Embed Size (px)
Citation preview

Eur. J. Biochem. f78, 367 - 372 (1 988) T FEBS 1988
Kinetic and biochemical properties of CTP : choline-phosphate cytidylyltransferase from the rat brain Francoise MACES. Catherine REY, Pierre FONLUPT and Henri PACHECO Unite 205, de 1’Institut National de la SantC et de la Recherche Medicale, Laboratoire de Chimie Biologique, Institut National des Sciences Appliquees, Villeurbanne
(Received May 2O/July 27,1988) - EJB 88 0575
In order to investigate the mechanisms involved in some brain disorders at the membrane level, we studied the kinetics and biochemical properties of brain CTP : choline-phosphate cytidylyltransferase (EC 2.7.7.15), the rate-limiting enzyme of the two-step biosynthesis of phosphatidylcholine. This enzyme catalyzes the biosynthesis of CDPcholine from choline phosphate and CTP. We found that its subcellular localization (mainly in microsomal and cytosolic fractions) was different from that of phosphatidylethanolamine N-methyltransferase (EC 2.1.1.17), the enzyme of the alternative pathway for phosphatidylcholine synthesis.
CTP:choline-phosphate cytidylyltransferase showed a K,,, of 10 mM for CTP and 0.3 mM for choline phos- phate and exhibited a random mechanism. CDPcholine, the reaction product, was a competitive inhibitor of choline phosphate and CTP utilization and had a Ki of 0.090 mM. Both particulate and soluble enzymes required Mg2+ and exhibited an optimal pH at about 7.
Cytosolic activity was enhanced by addition of unsaturated fatty acids or phospholipids extracted from brain membranes. Such an enhancement was increased with the centrifugation time used for preparing the soluble enzyme.
Phosphatidylcholine, a major phospholipid of mammalian cells, is mainly produced from choline phosphate through Kennedy’s pathway [l]. This lipid has primarily a structural function in biologicdl membranes. It may also provide fatty acids, like arachidonic acid for prostaglandin, leukotriene and thromboxane synthesis [2]. Phospholipid synthesis in the brain can modify the affinity of many receptors for their ligands, both endogenous, like catecholamines [3], and exogenous, like benzodiazepines [4].
CTP : choline-phosphate cytidylyltransferase catalyzes the synthesis of CDPcholine, the rate-limiting step of phosphati- dylcholine synthesis in various tissues [5]. The enzyme from several sources exhibits similar properties [6]. In both lung and liver [7], this enzyme is present under two forms.
a) A low-molecular-mass form (L form), which is cytosolic and which presents very low activity. The cytosolic L form exhibits a greater affinity for membranes after in vitro addition of unsaturated fatty acids [8] or in vivo after cholesterol- or cholate-rich diet [9].
b) A high-molecular-mass form (H form), located in cytosol and membranes. The H form, which is the active one, is a mixed aggregate of L form and lipids [lo]. The addition of phosphatidylglycerol or some other lipids [ l l , 121 to the cytosol induces the conversion of the L form into the H one [Ill. ~~
Correspondence 20 p. Fonlupt, INSERM U205, Laboratoire de Chimie Biologique, Batiment 406, Institut National des Sciences Appliquies, 20 Avenue Albert-Einstein, F-69621, Villeurbanne Cedex, France
Enzymes. CTP:choline-phosphatc cytidylyltransferase (EC 2.7.7.15); phosphatidylethanolamine N-methyltransferase (EC 2.1.1.17).
Choline-phosphate cytidylyltransferase from lung has been extensively studied [lo - 131, but few reports concerning the brain enzyme are available yet [14- 161. Nevertheless, studying the brain enzyme is of double interest: (a) choline- phosphate cytidylyltransferase may be modified by lipid en- vironment changes, which occur in some cerebral diseases such as ischemia [17], where the membrane balance is altered and the lipid breakdown increases; (b) CDPcholine, the prod- uct of the enzyme reaction, has been shown to exert protective effects in brain ischemia [18, 191, hypoxia treatment [20] and ageing [21].
In the context of these pathophysiological data, we first studied the kinetic parameters and biochemical properties of the rat brain choline-phosphate cytidylyltransferase from normal rats.
MATERIALS AND METHODS
Muteriuls
Fatty acids, choline phosphate and CTP used in this study were purchased from Sigma (St Louis, MO). [methyl- 14C]Choline phosphate was supplied by NEN (Boston, MA; reference NEC 544), [rvtethyl-’4C]CDPcholine was from Amersham International (Amersham, UK; reference CFA 528) and S-adenosyl-~-[methyl-’H]methionine was from CEA (Saclay, France; reference TMM-141).
Animals, preparation of crude exlract and its subcellular fractionation
Male OFA rats from Iffa Credo (Lyon, France; 180- 200 g) were decapitated. Brains were quickly removed and homogenized in 0.32 M sucrose solution (1 g tissue/l2.5 ml).
