Embed Size (px)
Citation preview

Article original
Mécanismes de la protection cardiaque et vasculaire des polyphénolsau niveau de l’endothélium
Cellular mechanism of vasculo-protection induced by polyphenolson the endothelium
S. Martin, R. Andriantsitohaina *
Pharmacologie et physico-chimie des interactions cellulaires et moléculaires, UMR CNRS 7034, faculté de pharmacie, université Louis-Pasteur,74, route du Rhin, 67401 Illkirch, France
Reçu le 23 septembre 2002; accepté le 23 septembre 2002
Résumé
Des études épidémiologiques ont établi une corrélation entre la consommation de polyphénols végétaux et une faible incidence demaladies coronariennes, probablement par leurs capacités à modifier de nombreux facteurs à l’origine de ces maladies. Certains polyphénolset notamment ceux du vin rouge possèdent des propriétés anti-oxydantes et captatrices de radicaux libres. Ils protègent les « lipoprotéinesde faibles densités » contre l’oxydation. Par ailleurs, les polyphénols diminuent l’agrégation des plaquettes et sont capables d’inhiber laprolifération des cellules musculaires lisses vasculaires. Au niveau de l’endothélium vasculaire, les polyphénols stimulent la production defacteurs relaxants comme le monoxyde d’azote. Enfin, ils contribuent à préserver l’intégrité de l’endothélium vasculaire en agissant à la foissur la prolifération, la migration et l’apoptose des cellules endothéliales. Grâce à ces propriétés, les polyphénols pourraient interférer avecle processus athérogène et/ou les phénomènes thrombotiques associés à l’athérosclérose et pourraient expliquer les effets vasculo- etcardio-protecteurs de ces composés. © 2002 E´ditions scientifiques et médicales Elsevier SAS. Tous droits réservés.
Abstract
Epidemiological studies have suggested that dietary factors, including moderate red wine consumption, might reduce the risk ofcardiovascular diseases. The beneficial effect of fruits, vegetables, or red wine may be in part explained by the presence of polyphenols witha multitude of biological activities, including antioxidant and free radical-scavenging properties, anti-aggregatory platelet property andinhibition of vascular smooth muscle cell proliferation. Another therapeutically relevant effect of polyphenols may be their ability to interactwith the generation of nitric oxide from vascular endothelium that leads not only to vasodilatation but also to the expression of genesprotective of the cardiovascular system. Finally, polyphenols contribute to the preservation of endothelial integrity by acting on theprocesses implicated in endothelial proliferation, migration and apoptosis. All these effects of polyphenols might interfere withatherosclerotic plaque development and stability, vascular thrombosis and occlusion and therefore might explain their cardio- and vascularprotective properties. © 2002 E´ditions scientifiques et médicales Elsevier SAS. All rights reserved.
Mots clés: Remodelage vasculaire; Prolifération; Migration; Apoptose; Vin rouge
Keywords: Vascular remodeling; Proliferation; Migration; Apoptosis; Red wine
1. Historique
Dès les années 1970, de grandes études épidémiologi-ques ont révélé une réduction considérable du risque de
maladies cardiovasculaires chez les consommateurs modé-rés d’alcool. Même si les effets cardioprotecteurs apportéspar cette consommation étaient connus de longue date parles chercheurs, c’est l’étude de St-Léger, publiée en 1979,qui a marqué le début de l’essor considérable des recherchesdans le domaine du vin[1]. Dans cette étude, St-Légermontre une relation inverse entre la consommation
* Auteur correspondant.Adresse e-mail : [email protected] (R. Andriantsitohaina).
Annales de cardiologie et d’angéiologie 51 (2002) 304–315
www.elsevier.com/locate/ancaan
© 2002 Éditions scientifiques et médicales Elsevier SAS. Tous droits réservés.PII: S 0 0 0 3 - 3 9 2 8 ( 0 2 ) 0 0 1 3 8 - 5

moyenne de vin dans 18 pays et la mortalité provoquée parles maladies cardiovasculaires. De nombreuses études pos-térieures ont confirmé cette observation. C’est à travers uneétude réalisée à partir de données recueillies par l’organi-sation mondiale de la santé (OMS), le programme WHO-Monica (World Health Organization - Monitoring Cardio-vascular Disease) (WHO-Monica project, 1988), quel’épidémiologiste Serge Renaud a mis en évidence le trèscontroversé «paradoxe français », terme utilisépour décrirela situation apparemment paradoxale dans laquelle, avec desfacteurs de risque identiques (tabagisme, cholestérolémie oumanque d’exercice physique), les Français, notamment ceuxdu Sud, meurent moins de maladies cardiovasculaires parrapport àd’autres populations, notamment celles du Nord del’Europe et des États-Unis. Même si le projet international« WHO-Monica » (1980-1990) et les projets européens« Euroaspire I et II (1995-2000) » confirment de plus enplus la réalité du « paradoxe français », ils mettent égale-ment en évidence que son assise géographique dépasse leslimites de l’hexagone. En fait, il s’agirait plutôt d’unparadoxe « méditerranéen ».
Actuellement, deux grandes études montrent qu’uneconsommation modérée de vin réduit la mortalité, toutescauses de maladies confondues. Une étude danoise avec unsuivi d’une cohorte pendant 15 ans a montré qu’uneconsommation de 3 à 5 verres de vin par jour est associée àune diminution de 50 % du risque de mortalité [2]. En 1998,dans l’étude de Nancy, l’équipe du Dr S. Renaud et al. (sur34 000 personnes suivies pendant 10 à 15 ans dans l’Est dela France) observe une réduction de 30 % de la mortalité,chez les consommateurs modérés de vin (1 à 3 verres parjour soit 150 à 450 ml/j) par rapport aux abstinents [3]. Lesdeux études conduisent à la même conclusion : si laréduction du risque de mortalité liée aux maladies cardio-vasculaires est significative (30 %), elle l’est égalementpour les autres maladies, parmi lesquelles les cancers(22 %).
Encore fallait-il que ces observations soient confortéespar des preuves biologiques. Celles-ci ont été apportées parle biais d’études fondamentales, dont certaines, commecelles du Pr Jack Masquelier, remontent déjà à plusieursdécennies [4]. Le rôle cardioprotecteur de la consommationmodérée du vin découle de plusieurs mécanismes comple-xes même s’ ils ont d’abord été attribués à l’alcool. Il a étédémontré que l’alcool contribue à élever le taux deslipoprotéines de haute densité (HDL) et àdiminuer celui deslipoprotéines de basse densité (LDL) [5], ces deux actionspermettant de protéger l’organisme contre le développementde l’athérosclérose. Par ailleurs, l’alcool inhibe l’agrégationdes plaquettes sanguines, réduit la teneur en fibrinogène etaccélère la fibrinolyse, ou la dissolution du caillot sanguin,à l’origine d’accidents vasculaires ischémiques [6]. Cepen-dant les effets, communs à l’ensemble des boissons alcoo-lisées, ne pouvaient expliquer les résultats obtenus par denombreuses études indiquant une protection supérieureobservée par la consommation de vins rouges par rapport
aux vins blancs ou aux autres boissons alcoolisées. Ladifférence majeure entre ces différentes boissons est leurcomposition en polyphénols. En effet, les vins rouges ontune teneur élevée en composés phénoliques, par rapport auxvins blancs et la bière [7].
Ces dernières années, notre laboratoire s’est intéressé detrès près aux effets cardiaques et vasculaires des polyphé-nols contenus dans le vin rouge [8-14]. Les études, réaliséesin vivo et in vitro chez le rat, ont permis de mettre enévidence des effets vasodilatateurs à court terme, des effetshypotenseurs, des effets protecteurs contre l’ ischémie car-diaque et contre la fibrose myocardique à long terme despolyphénols. L’ensemble de ce travail a montré le rôleprédominant de l’endothélium et en particulier du mo-noxyde d’azote (NO) en tant que médiateur des effetsvasculaires des polyphénols.
2. Endothélium vasculaire
Depuis la découverte de Furchgott et Zawadzki [15],plusieurs études ont montré que l’endothélium n’était pasune simple barrière entre le sang circulant et les couchesvasculaires sous-jacentes. En effet, l’endothélium vasculaireest impliqué à la fois dans la régulation physiologique dutonus vasculaire, dans la régulation de l’homéostasie vas-culaire et dans les transformations structurales des vais-seaux prenant place lors d’évolutions pathologiques [16].Longtemps considéré comme une simple barrière passive,l’endothélium occupe en fait une position stratégique,localisant idéalement les cellules endothéliales à l’ interfacesang/tissu, et leur permettant ainsi d’affecter à la fois lafonction des cellules sanguines et celle des cellules consti-tuant la paroi vasculaire. Du fait de sa localisation, il est aucontact de divers facteurs circulants tels que des agonistes,(comme l’acétylcholine ou l’histamine), des cytokines pro-inflammatoires comme le facteur de nécrose tumoral (TNF)ou les interleukines (IL) ou des macromolécules circulantescomme les LDL oxydées, les antioxydants ou le glucose,mais également des forces de cisaillement, exercées par desvariations de débit du flux sanguin, ou d’étirement, liées àdes modifications de la pression sanguine [16,17]. Auniveau des cellules endothéliales, ces différents facteurspeuvent [1] moduler la motricité vasculaire, [2] induire laproduction de facteurs endothéliaux, notamment le NO [18],qui interviennent dans de nombreuses fonctions physiologi-ques et [3] moduler des processus cellulaires telles quel’apoptose, la prolifération et la migration des cellulesendothéliales [19-21].
