) J. Chromat. 529, 81–91.
19. Benhayoun, S., Adjalla, C., Nicolas, J. P., Gueant, J. L., andLambert, D. (1993) Acta Haematol. 89, 195–199.
20. Jacob, E., Wrong, K. J. T., and Herbert, V. (1977) J. Lab. Med.89, 1145–1152.
21. Gueant, J. L., Safi, A., Gastin, I., Rabesona, H., Bronowicki, J. P.,
Plenat, F., Bigard, M. A., and Haertle, T. (1997) Proceedings Am.Ass. Phys. 109, 462–469.
22. Stigter, D., and Dill, K. A. (1990) Biochemistry 29, 1262–1271.23. Stites, W. E., Gittis, A. G., Lattman, E. E., and Shortle, D. (1991)
J. Mol. Biol. 221, 7–14.24. Minchiotti, L., Galliano, M., Zapponi, M. C., and Tenni, R. (1993)
Eur. J. Biochem. 214, 437–444.25. Hansen, M., and Nexo, E. (1989) Biochem. Biophys. Acta 992,
209–214.26. Begley, J. A., and Hall, C. A. (1990) in Biomedicine and Physi-
ology of Vitamin B12 (Linnell, J. C., and Bhatt, H. R., Eds.), pp.307–312, The Children’s Medical Charity, London.
27. Porck, H., Frants, R., Lindemans, J., Hooghwinkel, G., andPlanta, R. (1986) Biochem. Genet. 24, 103–114.
28. Arwert, F., Pronk, J. C., Zelle, B., Frater-Schroder, M., Porck,H. J., and Eriksson, A. W. (1990) in Biomedicine and Physiologyof Vitamin B12 (Linnell, J. C., and Bhatt, H. R., Eds.), pp.277–280, The Children’s Medical Charity, London.
29. Hansen, M., and Nexo, E. (1987) Biochim. Biophys. Acta 926,359–354.
30. Pancharuniti, N., Lewis, C. A., Sauberlich, H. E., Perkins, L. L.,Go, R. C., Alvarez, J. O., Macaluso, M., Acton, R. T., Copeland,R. B., and Cousins, A. L. (1994) Am. J. Clin. Nutr. 59, 940–948.
31. Guttormsen, A. B., Ueland, P. M., Nesthus, I., Nygard, O.,Schneede, J., Vollset, S. E., and Refsum, H. (1996) J. Clin. Invest.98, 2174–2183.
32. Eskes, T. K. (1998) Eur. J. Pediatr. 157(Suppl 2), S139–S141.
Vol. 251, No. 3, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
774





