368
ffl ffl .- c
m 5 - 1
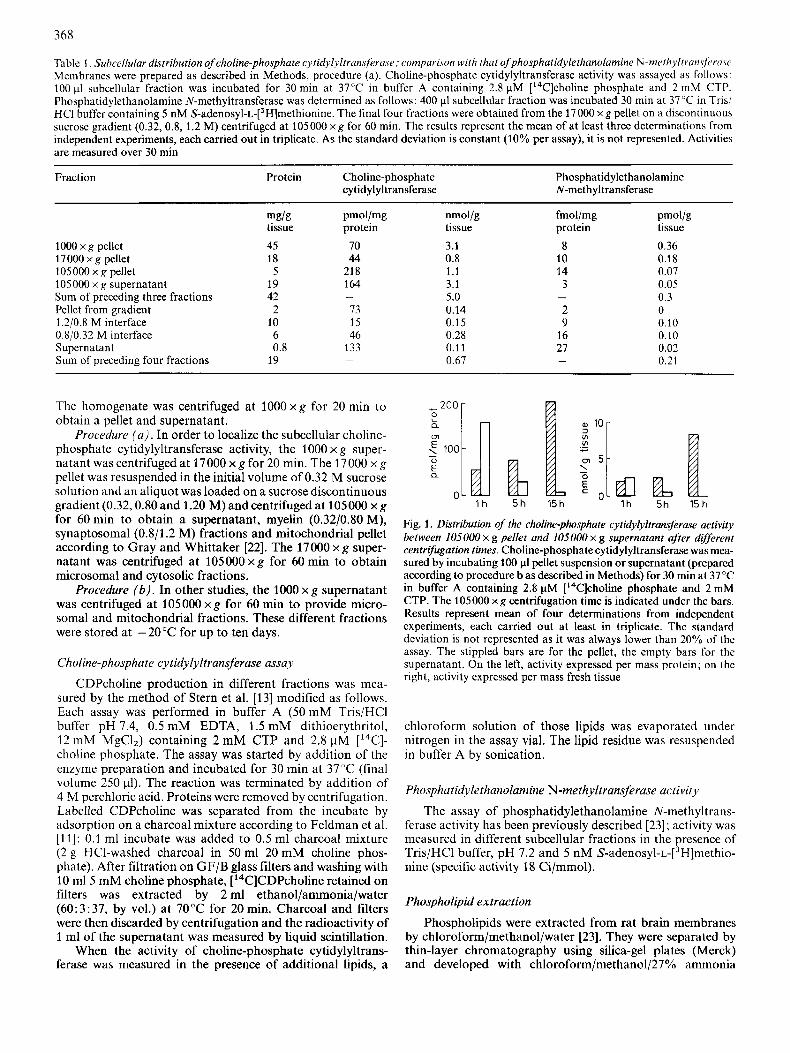
Table 1 . Subcellular distribution of choline-phosphate cytidylyltrunsferuse; comparison with that ofphosphutidylethanolamine N-meth~ltruws~rcrsc Membranes were prepared as described in Methods, procedure (a). Choline-phosphate cytidylyltransferase activity was assayed as follows: 100 pl subcellular fraction was incubated for 30 rnin at 37°C in buffer A containing 2.8 pM ['4C]choline phosphate and 2 mM CTP. Phosphatidylethanolamine N-methyltransferase was dctcrmined as follows: 400 p1 subcellular fraction was incubated 30 rnin at 37 'T in Tris/ HCI buffer containing 5 nM S-adenosyl-~-[~H]methionine. The final four fractions were obtained from the 17000 x g pellet on a discontinuous sucrose gradient (0.32,O.S. 1.2 M) centrifugcd at I05000 x g for 60 min. The results represent the mean of at least three determinations from independent experiments, each carried out in triplicate. As the standard deviation is constant (10% per assay), it is not represented. Activities are measured over 30 min
Fraction Protein Choline-phosphate cytidylyltransferase
Phosphatidylethanolaminc N-meth yltransferase
1000 x g pellet 17OOOxg pellet 105000 x g pellet 1OSOOO x g supernatant Sum of preceding three fractions Pellet from gradient 1.2/0.8 M interface 0.8/0.32 M interface Supernatant Sum of preceding four fractions
mg/g tissue 45 18
5 19 42 2
10 6 0.8
19
pmoI/mg protein
70 44
21 8 164
73 1s 46
133
-
-
nmol/g tissue 3.1 0.8 1.1 3.1 5.0 0.14 0.15 0.28 0.11 0.67
fmol/mg protein
8 10 14 3
2 9
16 27
-
-
pmol/g tissue 0.36 0.18 0.07 0.05 0.3 0 0.10 0.10 0.02 0.21
The homogenate was centrifuged at 1000 x g for 20 min to obtain a pellet and supernatant.