L’altération de ces processus physiologiques est souventla cause du développement de diverses pathologies ; à titred’exemple, la perte de l’activitébiologique et/ou l’altérationde la production de NO. Le NO libéré par l’endothéliumvasculaire est synthétisé à partir de la L-arginine sousl’action d’une enzyme, la NO-synthase (eNOS). Le NOdiffuse vers les cellules musculaires lisses vasculaires,active différentes cibles intracellulaires, notamment la gua-
S. Martin, R. Andriantsitohaina / Annales de cardiologie et d’angéiologie 51 (2002) 304–315 305

nylate cyclase soluble, pour entraîner l’augmentation dutaux de guanosine-3’ ,5’ -cyclique monophosphate (GMPc).Le GMPc induit le phénomène de relaxation vasculaireprincipalement en activant une protéine kinase spécifique, laprotéine kinase G.
2.1. Conséquences physiopathologiques de la pertede l’activité biologique et/ou l’altération de la productionde NO
Compte tenu des effets multiples du NO, l’altération dufonctionnement de la voie du NO induit :
• la transformation de l’endothélium de surface anticoa-gulante en surface procoagulante, puisque le NOn’exerce plus ses effets anti-agrégants plaquettaires etn’ inhibe plus l’expression de récepteurs tels que laE-sélectine, la P-sélectine, ICAM-1 ou VCAM-1, inter-venant dans l’adhésion des plaquettes et des leucocytes[22,23] ;
• l’augmentation de la perméabilité de l’endothéliumfacilitant l’ infiltration et le dépôt des lipides, desmonocytes et des cellules musculaires lisses dansl’ intima, contribuant au développement de lésions vas-culaires comme l’athérosclérose [22,24] ;
• la prolifération des cellules musculaires lisses condui-sant à l’épaississement de la paroi vasculaire et audéveloppement de pathologies vasculaires occlusivestelles que la resténose ou l’athérosclérose [11,25] ;
• l’altération du relâchement dépendant de l’endothé-lium. Un tel phénomène est observé dans des patholo-gies telles que l’hypertension artérielle, l’athérosclé-rose, l’hypercholestérolémie, le diabète ou desévolutions physiologiques tel que le vieillissementvasculaire.
La perte de l’activité biologique et/ou l’altération de laproduction de NO [26] peut être due à des altérations de latransduction du signal dans les cellules endothéliales, à unediminution de la disponibilité de la L-arginine ou descofacteurs de la eNOS (tétrahydrobioptérine), à une modi-fication de l’expression de la eNOS ou à une augmentationde la destruction du NO par les espèces réactives del’oxygène.
Les traitements des pathologies associées àune altérationde la voie du NO visent actuellement à restaurer uneproduction physiologique de NO. Elle est réalisée par uneadministration systémique de L-arginine [27,28] ou dedonneurs de NO (molsidomine, S-nitrosothiols ou la3-morpholinosydnonimine) [29,30]. Le NO peut aussi êtredélivré localement à l’aide de transporteur de NO(PROLI/NO ou SPER/NO, où le NO est transporté par laproline ou la spermine) [31,32] ou sous forme de pro-drogues (diazeniumdiolates) contenues dans des polymèresinsolubles ou des microsphères [33-35]. En chirurgie, l’uti-lisation de stent et de prothèses synthétiques recouvertes de
NO est de plus en plus courante [36]. À plus long terme, larestauration de l’expression de la eNOS par thérapie géni-que est envisagée [37]. L’ensemble des traitements viseainsi à empêcher la thrombose et la resténose après angio-plastie, à augmenter le contenu tissulaire en NO et à rétablirla fonction endothéliale. Il ne faut pas négliger l’utilisationd’anti-inflammatoires, d’antioxydants et d’œstrogènes quis’avèrent très efficaces pour activer la production de NOet/ou d’empêcher la dégradation du NO. L’endothéliumconstitue une nouvelle cible thérapeutique pour le traite-ment de nombreuses pathologies.
3. Les polyphénols, substances naturelles protectricesde l’endothélium ?
3.1. Présentation des polyphénols
3.1.1. Structure et classification des polyphénols
Les polyphénols constituent le groupe de métabolites leplus large et le plus répandu du règne végétal et font partieintégrante de l’alimentation humaine et animale. On lesretrouve en grande quantité dans le vin rouge où ils ont uneimportance particulière puisqu’ ils sont responsables de leurscaractéristiques gustatives et de leur qualité. Leur concen-tration est beaucoup moins importante dans les vins blancs.Les polyphénols sont apportés par les fruits (peaux etgrains) et les pieds de vigne, par le métabolisme des levureset par le bois des tonneaux.
Plus de 8000 structures phénoliques sont actuellementconnues, allant de molécules phénoliques simples de baspoids moléculaire tels les acides phénoliques à des compo-sés hautement polymérisés comme les tannins. Ils peuventêtre conjugués avec un ou plusieurs résidu(s) sucré(s) lié(s)ou ils peuvent également être liés avec d’autres composéschimiques, tels que des acides carboxyliques, des amines oudes lipides ou avec d’autres phénols existent également.
Les polyphénols sont des produits du métabolisme secon-daire des plantes. Ils sont synthétisés à partir de deux voiesbio-synthétiques :
• celle de l’acide shikimique, qui conduit après transa-mination et désamination, aux acides cinnamiques et àleurs nombreux dérivés tels que les acides benzoiquesou les phénols simples ;
• celle issue de l’acétate, qui conduit à des polyß-coesters (polyacétates) de longueur variable menantpar cyclisation à des composés polycycliques tels queles dihydroxy-1,8 anthraquinones ou les naphtoquino-nes.
De plus, la diversité structurale des composés polyphé-noliques, due à cette double origine bio-synthétique estencore accrue par la possibilité d’une participation simultanée des deux voies dans l’élaboration de composés d’ori-gine mixte, les flavonoïdes.
306 S. Martin, R. Andriantsitohaina / Annales de cardiologie et d’angéiologie 51 (2002) 304–315

3.1.2. Les polyphénols dans l’alimentation courante
3.1.2.1. Exemples de sources riches en polyphénols. Lespolyphénols sont présents dans de nombreux alimentsd’origine végétale, tels que les légumes, les fruits, lescéréales ou les fruits secs, ainsi que dans les boissons,comme le vin, le cidre, la bière ou le thé. En ce qui concerneles anthocyanes, des quantités importantes de glycosides demalvidine sont contenues dans les raisins et les produitsdérivés [38]. Les diglycosides de delphinidine sont lesanthocyanes majoritairement présentes dans la grenade [39].Ce fruit possède d’ailleurs une grande richesse en antho-cyanes puisqu’ il contient également des glycosides dedelphinidine, de cyanidine et de pelgargoninidine et desdiglycosides de cyanidine et de pelgargoninidine. Le vinrouge est parmi les sources alimentaires les plus riches enpolyphénols, puisque le taux de polyphénols s’élève à plusde 1 g/l. Leur quantité diminue significativement dans lesvins blancs où ils sont estimés être à 50 mg/l. Cependant, lateneur en anthocyanes est très variable selon l’origine, lecépage, les conditions de culture de la vigne et le mode devinification entrant dans sa fabrication. Il est, de ce fait, trèsdifficile d’évaluer la composition exacte d’un aliment, etd’autant plus du vin rouge (formé d’un vaste groupe destructures chimiques variées) en anthocyanes. Et bien quedifférentes techniques d’analyses soient utilisées actuelle-ment, de nombreuses incertitudes persistent encore.
3.1.2.2. Consommation quotidienne en polyphénols. Ac-tuellement, peu d’ informations sont disponibles sur laconsommation quotidienne de polyphénols végétaux dansl’alimentation. Une étude réalisée aux États-Unis a évaluéune consommation quotidienne de 1 g/j de flavonoïdes [40].Les plus grandes quantités de flavonoïdes ingérées provien-nent des boissons et des fruits (420 mg par le café, le thé, levin, le chocolat et la bière et 290 mg par les fruits et les jus defruit) [41]. Une étude plus récente, réalisée aux Pays-Bas, aestimé la consommation moyenne quotidienne de flavonolset de flavones à23 mg [42,43]. Ces résultats sont similaires àceux rapportés par Justesen [44], qui a estimé la consomma-tion de flavonoïdes au Dannemark à28 mg/j. La consomma-tion quotidienne d’anthocyanes aux États-Unis a été évaluéeà180–215 mg/j [40]. Des extraits riches en anthocyanes, telsque les extraits de myrtille et de sureau, sont disponibles dansle commerce. Ainsi, s’ ils sont utilisés comme complémentnutritionnel, les apports journaliers en anthocyanes sontconsidérablement augmentés. C’est pourquoi, il est encoredifficile d’évaluer la consommation totale de polyphénols etplus particulièrement d’anthocyanes dans l’alimentation.