Procedure ( u ) . In order to localize the subcellular choline- phosphate cytidylyltransferase activity, the 1000 x g super- natant was centrifuged at 17000 x g for 20 min. The 17000 x g pellet was resuspended in the initial volume of 0.32 M sucrose solution and an aliquot was loaded on a sucrose discontinuous gradient (0.32,O.SO and 1.20 M) and centrifuged at 105000 x g for 60 rnin to obtain a supernatant, myelin (0.32/0.80 M), synaptosomal (0.8/1.2 M) fractions and mitochondrial pellet according to Gray and Whittaker [22]. The 17000 x g super- natant was centrifuged at 105000 x g for 60 rnin to obtain microsomal and cytosolic fractions.
Procedure (6 ) . In other studies, the 1000 x g supernatant was centrifuged at 105000 x g for 60 rnin to provide micro- soma1 and mitochondrial fractions. These different fractions were stored at -20°C for up to ten days.
Choline-phosphate cytidylyltrun~-jeruse assay
CDPcholine production in different fractions was mea- sured by the method of Stern et al. [13] modified as follows. Each assay was performed in buffer A (50 mM Tris/HCl buffer pH 7.4, 0.5 mM EDTA, 1.5 mM dithioerythritol, 12 mM MgClJ containing 2 mM CTP and 2.8 pM ['4C]- choline phosphate. The assay was started by addition of the enzyme preparation and incubated for 30 rnin at 37°C (final volume 250 pl). The reaction was terminated by addition of 4 M perchloric acid. Proteins were removed by centrifugation. Labelled CDPcholine was separated from the incubate by adsorption on a charcoal mixture according to Feldman et al. [ll]: 0.1 ml incubate was added to 0.5 ml charcoal mixture (2 g HC1-washed charcoal in 50 ml 20mM choline phos- phate). After filtration on GF/B glass filters and washing with 10 m15 mM choline phosphate, ['4C]CDPcholine retained on filters was extracted by 2 ml ethanol/ammonia/water (60:3:37, by vol.) at 70°C for 20 min. Charcoal and filters were then discarded by centrifugation and the radioactivity of 1 ml of the supernatant was measured by liquid scintillation.
When the activity of choline-phosphate cytidylyltrans- ferase was mcasurcd in the presence of additional lipids, a
15h EL 5h
L 15 h
Fig. 1. Dzitributiun of the choline-phosphate cytidylyltransjrrase activity between 105000 x g pellet and 105000 x g supernatant after dgferent centrifugation times. Choline-phosphate cytidylyltransferase was mea- sured by incubating 100 p1 pellet suspension or supernatant (prepared according to procedure bas described in Methods) for 30 min at 37°C in buffer A containing 2.8 pM ['4C]choline phosphate and 2 mM CTP. The 105000 x g centrifugation time is indicated under the bars. Results represent mean of four determinations from indepcndent experiments, each carried out at least in triplicate. The standard deviation is not represented as it was always lower than 20% of thc assay. The stippled bars are for thc pellet, the empty bars for the supernatant. On the left, activity expresscd pcr mass protein; on the right, activity exprcssed per mass fresh tissue
chloroform solution of those lipids was evaporated under nitrogen in the assay vial. The lipid residue was resuspended in buffer A by sonication.
Phosphutidylethanolumine N-methyltransferuse uctivit-v
The assay of phosphatidylethanolamine N-methyltrans- ferase activity has been previously described [23]; activity was measured in different subcellular fractions in the presence of Tris/HCl buffer, pH 7.2 and 5 nM S-adenosyl-~-[~H]methio- nine (specific activity 18 Cilmmol).
Phospholipid extraction
Phospholipids were extracted from rat brain membranes by chloroform/methanol/water [23]. They were separated by thin-layer chromatography using silica-gel plates (Merck) and developed with chloroform/methano1/27% ammonia

369
B I 200 L A l oo i 0 61
2ool 100
0
- 0
C 0 0
L c
C
daB 1 5 15
500 -
V ._ -
centrifugation duration (hours)
Fig. 2. Elf& ofjutiy acids on choline-phosphate cytidylyltransferuse activity in 105000 x g supernatant. Homogenates were centrifuged for 5 h at 10500Oxg (A, B) or for 1, 5 and 15 h (C) (as described in Methods, procedure b). Supernatant choline-phosphate cytidylyltransferase activity determination was pcrformed as described in Fig. 1. Chloroform solutions of fatty acids were added to the assay vial; chloroform was evaporated under nitrogen and the lipid residue was resuspended in buffer A by sonication. Results represent the mean of three determinations from independent experiments, each carried out in triplicate. The standard deviation is not represented and increases from 10% (for the highest values) to 25% of the assay (for the lowest values). The results are expressed as a percentage of control (no Fatty acids added). (A) Effect of various saturated fatty acids at 100 pM (empty blocks) or 200 pM (stippled blocks) as a percentage of control (full block); the homogenate was centrifuged at 105000 x g for 5 h. (B) Effect of unsaturated fatty acids at (El) 4 pM, (El) 20 pM, 40 pM, (empty blocks) 80 pM or (stippled blocks) 200 pM as a percentage of control (full block); the homogenate was centrifuged at 105000 x g for 5 h. (C) Effect of oleic acid at 80 pM (empty blocks) or 200 pM (stippled blocks) on supernatants obtained from different centrifugation times; results are cxpresscd as percentages of the 105000 x g I-h supematant activity (full blocks)
(65 : 35 : 5 , by vol.) or chloroform/methanol/propanol/0.25% KCl/ethyl acetate (25: 13:25:9:25, by vol.). The migration of phospholipids was checked with standard phospholipids and visualized with iodine vapor. The spots were scraped off and analyzed for their phosphate content. In the brain phospho- lipid extract, the phosphorus associated with phosphatidyl- choline, -ethanolamine, -serine and -inositol amounted to 32%, 27%, 9% and 3%, respectively, of the total phosphorus content (28 lmol phospholipid phosphate obtained from membranes corresponding to I g fresh tissue).