3.1.2.3. Bio-disponibilité des polyphénols. On appelle bio-disponibilité d’une substance la proportion qui parvient à lacirculation générale après une administration orale ainsi quesa métabolisation, sa distribution et son élimination. Ainsi,il est important de connaître non seulement la quantitéjournalière ingérée d’anthocyanes, mais également leur
bio-disponibilité, puisque leur qualité nutritionnelle et leurseffets systémiques dépendront de leur absorption au niveaudu tractus digestif.
L’absorption, la distribution et la métabolisation desanthocyanes n’ont que peu été étudiées. Il est donc difficilede calculer les paramètres pharmacocinétiques des polyphé-nols et plus particulièrement des anthocyanes absorbéesquotidiennement dans l’alimentation, notamment en raisonde la complexité des structures chimiques de ces composés.Un dosage des flavonoïdes dans le plasma humain de sujetsn’ayant pas eu d’apport supplémentaire en polyphénols apermis de révéler la présence de ces composés à uneconcentration comprise entre 0,5 et 1,6 µM [45]. Les étudesde Matsumoto et al. [46] ont déterminé les paramètresd’absorption de la delphinidine-3-ruténoside (D3R), de lacyanidine-3-ruténoside (C3R), de la delphinidine-3-glucoside (D3G) et de la cyanidine-3-glucoside (C3G)administrées oralement à des rats (800 µmol/kg de poids).Ils ont mis en évidence une concentration plasmatiquemaximale (Cmax) d’anthocyanes de l’ordre de 580 ± 410,850 ± 120 et de 840 ± 190 nM deux heures après leuradministration pour la D3R, la C3R et la C3G respective-ment. Une étude a été réalisée chez l’Homme avec ingestiond’un mélange de 2,75 µM (1,68 mg) de D3R, 2,08 µM(1,24 mg) de C3R, 1,04 µM (0,488 mg) de D3G et 0,37 µM(0,165 mg) de C3G. Les résultats montrent que les quatreanthocyanes sont présentes dans le plasma et dans l’urinedes sujets ayant absorbé ce mélange. La Cmax atteintrespectivement 73,4 ± 35,0, 46,3 ± 22,5, 22,7 ± 12,4 et5,0 ± 3,7 nM pour D3R, la C3R, la D3G et la C3G deuxheures suivant la prise. Ces résultats suggèrent que lesanthocyanes sont directement absorbées et distribuées dansle sang [46].
3.2. Effets vasculaires des polyphénols
La consommation d’aliments riches en polyphénols réduitle développement de nombreuses pathologies, telles que lecancer, l’ ischémie cardiaque, l’athérosclérose et l’hyperten-sion [42,43, 47]. Cela peut-être expliqué par le fait que cescomposés ont la capacité de modifier de nombreux facteursimpliqués dans la genèse de ces maladies. Les polyphénolssont en effet capables d’abaisser la pression artérielle chez lerat [11,12], d’empêcher l’oxydation des LDL [48], d’ inhiberla prolifération des cellules musculaires lisses vasculaires[49], d’empêcher l’agrégation plaquettaire [50], de stabiliserles cellules immunitaires et de promouvoir le relâchementdes cellules musculaires lisses vasculaires [8]. Ils ont ainsiétédécrits comme étant des antioxydants, des anti-agrégantsplaquettaires, des anti-inflammatoires, des anti-allergènes,des anti-thrombotiques et des anti-tumoraux [47] (Fig. 1).Leurs propriétés sont liées au fait qu’ ils peuvent modulerl’activitéde nombreuses protéines intracellulaires (les protéi-nes kinases, les phospholipases, l’adénylate cyclase, lesATPases, les cyclo-oxygénases (COX), les NOS ou lecytochrome P450) et agir sur différents types cellulaires (les
S. Martin, R. Andriantsitohaina / Annales de cardiologie et d’angéiologie 51 (2002) 304–315 307
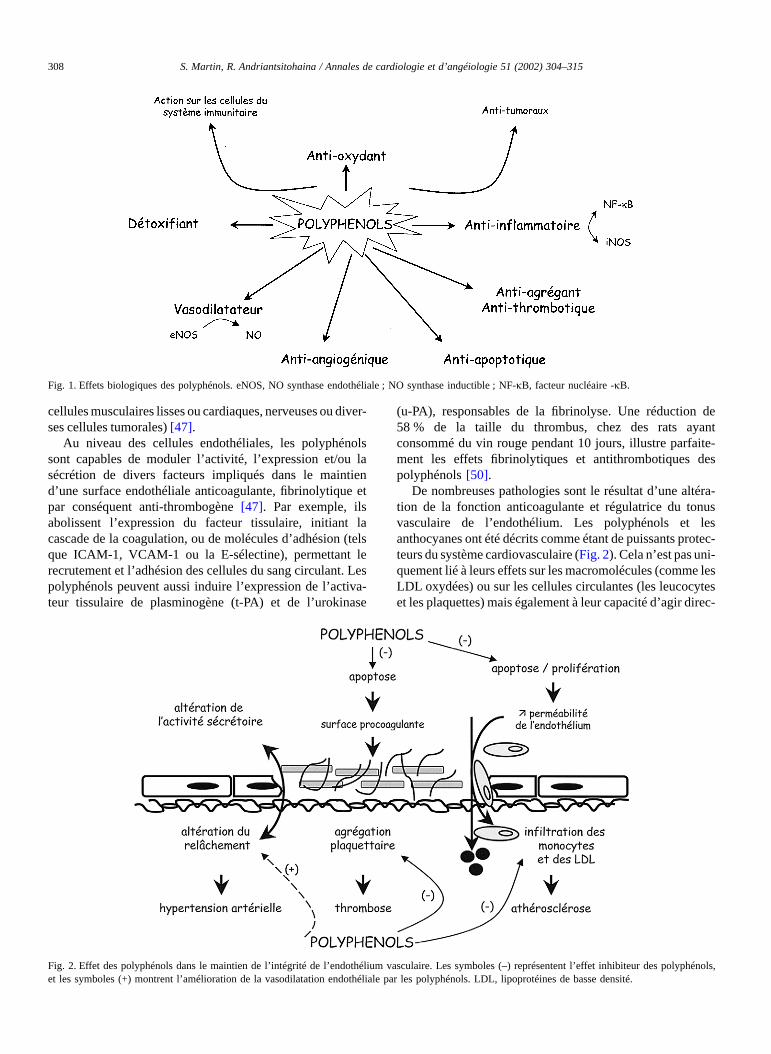
cellules musculaires lisses ou cardiaques, nerveuses ou diver-ses cellules tumorales) [47].
Au niveau des cellules endothéliales, les polyphénolssont capables de moduler l’activité, l’expression et/ou lasécrétion de divers facteurs impliqués dans le maintiend’une surface endothéliale anticoagulante, fibrinolytique etpar conséquent anti-thrombogène [47]. Par exemple, ilsabolissent l’expression du facteur tissulaire, initiant lacascade de la coagulation, ou de molécules d’adhésion (telsque ICAM-1, VCAM-1 ou la E-sélectine), permettant lerecrutement et l’adhésion des cellules du sang circulant. Lespolyphénols peuvent aussi induire l’expression de l’activa-teur tissulaire de plasminogène (t-PA) et de l’urokinase
(u-PA), responsables de la fibrinolyse. Une réduction de58 % de la taille du thrombus, chez des rats ayantconsommé du vin rouge pendant 10 jours, illustre parfaite-ment les effets fibrinolytiques et antithrombotiques despolyphénols [50].
De nombreuses pathologies sont le résultat d’une altéra-tion de la fonction anticoagulante et régulatrice du tonusvasculaire de l’endothélium. Les polyphénols et lesanthocyanes ont étédécrits comme étant de puissants protec-teurs du système cardiovasculaire (Fig. 2). Cela n’est pas uni-quement lié àleurs effets sur les macromolécules (comme lesLDL oxydées) ou sur les cellules circulantes (les leucocyteset les plaquettes) mais également à leur capacitéd’agir direc-
Fig. 1. Effets biologiques des polyphénols. eNOS, NO synthase endothéliale ; NO synthase inductible ; NF-jB, facteur nucléaire -jB.
Fig. 2. Effet des polyphénols dans le maintien de l’ intégrité de l’endothélium vasculaire. Les symboles (–) représentent l’effet inhibiteur des polyphénols,et les symboles (+) montrent l’amélioration de la vasodilatation endothéliale par les polyphénols. LDL, lipoprotéines de basse densité.