RESULTS
Choline-phosphate cytidylyltransferase subcellular distribution: comparison with that ofphosphatidyl- ethanolamine N-methyltransferase
The 1000 x g pellet exhibited a high level of activity, both for phosphatidylethanolamine N-methyltransferase and cho- line-phosphate cytidylyltransferase (Table 1). This is probably due to the presence of unbroken cells as shown by the low specific activity expressed per mass of protein. Separation of the 17000 x g pellet on a sucrose gradient showed that both activities were slightly higher in synaptosomes (0.8/1.2 M su- crose solutions interface) than in the other fractions.
Methyltransferase activity was mainly particulate and re- covered in the 17000 x g and 105000 x g pellets. In contrast, choline-phosphate cytidylyltransferase activity was found in both supernatant and pellet from the 105000 x g centrifuga- tion. Thus, cytidylyltransferase is associated with lower-den- sity particles and the cytosol. The values we obtained for phosphatidylethanolamine N-methyltransferase are in agree- ment with those previously reported [23 , 291.
For further studies, we have optimized the recovery of choline-phosphate cytidylyltransferase activity. Tissue was
Table 2. Stimulation of cytosolic choline-phosphate cytidylyltrunsferase uctivity 1105000 x g 15-h supernatant) ufter addition qf membranes, phospholipids or oleate Each assay, containing 100 pl supernatant prepared according to procedure (b) (1 10 pg protein obtained from 19 mg fresh tissue), was incubated for 30min at 37'.'C in the presence of 2 m M CTP and 2.8 FM labeled choline phosphate in buffer A. 10 p1 membrane sus- pension (105000 x g, 1 h pellet) containing 64 pg protein and 85 nmol phosphorus, was prepared from 2 mg rresh tissue. 10 pI phospholipid suspension (56 nmol phosphorus) was extracted from 2 mg fresh tis- sue according to Hirata's procedure. Oleate concentration was 200 pM. Supernatant activity in the absence of oleate was almost absent. Stimulation of cytosolic eholine-phosphate cytidylyltrans- ferase was calculated as follows. In expts 1 and 2, activity was mea- sured in the assay containing supernatant + membrane extract; mem- brane extract and supernatant activities were subtracted from this value. In expt 5, activity was measured in the assay containing super- natant + oleate; supernatant activity was subtracted from this value. In expt 6, activity was measured in the assay containing supernatant + oleate + membrane extract; membrane extract and supernatant activities were subtracted from this value
Expt Addition Increased activity
pmol/assay
1. Membranes 5 p1 2. 10 p1
3.5 & 0.42 6.3 t- 0.51
3. Phospholipid 5 pl 1.8 & 0.28 4. 10 d 3.4 & 0.32 5. Oleate 6.0 0.45
7.0 0.48 6. Oleate + membranes 10 p1
then prepared according to procedure (b) described in Methods. The 1000 x g supernatant was centrifuged at 105000 x g for various periods of time, in order to determine the influence of the centrifugation duration. We found (Fig. 1)
370
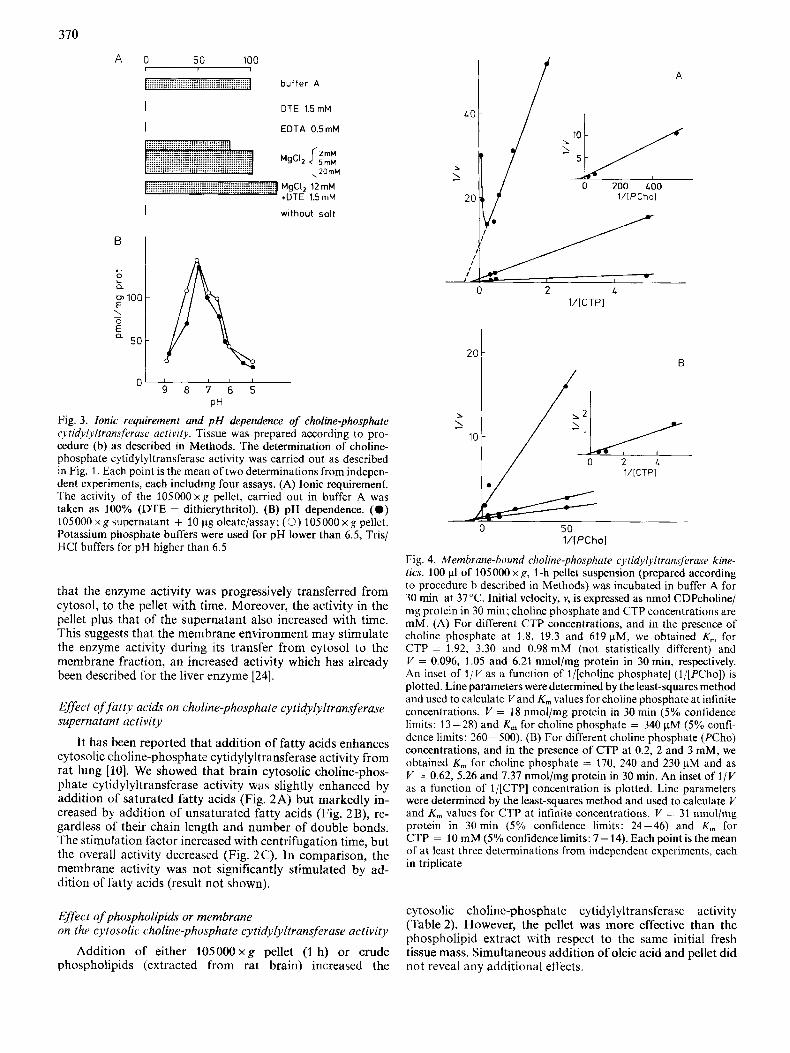
A 0 50 100 I , 1
b-ffer A A
DTE 1.5 mM
EDTA 0.5mM
B
c
L 0
a
MgCI, l2mM *DTE 1 5 m M
I wlthout salt
u 9 8 7 6 5
PH Fig. 3. Ionic requirement and p H dependence of choline-phosphate cytidylyltransferuse activity. Tissue was prepared according to pro- cedure (b) as described in Methods. The determination of choline- phosphate cytidylyltransferase activity was carricd out as described in Fig. 1. Each point is the mean of two determinations from indepcn- dent cxperiments, each including four assays. (A) Ionic requirement. The activity of the 105000xg pellet, carried out in buffer A was taken as 100% (DTE = dithierythritol). (B) pH dependence. (0 ) 105000xg supernatant + 10 pg oleate/assay; (0) 105000xg pellct. Potassium phosphate buffers were used for pH lower than 6.5, Tris/ HCI buffers for pH higher than 6.5