308 S. Martin, R. Andriantsitohaina / Annales de cardiologie et d’angéiologie 51 (2002) 304–315

tement sur l’endothélium vasculaire. Ainsi, la partie suivanteregroupe les informations concernant les effets spécifiquesdes polyphénols sur l’endothélium vasculaire et principale-ment sur les différents paramètres endothéliaux impliquésdans le contrôle de la vasomotricitépar l’endothélium. Nousévoquerons également les effets connus des polyphénols surla migration, la prolifération et l’apoptose des cellulesendothéliales qui sont des processus impliqués dans leremodelage vasculaire et l’angiogenèse. Cela permettra demieux appréhender les raisons pour lesquelles des propriétésvasculoprotectrices ont étéattribuées àces composés.
3.2.1. Effets des polyphénols sur le contrôlede la vasomotricité par l’endothélium
Une fonction physiologique importante de l’endothéliumest de moduler la motricité vasculaire. En effet, de nom-breuses pathologies sont associées à un dysfonctionnementde l’endothélium reflété par une altération du relâchementdu vaisseau sanguin. De nombreuses études montrent qu’ invitro, les polyphénols induisent à faible concentration(< 10–2 g/l) une vasodilatation dépendante de l’endothé-lium dépendante et à forte concentration (>10–1 g/l), unevasodilatation dépendante du muscle lisse. Ces études ontété réalisées tant avec les vins rouges, les vins blancs [51],qu’avec des extraits polyphénoliques et des composéspolyphénoliques purs [8,9]. De nombreuses études ont tentéd’ isoler LE composé polyphénolique responsable du relâ-chement vasculaire. Les études menées dans notre labora-toire ont montré que les anthocyanes et les tannins conden-sés de faible poids moléculaire sont les composés les plusefficaces pour induire un relâchement dépendant de l’endo-thélium. Le travail de Burns [52] a également montré queseules les anthocyanes présentent une activité vasodilatatrice dépendante de l’endothélium. Et, parmi la classe desanthocyanes, seule la delphinidine entraîne un relaxationvasculaire de même efficacitéque celle produite par l’extraitpolyphénolique de vin total [8]. En revanche, les anthocya-nes de structure comparable, telles que la malvidine et lacyanidine, ne relâchent pas le vaisseau [8]. Inversement, lamyricétine, un biflavonide, possède un effet contractiledépendant de l’endothélium [53].
Les effets des polyphénols sur la vasomotricité sont lereflet de leur action sur l’endothélium ou directement sur lescellules musculaires lisses. Cette partie vise essentiellementàdéterminer les effets des polyphénols sur les facteurs et lesvoies de signalisation impliqués dans la régulation de lavasomotricité par les cellules endothéliales.
3.2.1.1. Polyphénols et facteur relaxant : le monoxyded’azote (NO). De nombreuses études réalisées sur des vais-seaux isolés comme l’aorte ou l’artère mésentérique de ratmontrent que des extraits concentrés de polyphénols ou descomposés polyphénoliques purs induisent une relaxation desvaisseaux dépendante de la présence de l’endothélium [8,54].Il a été montré que l’action des polyphénols sur l’endo-thélium vasculaire est due àla production du NO, cela en me-
surant directement sa production par les expériences de réso-nance paramagnétique électronique [8]. Le dosage sanguinde NO réalisé chez l’Homme a montré que 30 min après laconsommation de vin rouge et de polyphénols (1 g/kg depoids), le taux de NO augmente de 30 et 40 nM respective-ment. Parallèlement, une réduction de la pression sanguinede 11 mmHg et une augmentation du rythme cardiaque sontobservées [55]. En revanche, les mécanismes par lesquels lespolyphénols activent la production de NO endothélial ne sontpas encore clairement élucidés. De même, les travaux deRamasamy et al. [56] montrent que les antioxydantsphénoliques tels que l’acide caféique induisent la productionde NO endothélial. Toutefois, notre équipe a montré quetoute relaxation est abolie en absence de Ca2+ extracellulaire,suggérant ainsi que les polyphénols peuvent induire un signalcalcique dans la cellule endothéliale et de ce fait moduler di-rectement la production de NO [10]. Les polyphénols sontdes composés capables de moduler l’homéostasie calcique.Ainsi, le resveratrol et la quercetine induisent une élévationde la [Ca2+]i en activant préalablement des canaux K+ ou eninhibant les Ca2+-ATPases du réticulum endoplasmique dansles cellules endothéliales [57,58]. La stimulation des cellulesendothéliales par la delphinidine induit une augmentation àlafois de la teneur cytosolique en calcium ([Ca2+]i) et de laphosphorylation sur tyrosine de protéines intracellulaires,deux acteurs participant à la régulation de l’activité de laeNOS et par conséquent de la production de NO. Cette éléva-tion de [Ca2+]i est tributaire de l’activation de la voie de laPLC, probablement secondaire àcelle d’une protéine G sen-sible à la toxine pertussique, et de l’activation de protéinestyrosine kinases [14].
Il est aussi connu que les polyphénols peuvent moduler lateneur en NO en agissant sur les phosphodiéstérases (PDE)présentes dans les cellules endothéliales : PDE2 et PDE4[59]. L’ inhibition des PDE par les polyphénols et plusparticulièrement par la delphinidine [60,61] augmente leNO dans les cellules endothéliales.
La consommation modérée à plus long terme de poly-phénols modifie également la motricité vasculaire. En effet,le gavage de rats normotendus avec des polyphénols(20 mg/kg, Provinols™) pendant 7 jours a pour effet d’amé-liorer la fonction endothéliale, reflétée par une relaxationplus importante à l’acétylcholine liée àune augmentation dela production de NO endothélial. De manière concomitante,une réduction de la pression artérielle sans induction d’unstress oxydatif est observée [12]. La supplémentation ali-mentaire des rats en polyphénols (40 mg/kg, Provinols™)pendant 3 semaines produit les mêmes effets et s’accompa-gne d’une élévation de l’activité de la eNOS dans leventricule gauche et dans l’aorte de ces animaux [11]. Bienque cela n’ait jamais été mis en évidence dans les tissusd’animaux gavés, les polyphénols modulent non seulementdirectement l’expression de la eNOS mais également sonactivité dans divers types de cellules endothéliales.
Le NO produit par la NO-synthase inductible, responsa-ble ne module la vasomotricité que lors des processus
S. Martin, R. Andriantsitohaina / Annales de cardiologie et d’angéiologie 51 (2002) 304–315 309

d’ inflammation et ne sera pas discuté dans ce manuscrit. Deplus, les polyphénols ont été très largement décrits commedes inhibiteurs de l’activité NO-synthase inductible [62]suggérant une protection des polyphénols contre les effetsdélétères du NO.
3.2.1.2. Polyphénols et facteur contracturant : l’endothé-line (ET-1). L’ET-1 est le facteur contracturant le plus puis-sant produit par l’endothélium. L’ET-1 est un peptidetranscrit dans les cellules endothéliales en réponse à l’hy-poxie, aux forces de cisaillement du flux sanguin ou àl’ ischémie. Une fois liée à son récepteur, situé sur lesmuscles musculaires lisses vasculaires, elle induit uneélévation de la [Ca2+]i, responsable de la contraction duvaisseau. Les effets des polyphénols sur la transduction dusignal engendrépar l’ET-1 n’ont étéque très récemment misen évidence par le groupe de Corder [63]. Ainsi, de trèsfaibles quantités de polyphénols de vin rouge suffisent àabolir la synthèse d’ET-1 dans les cellules endothélialesd’aorte de bœuf (BAEC) (IC50 = 5,0 ± 0,4 µl/ml). Aucundes composés polyphénoliques purifiés (resveratrol, caté-chine, épicatéchine, pelargonidine, delphinidine, cyanidine),aux mêmes concentrations, n’exerce d’effet inhibiteur sur lasynthèse d’ET-1, ce qui est en contradiction avec des étudesantérieures montrant que la quercetine (0,5–50 µM) inhibela production d’ET-1 dans les cellules endothéliales de veineombilicale humaine (HUVEC) [64]. À ce jour, les mécanis-mes d’action des polyphénols ne sont pas encore élucidés,bien que l’on puisse supposer qu’ ils agissent sur les voiesimpliquées dans la régulation de la production d’ET-1 tellesque les tyrosine kinases, la PKC ou directement le NO.Aucune donnée n’est actuellement disponible quant auxeffets in vivo des polyphénols sur la production d’ET-1.
En conclusion, les polyphénols sont capables d’agirdirectement sur les deux facteurs endothéliaux majeursrégulant la vasomotricité. Bien que les conséquences in vivod’un traitement chronique soient bien documentées en cequi concerne le NO, celles concernant l’ET-1 restent àétablir. Pourtant, il y a vraisemblablement mise en place demécanismes de compensation puisqu’ il est difficile de croirequ’une élévation de la teneur en NO et inversement qu’unediminution de la production d’ET-1 soient physiologique-ment bénéfiques à long terme. Toutefois, ces propriétés sontun atout dans des situations pathologiques telles que l’hy-pertension artérielle et expliquent les propriétés vasculo-protectrices attribuées à ces composés.
3.2.2. Effets des polyphénols sur la prolifération,la migration et l’apoptose des cellules endothélialesConséquences sur le remodelage vasculaireet l’angiogenèse
La prolifération, la migration et l’apoptose des cellulesendothéliales sont trois processus régulés de manière dras-tique par l’organisme. Dans les conditions physiologiques,l’apoptose et la prolifération endothéliales sont faibles voirquasi inexistantes. Mais il ne faut pas perdre de vue que
l’endothélium, de par sa position, est sans cesse au contactde macromolécules ou de cellules qui peuvent altérer ou aucontraire contribuer à préserver son intégrité, en modulantdes processus comme l’apoptose, la prolifération ou lamigration cellulaire. Cette dernière propriété est couram-ment attribuée aux polyphénols apportés par l’alimentationcourante. Comme nous l’avons vu jusqu’à présent, lespolyphénols renforcent le contrôle de la motricité, parl’endothélium, en agissant directement sur des facteursendothéliaux. Mais ils peuvent également contribuer àpréserver l’ intégrité de l’endothélium afin que ce dernierpuisse remplir ces différentes fonctions. Le maintien del’ intégrité de l’endothélium vasculaire est intimement lié àla régulation de la prolifération, la migration et l’apoptosedes cellules endothéliales.
3.2.2.1. Polyphénols et prolifération des cellules endothé-liales. Les effets des polyphénols ont été très largementdocumentés en ce qui concerne la prolifération des cellulestumorales ou musculaires lisses pour lesquelles ils sontdécrits comme étant inhibiteurs, ce qui leur confère despropriétés anti-tumorales et anti-hyperplasiques [47]. Enrevanche bien que très peu d’études aient été menées sur lescellules endothéliales, ils y ont également étédécrits commeinhibiteurs de la prolifération [65,66]. Ainsi, les travaux deBrakenhielm et al. [67] montrent que le resveratrol(0,5–10 µM) inhibe la prolifération des cellules endothélia-les de capillaires bovins induite par les facteurs de croiss-sance comme le VEGF et le FGF, en bloquant l’activationdes MAP kinase de type ERK1/2. Les travaux de Hsieh etal. [68] ont mis en évidence que le resveratrol (10–100 µM)bloque la prolifération des cellules endothéliales d’artèrespulmonaires bovines (BPAEC) en accumulant les protéinesp53 et p21WAF1/CIP1 ce qui empêche la transition de G1 à Sdu cycle cellulaire. De même, le resveratrol inhibe laprolifération des BAEC [69]. Dans ces deux dernièresétudes, bien qu’une induction de l’expression de la eNOS aitété observée, l’ implication du NO dans les effets inhibiteursdes polyphénols sur la prolifération n’a jamais clairementété déterminée. À ce jour, aucune étude ne permet d’établirun lien entre les effets inhibiteurs des polyphénols sur laprolifération cellulaire et le NO. Toutefois, l’exploration decette voie serait intéressante compte tenu des effets connusdu NO. Le NO peut être tour à tour activateur ou inhibiteurde la prolifération des cellules endothéliales. Un autreaspect susceptible d’expliquer l’ inhibition de la proliféra-tion par les polyphénols serait leurs effets inhibiteurs sur lesphosphodiésterases (PDE). L’élévation du taux intracellu-laire d’adénosine monophosphate cyclique résultant del’ inhibition des PDE empêche la progression de la phase G1à S du cycle cellulaire des HUVEC. L’augmentation de laconcentration d’AMPc induirait l’ inhibition de l’expressiondes cyclines (D1, A ou B) et l’ induction de CKI, principa-lement p27KIP1 [70]. Mais aucune donnée ne permetd’établir un lien entre les effets inhibiteurs des polyphénolssur la prolifération cellulaire et l’ inhibition des PDEs.
310 S. Martin, R. Andriantsitohaina / Annales de cardiologie et d’angéiologie 51 (2002) 304–315

3.2.2.2. Polyphénols et migration des cellules endothé-liales. Les effets des polyphénols sur la migration cellulairesont très largement documentés en ce qui concerne lescellules tumorales et les cellules musculaires lisses où ilssont décrits comme des inhibiteurs [47]. De même, lespolyphénols tels que le resveratrol et la quercetine inhibentla migration des HUVEC et des BAEC, stimulée ou non parle VEGF [65,67]. Les mécanismes par lesquels les polyphé-nols inhibent la migration des cellules endothéliales sonttotalement inconnus à ce jour. Les polyphénols contenusdans le thé vert ont la capacité d’ inhiber l’ invasion del’endothélium par des cellules tumorales d’un fibrosarcomehumain, en bloquant la sécrétion de métallo-protéinases 2 et9 (MMP2 et MMP9) par ces cellules [71]. De même, ladelphinidine et le resveratrol inhibent la sécrétion des MMPdans les cellules HT180 et dans les cellules musculaireslisses respectivement. Les MMP sont responsables du cli-vage des molécules de la matrice interstitielle (collagène,fibronectine, vitronectine ou élastine), ce qui permet larégulation de l’ interaction cellule/matrice, un point clef dela migration cellulaire [72]. Bien que cela n’ait jamais étémontré, les polyphénols pourraient agir de manière similairepour bloquer la migration des cellules endothéliales. Lespolyphénols peuvent aussi agir directement sur des acteursrégulant le cytosquelette. Ainsi, le kaempférol a été décritcomme un inhibiteur de la kinase des chaînes légères de lamyosine de par son entrée en compétition avec l’ATP [73].Cette kinase régule la contraction du cytosquelette enphosphorylant la myosine, ce qui permet l’établissement deponts entre les filaments d’actine et de myosine et ainsi ledéveloppement de forces contractiles conduisant à la migra-tion. Un renforcement des points focaux d’adhésion aégalement été observé dans les BPAEC traitées avec leresveratrol (100 µM). Ce phénomène dépend de l’activitédes tyrosine kinases et d’une modulation de la [Ca2+]i [69].
En conclusion, les polyphénols pourraient bloquer lamigration des cellules endothéliales en empêchant le déta-chement de la cellule de sa matrice et en empêchant ledéveloppement des forces contractiles nécessaires au dépla-cement de la cellule.
3.2.2.3. Polyphénols et apoptose des cellules endothé-liales. L’apoptose des cellules endothéliales réduit forte-ment l’ imperméabilité de l’endothélium, facilitant ainsi ledéveloppement de pathologies diverses. Les effets despolyphénols sur l’apoptose sont très largement documentésen ce qui concerne les cellules tumorales où ils sont décritscomme des inducteurs [47]. En revanche, les extraitspolyphénoliques de vin rouge n’ont aucun effet sur l’apop-tose des cellules musculaires lisses alors qu’ ils inhibent leurprolifération [49]. Inversement, les polyphénols de vinrouge et l’épicatéchine ont été décrits comme protecteurscontre l’apoptose induite par les LDL oxydées ou leperoxyde d’hydrogène, respectivement, dans les BAEC etles fibroblastes [74]. En revanche, le resveratrol induitl’apoptose des HUVEC [66]. Ces effets, opposés, des
polyphénols semblent être liés à leur action sur différentsacteurs régulant l’apoptose. Ainsi, bien que cela n’ait jamaisété mis en évidence dans les cellules endothéliales, lespolyphénols ont la capacité de moduler l’activité des cas-pases. Les polyphénols de thé vert induisent l’apoptose descellules tumorales en activant les caspases-3 et 9. Enrevanche, l’épicatéchine est capable de bloquer l’apoptosedes fibroblastes en bloquant l’activation de la caspase-3. Laquercetine, quant à elle, inhibe directement la pompe àprotons F0F1-ATPase de la mitochondrie. La perturbationrésultante du potentiel mitochondrial provoque la libérationdu cytochrome C, qui une fois lié à la protéine APAF-1,active la caspase-9, un exécuteur final de l’apoptose. Leseffets des polyphénols s’expliquent également par leurcapacité à moduler le taux d’expression des protéinesprotectrices (Bcl-2, Bcl-xL et A1) ou inductrices (Bax, Bidet Bak) de l’apoptose. Ainsi, l’acide tannique et le resvera-trol induisent l’apoptose en augmentant le taux cellulaire deBax, en inhibant sa dégradation par le protéasome, ou eninhibant l’expression de Bcl-2 respectivement dans lescellules tumorales. L’ inhibition de l’apoptose, induite parles LDL oxydées, des BAEC par les polyphénols de vinrouge est, quant à elle, liée à une modulation de l’homéos-tasie calcique, un médiateur universel de l’apoptose. Leseffets protecteurs des polyphénols peuvent également êtreattribués au NO, puisque ce dernier est capable d’ induiredes protéines de stress (HSP-32 ou –70) (inhibant l’ induc-tion des caspases), d’ inhiber les caspases par S-nitrosylationet d’ inhiber le relarguage du cytochrome C de la mitochon-drie. Le NO exerce également ses effets protecteurs par leGMPc, qui peut moduler l’homéostasie Ca2+ et inhiberl’activation des caspases par les protéines kinases G. Enconclusion, les polyphénols semblent avoir un effet doublesur l’apoptose des cellules endothéliales. Dans le premiercas, ils permettent la protection de l’endothélium contredivers stimuli (les LDL oxydées ou les cytokines pro-inflammatoires) ce qui permet de lutter contre le dévelop-pement de pathologies telles que l’athérosclérose. Dans ledeuxième cas, ils entraînent la destruction de l’endothéliumce qui n’a lieu d’être que dans la lutte contre le cancer.