that the enzyme activity was progressively transferred from cytosol, to the pellet with time. Moreover, the activity in the pellet plus that of the supernatant also increased with time. This suggests that the membrane environment may stimulate the enzyme activity during its transfer from cytosol to the membrane fraction, an increased activity which has already been described for the liver enzyme [24].
Effect offatty acids on choline-phosphate cytidylyltransferuse supernutunt uctivity
It has been reported that addition of fatty acids enhances cytosolic choline-phosphate cytidylyltransferase activity from rat lung [lo]. We showed that brain cytosolic choline-phos- phate cytidylyltransferase activity was slightly enhanced by addition of saturated fatty acids (Fig. 2A) but markedly in- creased by addition of unsaturated fatty acids (Fig. 2B), re- gardless of their chain length and number of double bonds. The stimulation factor increased with centrifugation time, but the overall activity decreased (Fig. 2 C). In comparison, the membrane activity was not significantly stimulated by ad- dition of fatty acids (result not shown).
Ejfect ojphospholipidy or membrane on the cytosolic choline-phosphate cytidylyltransferase uclivity
Addition of either 105000xg pellet (1 h) or crude phospholipids (extracted from rat brain) increased the
20
b
I 10
2
B
0 2 1 l/[CTPI
a /
/&/-=L 50 l/IPChol
Fig. 4. Membrane-hound choline-phosphate cytidylyltransf~rase kine- tics. 100 p1 of 105000xg, 1-h pellet suspension (prepared according to procedure b described in Methods) was incubated in buffer A for 30 min at 37°C. Initial velocity, v, is exprcssed as nmol CDPcholine/ mg protein in 30 min; cholinc phosphate and CTP concentrations are mM. (A) For different CTP concentrations, and in the prcscnce of choline phosphate at 1.8, 19.3 and 619 pM, we obtained K, for CTP = 1.92, 3.30 and 0.98 mM (not statistically different) and V = 0.096, 1 .OS and 6.21 nmol/mg protein in 30 min, respectively. An inset of l/V as a function of l/[choline phosphate] (l/[PCho]) is plotted. Line parameters were determined by the least-squares method and used to calculate V and K, values for choline phosphate at inh i te concentrations. V = 18 nmol/mg protein in 30 min (5% confidence limits: 13-28) and K , for choline phosphate = 340 FM (5% confi- dcnce limits: 260- 500). (B) For different choline phosphate (PCho) concentrations, and in the presence of CTP at 0.2, 2 and 3 mM, we obtained K, for choline phosphate = 170, 240 and 230 pM and as V = 0.62, 5.26 and 7.37 nmol/mg protein in 30 min. An inset of 1 / V as a function of l/[CTP] concentration is plotted. Line parameters were determincd by the least-squares method and used to calculate V and K., values for CTP at infinite concentrations. I/ = 31 nmo\/mg protein in 30min (5% confidence limits: 24-46) and K , for CTP = 10 mM(5%confidcnceIimits:7-14). Eachpointis themean of at least three determinations from independent cxperiments, each in triplicate
cytosolic choline-phosphate cytidylyltransferase activity (Fable 2). However, the pellet was more effective than the phospholipid extract with respect to the same initial fresh tissue mass. Simultaneous addition of oleic acid and pellet did not reveal any additional effects.