3.2.2.4. Polyphénols, remodelage vasculaire et angio-genèse. Le remodelage vasculaire et l’angiogenèse sontdeux phénomènes impliqués directement dans le dévelop-pement et l’entretien de pathologies telles que le cancer, lediabète, l’arthrite rhumatoide, l’athérosclérose et l’hyper-tension [75]. Ils nécessitent une régulation coordonnée del’apoptose, de la prolifération et de la migration des cellulesendothéliales.
Le remodelage vasculaire représente la capacité de laparoi vasculaire à se réorganiser en fonction des stimuliauxquels il est soumis, notamment le flux sanguin. Dans unmodèle animal in vivo d’angioplastie par ballonnet, lesdommages causés par le ballonnet conduisent à une hyper-plasie de l’ intima, reflet d’une prolifération excessive de lacouche musculaire lisse. À long terme, une réduction du
S. Martin, R. Andriantsitohaina / Annales de cardiologie et d’angéiologie 51 (2002) 304–315 311

diamètre du vaisseau sanguin et une augmentation de larésistance vasculaire peuvent être observées. Il a été montréque l’épaississement du vaisseau est atténuée de près de53 % chez des lapins ayant ingéré du vin rouge (5 ml/kg depoids) [76]. De même, la consommation de quercetine(10 mg/kg) par des rats spontanément hypertendus (SHR)pendant 5 semaines a pour conséquence de réduire l’hyper-trophie cardiaque et rénale alors que le traitement de ratsnormotendus (WKY) reste sans effet [77]. Notre groupe amis en évidence qu’un apport nutritionnel de polyphénolsde vin rouge (ProvinolsTM : 40 mg/kg) pendant 3 semaines,à des rats hypertendus, a pour conséquences de réduire lafibrose cardiaque de près de 74 % et l’épaississement del’aorte associée à l’hypertension [11]. Dans tous les cas, unediminution de la pression sanguine, une amélioration consi-dérable de la relaxation dépendante de l’endothélium et dela fonction vasculaire sont observées.
L’angiogenèse représente la capacité de développementde nouveaux vaisseaux à partir de vaisseaux préexistants.Les polyphénols sont de puissants agents anti-angio-géniques. Fotsis et al. [78] ont été les premiers à montrerque la génistéine inhibe l’angiogenèse en bloquant laprolifération des cellules endothéliales. Depuis, de nom-breuses études abondant dans le même sens, réalisées tant invitro qu’ in vivo, ont été répertoriées. Ainsi, l’administrationorale de resveratrol (48 µg/kg) à des souris pendant 3 joursralentit considérablement l’angiogenèse vasculaire associéeà une réparation tissulaire ou à une revascularisation de lacornée [67]. De même, le resveratrol (6–100 µM) et laquercetine (6–100 µM) inhibent l’angiogenèse in vitro [65].
En conclusion, les polyphénols protègent le systèmevasculaire d’un remodelage anormal et d’une angiogenèsepathologique. Ces propriétés sont intimement liées à leurcapacité àinhiber à la fois la prolifération [68], la migration[65] et à moduler l’apoptose des cellules endothéliales. Il nefaut toutefois pas perdre de vue que s’ ils ont des effets surl’endothélium, ils agissent également de manière similairesur la couche musculaire lisse [49].
4. Conclusion
Ainsi, les polyphénols possèdent des propriétés antioxy-dantes, anti-agrégantes plaquettaires, anti-inflammatoires,anti-allergènes, antitumorales et antithrombotiques. Ils ontégalement la capacité de moduler les processus hémostati-ques, vasomoteurs, apoptotiques, prolifératifs et migratoireset finalement le remodelage vasculaire et l’angiogenèse.
Les polyphénols, de par leurs effets multiples sur le sys-tème cardio-vasculaire, présentent un intérêt à ne pas négli-ger dans la prévention mais également dans le traitement desnombreuses pathologies. Les composés polyphénoliquessont d’ailleurs de plus en plus utilisés en thérapeutique. Plusde 50 spécialités contenant un ou plusieurs polyphénols sontactuellement disponibles en France. Ils sont regroupés dansla catégorie des veinotoniques et des vasculo-protecteurs.Parmi les veinotoniques, nous citerons le Relvenet ou le
Cirkant renfermant du ruténoside, le Daflont ou le Diosmiltrenfermant de la diosmine et le Vasococitrolt ou leCemaflavonet renfermant des citroflavonoïdes. Un certainnombre de molécules polyphénoliques sont également enétude clinique comme anti-agrégant plaquettaire ouhypotenseur sans résultats probants. En revanche, les étudescliniques réalisées avec le Daflont 500 mg montrent uneamélioration significative de l’ insuffisance veineuse [79-82].Les études réalisées par Fuhrman et al. [83] montrent qu’uneconsommation quotidienne de 400 ml de vin rouge réduitconsidérablement l’oxydation des LDL. Une augmentationde la capacité antioxydante du plasma est également obser-vée lors d’une supplémentation en extraits polyphénoliquesde vin rouge [84-86]. Une étude clinique a permis de montrerune activité anticancéreuse de la quercetine administrée parvoie intraveineuse chez des patients atteints de cancer [87].Le resveratrol est actuellement en phase I d’étude clinique(pharmacocinétique) à l’ institut de virologie humaine deBaltimore (Institute of Human Virology, Maryland) pour sonutilisation dans le traitement du sida (Source : Institute ofHuman Virology). Il est également en phase d’études pré-clinique et clinique en Caroline du Sud (Medical Universityof South Carolina - Caroline du Sud) pour son utilisationdans le traitement de divers cancers (Sources : The NationalCancer Institute). Cet intérêt pour le resevratrol fait suite auxtravaux réalisés in vitro et in vivo sur l’animal montrant unintérêt potentiel de ce composé pour le traitement du sida etde divers cancers.
Ainsi, les études cliniques réalisées chez l’Homme révè-lent un intérêt thérapeutique évident des polyphénols. Tou-tefois, de nombreuses études fondamentales sont encorenécessaires pour comprendre les mécanismes d’action et leseffets au niveau cellulaire et intégré des polyphénols.
Remerciements
Les auteurs tiennent à remercier Dr M.C. Martinez pourles discussions scientifiques et la lecture constructive dumanuscrit.
Références
[1] St Leger AS, Cochrane AL, Moore F. Factors associated with cardiacmortality in developed countries with particular reference to theconsumption of wine. Lancet 1979;1:1017–20.
[2] Tjonneland A, Gronbaek M, Stripp C, Overvad K. Wine intake anddiet in a random sample of 48763 Danish men and women. Am JClin Nutr 1999;69:49–54.
[3] Renaud SC, Gueguen R, Schenker J, d’Houtaud A. Alcohol andmortality in middle-aged men from eastern France. Epidemiology1998;9:184–8.
[4] Masquelier J. Wine in nutrition. Aliment Vie 1965;53:261–70.
[5] Van Tol A, van der Gaag MS, Scheek LM, van Gent T, Hendriks HF.Changes in postprandial lipoproteins of low and high density causedby moderate alcohol consumption with dinner. Atherosclerosis1998;141(Suppl 1):S101–3.
312 S. Martin, R. Andriantsitohaina / Annales de cardiologie et d’angéiologie 51 (2002) 304–315

[6] Zoucas E, Bergqvist D, Goransson G, Bengmark S. Effect of acuteethanol intoxication on primary haemostasis, coagulation factors andfibrinolytic activity. Eur Surg Res 1982;14:33–44.
[7] Bravo L. Polyphenols: chemistry, dietary sources, metabolism, andnutritional significance. Nutr Rev 1998;56:317–33.
[8] Andriambeloson E, Kleschyov AL, Muller B, Beretz A, Stoclet JC,Andriantsitohaina R. Nitric oxide production and endothelium-dependent vasorelaxation induced by wine polyphenols in rat aorta.Br J Pharmacol 1997;120:1053–8.
[9] Andriambeloson E, Magnier C, Haan-Archipoff G, Lobstein A,Anton R, Beretz A, et al. Natural dietary polyphenolic compoundscause endothelium-dependent vasorelaxation in rat thoracic aorta. JNutr 1998;128:2324–33.
[10] Andriambeloson E, Stoclet JC, Andriantsitohaina R. Mechanism ofendothelial nitric oxide-dependent vasorelaxation induced by winepolyphenols in rat thoracic aorta. J Cardiovasc Pharmacol1999;33:248–54.
[11] Bernatova I, Pechanova O, Babal P, Kysela S, Stvrtina S, Andriant-sitohaina R. Wine polyphenols improve cardiovascular remodelingand vascular function in NO-deficient hypertension. Am J PhysiolHeart Circ Physiol 2002;282:H942–8.