371
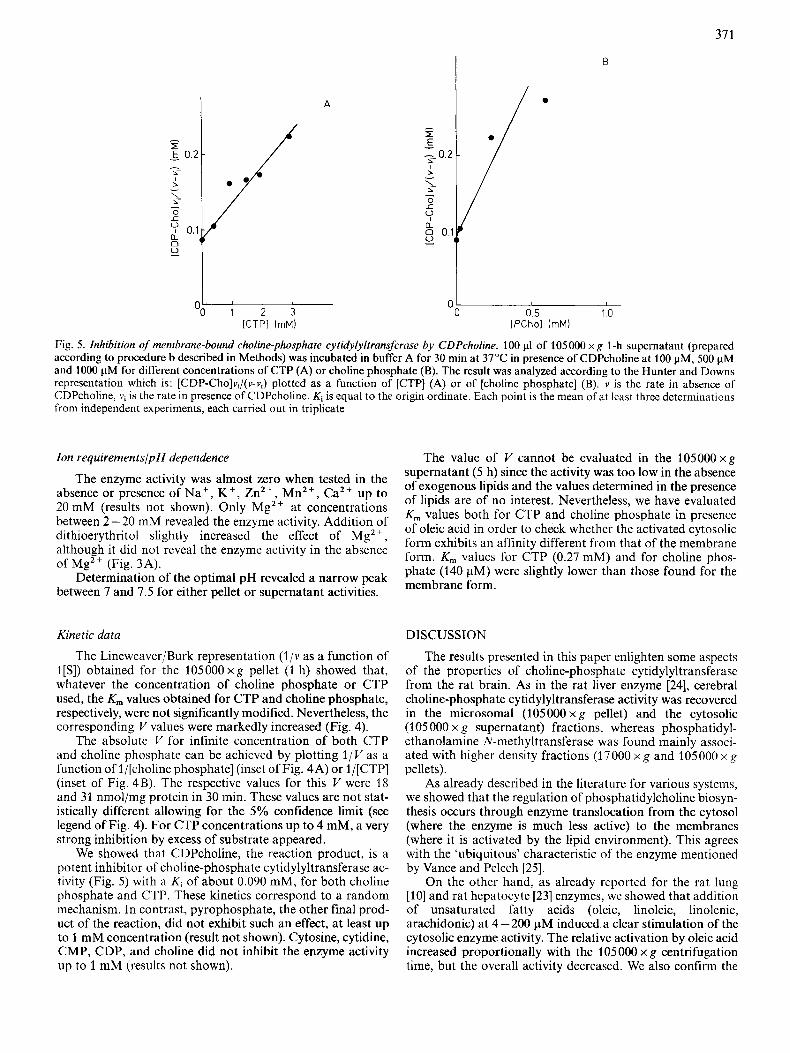
B
A
0- 0 0.5 1 0
[PChol (mM)
Fig. 5 . Inhibition of membrane-bound choline-phosphate cytidylyltransferase by CDPcholine. 100 p1 of 105 000 x g 1-h supernatant (prepared according to procedure b described in Methods) was incubated in buffer A for 30 min at 37°C in presence of CDPcholine at 100 pM, 500 pM and 1000 pM for different concentrations of CTP (A) or choline phosphate (B). The result was analyzed according to the Hunter and Downs representation which is: [CDP-Cho]vi/(v-vi) plotted as a function of [CTP] (A) or of [choline phosphate] (B). v is the rate in absence of CDPcholine, vi is the rate in presence of CDPcholine. Ki is equal to the origin ordinate. Each point is the mean of at least three determinations from independent experiments, each carried out in triplicate
Ion requirementslpH dependence The value of V cannot be evaluated in the 105000 X R supernatant (5 h) since the activity was too low in the absence
of lipids are of no interest. Nevertheless, we have evaluated K , values both for CTP and choline phosphate in presence of oleic acid in order to check whether the activated cytosolic
form. K, values for CTP (o.27 mM) and for cho,ine phos- phate (140 pM) were slightly lower than those found for the membrane form.
The enzyme activity was almost zero when tested in the of exogenous lipids and the values determined in the presence absence Or presence Of Nat ' K + ' Zn2+' Mn2+' up to 20 mM (results not shown). Only Mg2+ at concentrations between 2 - 20 mM revealed the enzyme activity. Addition of
although it did not reveal the enzyme activity in the absence of Mg2+ (Fig. 3A).
Determination of the optimal pH revealed a narrow peak between 7 and 7.5 for either pellet or supernatant activities.
dithioerythritol 'lightly increased the effect Of Mg2+, form exhibits an affinity different from that of the membrane
Kinetic data DISCUSSION
The Lineweaver/Burk representation ( l / v as a function of l[S]) obtained for the 105000xg pellet (1 h) showed that, whatever the concentration of choline phosphate or CTP used, the K, values obtained for CTP and choline phosphate, respectively, were not significantly modified. Nevertheless, the corresponding V values were markedly increased (Fig. 4).
The absolute V for infinite concentration of both CTP and choline phosphate can be achieved by plotting l / V as a function of l/[choline phosphate] (inset of Fig. 4A) or l/[CTP] (inset of Fig. 4B). The respective values for this V were 18 and 31 nmol/mg protein in 30 min. These values are not stat- istically different allowing for the 5% confidence limit (see legend of Fig. 4). For CTP concentrations up to 4 mM, a very strong inhibition by excess of substrate appeared.
We showed that CDPcholine, the reaction product, is a potent inhibitor of choline-phosphate cytidylyltransferase ac- tivity (Fig. 5) with a Ki of about 0.090 mM, for both choline phosphate and CTP. These kinetics correspond to a random mechanism. In contrast, pyrophosphate, the other final prod- uct of the reaction, did not exhibit such an effect, at least up to 1 mM concentration (result not shown). Cytosine, cytidine, CMP, CDP, and choline did not inhibit the enzyme activity up to 1 mM (results not shown).
The results presented in this paper enlighten some aspects of the properties of choline-phosphate cytidylyltransferase from the rat brain. As in the rat liver enzyme 1241, cerebral choline-phosphate cytidylyltransferase activity was recovered in the microsomal (105000 x g pellet) and the cytosolic (105 000 x g supernatant) fractions, whereas phosphatidyl- ethanolamine N-methyltransferase was found mainly associ- ated with higher density fractions (17000 x g and 105000 x g pellets).
As already described in the literature for various systems, we showed that the regulation of phosphatidylcholine biosyn- thesis occurs through enzyme translocation from the cytosol (where the enzyme is much less active) to the membranes (where it is activated by the lipid environment). This agrees with the 'ubiquitous' characteristic of the enzyme mentioned by Vance and Pelech [25].
On the other hand, as already reported for the rat lung [lo] and rat hepatocyte [23] enzymes, we showed that addition of unsaturated fatty acids (oleic, linoleic, linolenic, arachidonic) at 4-200 pM induced a clear stimulation of the cytosolic enzyme activity. The relative activation by oleic acid increased proportionally with the 105000 x g centrifugation time, but the overall activity decreased. We also confirm the

312
inability of saturated fatty acids (myristic, palmitic, stearic, arachidic) to stimulate the soluble cytidylyltransferase ac- tivity.
Although previous results on cultured rat hepatocytes [23] indicated that maximal stimulation of the cytosolic activity by oleate was lower than that obtained with total phospho- lipids, we found cerebral cytosolic activity to be more stimu- lated by oleate than by membrane crude phospholipids (Table 2). Moreover, since no additional effects of oleate and membrane phospholipids could be observed, this indicates that the maximal stimulation could be achieved in the presence of either lipid.
The phospholipid extract does not seem to be the main membrane stimulating agent, since its activating potential was lower than that of corresponding membranes. An alternative explanation would be that membrane integrity is required for the total expression of the phospholipid stimulating effect.
The kinetic properties of the brain enzyme that we found were similar to those obtained from other tissues [7, 141: neutral optimal pH, K , for choline phosphate at about 0.3 mM and K,,, for CTP btween 2 - 10 mM. CDPcholine, one of the reaction products, was a potent inhibitor, competing for both CTP and choline phosphate with a Ki near 0.090 mM.
In conclusion, the present work has allowed the brain choline-phosphate cytidylyltransferase to be characterized. Its properties are very similar to those of the enzyme obtained from other sources (liver, lung, etc.). Besides its fundamental interest, knowledge of the properties of the brain enzyme is important from the pharmacological point of view since it is assumed that it plays an essential role in brain disorders.
First, various very frequent diseases, such as ischemia [17 -191, hypoxia [20, 261 and ageing [17, 271, induce brain tissue modifications which could affect the enzyme via an increase of free fatty acids, coupled with destruction of mem- brane phospholipids, especially phosphatidylethanolamine and phosphatidylcholine [28]. Second, in most of these dis- eases, administration of CDPcholine and CDPethanolamine can reverse the process [18]. This makes it likely that choline- phosphate cytidylyltransferase, the key enzyme of phospha- tidylcholine biosynthesis [25], plays a significant role in brain disorders.