[12] Diebolt M, Bucher B, Andriantsitohaina R. Wine polyphenolsdecrease blood pressure, improve NO vasodilatation, and inducegene expression. Hypertension 2001;38:159–65.
[13] Martin S, Diebolt M, Andriantsitohaina R. Moderate alcoholconsumption and cardiovascular diseases. Pathol Biol (Paris)2001;49:769–74.
[14] Martin S, Andriambeloson E, Takeda K, Andriantsitohaina R. Redwine polyphenols increase calcium in bovine aortic endothelialcells: a basis to elucidate signalling pathways leading to nitric oxideproduction. Br J Pharmacol 2002;135:1579–87.
[15] Furchgott RF, Zawadzki JV. The obligatory role of endothelial cellsin the relaxation of arterial smooth muscle by acetylcholine. Nature1980;288:373–6.
[16] Cines DB, Pollak ES, Buck CA, Loscalzo J, Zimmerman GA,McEver RP, et al. Endothelial cells in physiology and in thepathophysiology of vascular disorders. Blood 1998;91:3527–61.
[17] Brown AA, Hu FB. Dietary modulation of endothelial function:implications for cardiovascular disease. Am J Clin Nutr2001;73:673–86.
[18] Boulanger CM, Vanhoutte PM. Facteurs vasoactifs produits parl’endothélium. In: Flammarion, editor. Biologie et pathologie ducœur et des vaisseaux. Paris: GRRC; 2002. p. 447–53.
[19] Akimoto S, Mitsumata M, Sasaguri T, Yoshida Y. Laminar shearstress inhibits vascular endothelial cell proliferation by inducingcyclin-dependent kinase inhibitor p21(Sdi1/Cip1/Waf1. Circ Res2000;86:185–90.
[20] Dimmeler S, Haendeler J, Galle J, Zeiher AM. Oxidized low-densitylipoprotein induces apoptosis of human endothelial cells by activa-tion of CPP32-like proteases. A mechanistic clue to the ’ response toinjury’ hypothesis. Circulation 1997;95:1760–3.
[21] Ziche M, Morbidelli L, Masini E, Amerini S, Granger HJ,Maggi CA, et al. Nitric oxide mediates angiogenesis in vivo andendothelial cell growth and migration in vitro promoted by subs-tance P. J Clin Invest 1994;94:2036–44.
[22] Cai H, Harrison DG. Endothelial dysfunction in cardiovasculardiseases: the role of oxidant stress. Circ Res 2000;87:840–4.
[23] Ross R. Atherosclerosis – an inflammatory disease. N Engl J Med1999;340:115–26.
[24] Lusis AJ. Atherosclerosis. Nature 2000;407:233–41.[25] Rudic RD, Shesely EG, Maeda N, Smithies O, Segal SS, Sessa WC.
Direct evidence for the importance of endothelium-derived nitricoxide in vascular remodeling. J Clin Invest 1998;101:731–6.
[26] Richard V, Thulliez C. Dysfonction endothéliale. In: Flammarion,editor. Biologie et pathologie du cœur et des vaisseaux. Paris:GRRC; 2002. p. 381–90.
[27] Bode-Boger SM, Boger RH, Alfke H, Heinzel D, Tsikas D,Creutzig A, et al. L-arginine induces nitric oxide-dependent vasodi-lation in patients with critical limb ischemia. A randomized, control-led study. Circulation 1996;93:85–90.
[28] Hamon M, Vallet B, Bauters C, Wernert N, McFadden EP, Lablan-che JM, et al. Long-term oral administration of L-arginine reducesintimal thickening and enhances neoendothelium-dependentacetylcholine-induced relaxation after arterial injury. Circulation1994;90:1357–62.
[29] Marks DS, Vita JA, Folts JD, Keaney Jr JF, Welch GN, Los-calzo J. Inhibition of neointimal proliferation in rabbits aftervascular injury by a single treatment with a protein adduct of nitricoxide. J Clin Invest 1995;96:2630–8.
[30] Pepke-Zaba J, Higenbottam TW, Dinh-Xuan AT, Stone D, Wal-lwork J. Inhaled nitric oxide as a cause of selective pulmonaryvasodilatation in pulmonary hypertension. Lancet 1991;338:1173–4.
[31] Chaux A, Ruan XM, Fishbein MC, Ouyang Y, Kaul S, Pass JA, et al.Perivascular delivery of a nitric oxide donor inhibits neointimalhyperplasia in vein grafts implanted in the arterial circulation. JThorac Cardiovasc Surg 1998;115:604–12.
[32] Chen C, Hanson SR, Keefer LK, Saavedra JE, Davies KM,Hutsell TC, et al. Boundary layer infusion of nitric oxide reducesearly smooth muscle cell proliferation in the endarterectomizedcanine artery. J Surg Res 1997;67:26–32.
[33] Hanson SR, Hutsell TC, Keefer LK, Mooradian DL, Smith DJ. Ni-tric oxide donors: a continuing opportunity in drug design. AdvPharmacol 1995;34:383–98.
[34] Pulfer SK, Ott D, Smith DJ. Incorporation of nitric oxide-releasingcrosslinked polyethyleneimine microspheres into vascular grafts. JBiomed Mater Res 1997;37:182–9.
[35] Smith DJ, Chakravarthy D, Pulfer S, Simmons ML, Hrabie JA,Citro ML, et al. Nitric oxide-releasing polymers containing the[N(O)NO]- group. J Med Chem 1996;39:1148–56.
[36] Stamler JS, Simon DI, Osborne JA, Mullins ME, Jaraki O, Mi-chel T, et al. Snitrosylation of proteins with nitric oxide: synthesisand characterization of biologically active compounds. Proc NatlAcad Sci U S A 1992;89:444–8.
[37] von der Leyen HE, Gibbons GH, Morishita R, Lewis NP, Zhang L,Nakajima M, et al. Gene therapy inhibiting neointimal vascularlesion: in vivo transfer of endothelial cell nitric oxide synthase gene.Proc Natl Acad Sci U S A 1995;92:1137–41.
[38] Mazza G. Anthocyanins in grapes and grape products. Crit Rev FoodSci Nutr 1995;35:341–71.
[39] Harborne JB. The anthocyanin pigments. Comparative Biochemistryof the flavonoides. New York: Academic press; 1967 p. 1–30.
[40] Kuhnau J. The flavonoids. A class of semi-essential food compo-nents: their role in human nutrition. World Rev Nutr Diet1976;24:117–91.
[41] Pierpoint WS. Flavonoids in the human diet. Prog Clin Biol Res1986;213:125–40.
[42] Hertog MG, Feskens EJ, Hollman PC, Katan MB, Kromhout D.Dietary antioxidant flavonoids and risk of coronary heart disease:the Zutphen Elderly Study. Lancet 1993;342:1007–11.
[43] Hertog MG, Hollman PC, Katan MB, Kromhout D. Intake ofpotentially anticarcinogenic flavonoids and their determinants inadults in The Netherlands. Nutr Cancer 1993;20:21–9.
[44] Justesen U, Knuthsen P, Leth T. Determination of plant polyphenolsin Danish foodstuffs by HPLC-UV and LC-MS detection. CancerLett 1997;114:165–7.
[45] Paganga G, Rice-Evans CA. The identification of flavonoids asglycosides in human plasma. FEBS Lett 1997;401:78–82.
[46] Matsumoto H, Inaba H, Kishi M, Tominaga S, Hirayama M,Tsuda T. Orally administered delphinidin 3-rutinoside and cyanidin3-rutinoside are directly absorbed in rats and humans and appear inthe blood as the intact forms. J Agric Food Chem 2001;49:1546–51.
S. Martin, R. Andriantsitohaina / Annales de cardiologie et d’angéiologie 51 (2002) 304–315 313

[47] Middleton Jr E, Kandaswami C, Theoharides TC. The effects ofplant flavonoids on mammalian cells: implications for inflammation,heart disease, and cancer. Pharmacol Rev 2000;52:673–751.
[48] Osakabe N, Yasuda A, Natsume M, Takizawa T, Terao J, Kondo K.Catechins and their oligomers linked by C4 --> C8 bonds are majorcacao polyphenols and protect low-density lipoprotein from oxida-tion in vitro. Exp Biol Med (Maywood) 2002;227:51–6.
[49] Iijima K, Yoshizumi M, Hashimoto M, Kim S, Eto M, Ako J, et al.Red wine polyphenols inhibit proliferation of vascular smoothmuscle cells and downregulate expression of cyclin A gene. Circu-lation 2000;101:805–11.
[50] Wollny T, Aiello L, Di Tommaso D, Bellavia V, Rotilio D,Donati MB, et al. Modulation of haemostatic function and preven-tion of experimental thrombosis by red wine in rats: a role forincreased nitric oxide production. Br J Pharmacol 1999;127:747–55.
[51] Flesch M, Schwarz A, Bohm M. Effects of red and white wine onendothelium-dependent vasorelaxation of rat aorta and human coro-nary arteries. Am J Physiol 1998;275:H1183–90.
[52] Burns J, Gardner PT, O’Neil J, Crawford S, Morecroft I, Mc-Phail DB, et al. Relationship among antioxidant activity, vasodila-tion capacity, and phenolic content of red wines. J Agric Food Chem2000;48:220–30.