We want to thank Dr D. Gautheron (University Lyon 1) and Dr D. Vance (University of Alberta) for thcir helpful advice in the interpretation of the results and their critical reading of the manu- script.
REFERENCES 1. Kennedy, E. & Weiss, S. (1956) J . Biol. Chem. 222, 193-214. 2. Lee, T., Austen, F., Corey, J. & Drazen, M. (1984) Proc. Nut/
Acad. Sci. U S A 81,4922-4925.
3. Hirdta, F., Strittmatter. W. & Axelrod, J. (1979) Proc. Nut1 Acad.
4. Di Perri, B.. Calderini, G., Battistella, A., Raciti, R. & Toffano,
5. Infante. J. P. & Kinsella, J. E. (1978) Biochim. Biophys. Acta 526,
6. Postle. A. D.. Hunt, A. N. & Normand, I. C . S. (1985) The physiological development of the fetus and newborn, pp. 305 - 309, Academic Press, London.
7. Choy, P. C., Lim, P. H. & Vance, D. E. (1977) J . Biol. Chem. 252,
8. Pelech, S. L., Cook, H. W., Paddon, H. B. & Vance, D. E. (1984) Biochim. Biophys. Acta 795, 433 -440.
9. Lim, P. H., Pritchard, P. H., Paddon, H. B. & Vance, D. E. (1983) Biochim. Biophys. Acta 753, 74-82.
10. Weinhold, P. A,, Rounsifer, M. E., Williams, S. E., Brubaker, P. G.& Feldman, D. A. (1984)J. Biol. Chem.259,10315-10321.
11. Feldman, D. A., Kovac, C., Dranginis, P. & Weinhold, P. A. (1978) J . Biol. Chem. 253,4980-4986.
12. Feldman, D. A., Dietrich, J. W. & Weinhold, P. A. (1980) Bio- chim. Biophys. Acta 620, 603-61 1.
13. Stem, W., Kovac, C. & Weinhold, P. A. (1976) Biochim. Biophys. Acta 441, 280-293.
14. Freysz, L., Binaglia, L., Dreyfus, H., Massarelli, R., Golly, F. & Porcellati, G. (1985) J . Neurochem. 45, 57-62.
15. Arienti, G., Corazzi, L., Freysz, L., Binaglia, L., Roberti, R. & Porcellati, G. (1985) J . Neurochem. 44, 38-41.
16. Trovarelli, G., Palmerini, C. A,, Floridi, A., Piccinin, G. L. & Porcellati, G. (1984) Neurochem. Res. 9, 73-79.
17. De Medio, G. E., Brunetti, M., Dorman, R. V., Droz, B., Horrocks, L. A,, Porcellati, G., Souyri, F. & Trovarelli, G. (1983) Birth Defects Orig. Artic. Ser. 19, 175-187.
18. Dorman, R. V., Dabrowiecki, Z. & Horrocks, L. A. (1983) J . Neurochem. 40,276 - 279.
19. Trovarelli, G., De Medio, G. E., Dorman, R. V., Piccinin, G. L., Horrocks, L. A. & Porcellati, G. (1981) Neurochern. Res. 6,
20. Alberghina, M., Viola, M. & Giuffrida, A. M. (1981) J . Neurosci.
21. De Medio, G. E., Trovarelli, G., Piccinin, G. L. & Porcellati, G.
22. Gray, E. & Whittaker, V. (1962) J. Anat. 96, 79-91. 23. Fonlupt, P., Rey, C. & Pacheco, H. (1981) Biochem. Biophys. Res.
Commun. 100, 1720 - 1726. 24. Pelech, S. L., Pritchard, H. W., Brindley, D. N. & Vance, D. E.
(1983) J . Biol. Chern. 258,6782-6788. 25. Vance, D. E. & Pelech, S. L. ( 1 984) Trends Biochem. Sci. 9, 17 -
20. 26. Alberghina, M., Viola, M., Serra, I., Mistretta, A. & Giuffrida,
A. M. (1981) J . Neurosci. Res. 6,421-433. 27. Porcellati, G., Gaiti, A. & Brunetti, M. (1982) The aging brain:
cellular and molecular mechanism of aging in the nervous system (Giacobini, E., Filogamo, G., Giacobini, G. & Vernadakis, A., eds) pp. 77 - 86, Raven Press, New York.
28. Goracci, G., Francescangeli, E., Horrocks, L. A. & Porcellati, G. (1981) Biochim. Biophys. Acta 664, 373-379.
29. Crews, F., Hirata, F. & Axelrod, J . (1980) J . Neurochem. 34, 1491 - 1498.
Sci. USA 76, 368-372.
G. (1983) J . Neurochem. 41, 302-308.
440 - 449.
7673 - 7677.
821 -823.
R ~ s . 6, 719-722.
(1984) J . Neurosci. Res. 11, 49-58.





