[53] Jimenez R, Andriambeloson E, Duarte J, Andriantsitohaina R,Jimenez J, Perez-Vizcaino F, et al. Involvement of thromboxane A2in the endothelium-dependent contractions induced by myricetin inrat isolated aorta. Br J Pharmacol 1999;127:1539–44.
[54] Fitzpatrick DF, Hirschfield SL, Coffey RG. Endothelium-dependentvasorelaxing activity of wine and other grape products. Am J Physiol1993;265:H774–8.
[55] Matsuo S, Nakamura Y, Takahashi M, Ouchi Y, Hosoda K, No-zawa M. Effect of red wine and ethanol on production of nitric oxidein healthy subjects. Am J Cardiol 2001;87:1029–31.
[56] Ramasamy S, Drummond GR, Ahn J, Storek M, Pohl J, Parthasa-rathy S. Modulation of expression of endothelial nitric oxidesynthase by nordihydroguaiaretic acid, a phenolic antioxidant incultured endothelial cells. Mol Pharmacol 1999;56:116–23.
[57] Li HF, Chen SA, Wu SN. Evidence for the stimulatory effect ofresveratrol on Ca2+-activated K+ current in vascular endothelialcells. Cardiovasc Res 2000;45:1035–45.
[58] McKenna E, Smith JS, Coll KE, Mazack EK, Mayer EJ, Antana-vage J, et al. Dissociation of phospholamban regulation of cardiacsarcoplasmic reticulum Ca2+ ATPase by quercetin. J Biol Chem1996;271:24517–25.
[59] Lugnier C, Schini VB. Characterization of cyclic nucleotide phos-phodiesterases from cultured bovine aortic endothelial cells. Bio-chem Pharmacol 1990;39:75–84.
[60] Beretz A, Anton R, Cazenave JP. The effects of flavonoids on cyclicnucleotide phosphodiesterases. Prog Clin Biol Res1986;213:281–96.
[61] Beretz A, Briancon-Scheid F, Stierle A, Corre G, Anton R, Caze-nave JP. Inhibition of human platelet cyclic AMP phosphodiesteraseand of platelet aggregation by a hemisynthetic flavonoid, amento-flavone hexaacetate. Biochem Pharmacol 1986;35:257–62.
[62] Paquay JB, Haenen GR, Stender G, Wiseman SA, Tijburg LB,Bast A. Protection against nitric oxide toxicity by tea. J Agric FoodChem 2000;48:5768–72.
[63] Corder R, Douthwaite JA, Lees DM, Khan NQ, Viseu Dos San-tos AC, Wood EG. Endothelin-1 synthesis reduced by red wine.Nature 2001;414:863–4.
[64] Zhao X, Gu Z, Attele AS, Yuan CS. Effects of quercetin on therelease of endothelin, prostacyclin and tissue plasminogen activatorfrom human endothelial cells in culture. J Ethnopharmacol1999;67:279–85.
[65] Igura K, Ohta T, Kuroda Y, Kaji K. Resveratrol and quercetin inhibitangiogenesis in vitro. Cancer Lett 2001;171:11–6.
[66] Szende B, Tyihak E, Kiraly-Veghely Z. Dose-dependent effect ofresveratrol on proliferation and apoptosis in endothelial and tumorcell cultures. Exp Mol Med 2000;32:88–92.
[67] Brakenhielm E, Cao R, Cao Y. Suppression of angiogenesis, tumorgrowth, and wound healing by resveratrol, a natural compound inred wine and grapes. FASEB J 2001;15:1798–800.
[68] Hsieh TC, Juan G, Darzynkiewicz Z, Wu JM. Resveratrol increasesnitric oxide synthase, induces accumulation of p53 andp21(WAF1/CIP1), and suppresses cultured bovine pulmonary arteryendothelial cell proliferation by perturbing progression through Sand G2. Cancer Res 1999;59:2596–601.
[69] Bruder JL, Hsieh TT, Lerea KM, Olson SC, Wu JM. Inducedcytoskeletal changes in bovine pulmonary artery endothelial cells byresveratrol and the accompanying modified responses to arterialshear stress. BMC Cell Biol 2: 1 2001;2:1.
[70] Koyama H, Bornfeldt KE, Fukumoto S, Nishizawa Y. Molecularpathways of cyclic nucleotide-induced inhibition of arterial smoothmuscle cell proliferation. J Cell Physiol 2001;186:1–10.
[71] Maeda-Yamamoto M, Kawahara H, Tahara N, Tsuji K, Hara Y,Isemura M. Effects of tea polyphenols on the invasion and matrixmetalloproteinases activities of human fibrosarcoma HT1080 cells. JAgric Food Chem 1999;47:2350–4.
[72] Leppert D, Lindberg RL, Kappos L, Leib SL. Matrix metalloprotei-nases: multifunctional effectors of inflammation in multiple sclerosisand bacterial meningitis. Brain Res Brain Res Rev 2001;36:249–57.
[73] Rogers JC, Williams Jr DL. Kaempferol inhibits myosin light chainkinase. Biochem Biophys Res Commun 1989;164:419–25.
[74] Vieira O, Escargueil-Blanc I, Meilhac O, Basile JP, Laranjinha J,Almeida L, et al. Effect of dietary phenolic compounds on apoptosisof human cultured endothelial cells induced by oxidized LDL. Br JPharmacol 1998;123:565–73.
[75] Carmeliet P, Jain RK. Angiogenesis in cancer and other diseases.Nature 2000;407:249–57.
[76] Feng AN, Chen YL, Chen YT, Ding YZ, Lin SJ. Red wine inhibitsmonocyte chemotactic protein-1 expression and modestly reducesneointimal hyperplasia after balloon injury in cholesterol-Fed rab-bits. Circulation 1999;100:2249–54.
[77] Duarte J, Perez-Palencia R, Vargas F, Ocete MA, Perez-Vizcaino F,Zarzuelo A, et al. Antihypertensive effects of the flavonoid quercetinin spontaneously hypertensive rats. Br J Pharmacol2001;133:117–24.
[78] Fotsis T, Pepper M, Adlercreutz H, Fleischmann G, Hase T,Montesano R, et al. Genistein, a dietary-derived inhibitor of in vitroangiogenesis. Proc Natl Acad Sci U S A 1993;90:2690–4.
[79] Guilhou JJ, Dereure O, Marzin L, Ouvry P, Zuccarelli F, De-bure C, et al. Efficacy of Daflon 500mg in venous leg ulcer healing:a double-blind, randomized, controlled versus placebo trial in 107patients. Angiology 1997;48:77–85.
[80] Guilhou JJ, Fevrier F, Debure C, Dubeaux D, Gillet-Terver MN,Guillot B, et al. Benefit of a 2-month treatment with a micronized,purified flavonoidic fraction on venous ulcer healing. A randomized,double-blind, controlled versus placebo trial. Int J Microcirc ClinExp 1997;17(Suppl 1):21–6.
[81] Menyhelyi G, Acsady G, Hetenyi A, Dubeaux D, Rado G. Thechronobiology and clinical efficacy of Daflon in the treatment ofchronic venous insufficiency. Ter Arkh 1997;69:46–8.
[82] Pecking AP, Fevrier B, Wargon C, Pillion G. Efficacy of Daflon500mg in the treatment of lymphedema (secondary to conventionaltherapy of breast cancer). Angiology 1997;48:93–8.
[83] Fuhrman B, Lavy A, Aviram M. Consumption of red wine withmeals reduces the susceptibility of human plasma and low-densitylipoprotein to lipid peroxidation. Am J Clin Nutr 1995;61:549–54.
314 S. Martin, R. Andriantsitohaina / Annales de cardiologie et d’angéiologie 51 (2002) 304–315

[84] Carbonneau MA, Leger CL, Monnier L, Bonnet C, Michel F,Fouret G, et al. Supplementation with wine phenolic compoundsincreases the antioxidant capacity of plasma and vitamin E oflow-density lipoprotein without changing the lipoprotein Cu2+-oxidizability: possible explanation by phenolic location. Eur J ClinNutr 1997;51:682–90.
[85] Day AP, Kemp HJ, Bolton C, Hartog M, Stansbie D. Effect ofconcentrated red grape juice consumption on serum antioxidant
capacity and low-density lipoprotein oxidation. Ann Nutr Metab1997;41:353–7.
[86] Rice-Evans C, Miller N. Measurement of the antioxidant status ofdietary constituents, low density lipoproteins and plasma. Prosta-glandins Leukot Essent Fatty Acids 1997;57:499–505.
[87] Ferry DR, Smith A, Malkhandi J, Fyfe DW, deTakats PG, Ander-son D, et al. Phase I clinical trial of the flavonoid quercetin:pharmacokinetics and evidence for in vivo tyrosine kinase inhibi-tion. Clin Cancer Res 1996;2:659–68.
S. Martin, R. Andriantsitohaina / Annales de cardiologie et d’angéiologie 51 (2002) 304–315 315