Embed Size (px)
Citation preview

Membrane lipidomics for the discoveryof new antiparasitic drug targetsEric Marechal1*, Mickae l Riou2*, Dominique Kerboeuf3*, Frederic Beugnet4*,Pierre Chaminade5* and Philippe M. Loiseau6*
1 Laboratoire de Physiologie Cellulaire Ve ge tale, UMR 5168, CNRS-CEA-INRA-Universite Joseph Fourier, Institut de Recherches en
Sciences et Technologies pour le Vivant, CEA Grenoble, 17 rue des Martyrs, Grenoble, France2 INRA, UE 1277 Plateforme d’Infectiologie Expe rimentale (PFIE), 37380 Nouzilly, France3 INRA, UR 1282 d’Infectiologie Animale et Sante Publique (IASP), 37380 Nouzilly, France4 Me rial, 29 avenue Tony Garnier, 69007, Lyon, France5 Universite Paris-Sud, Groupe de Chimie Analytique de Paris Sud, EA4041, Faculte de Pharmacie, 5 rue J-B Cle ment, 92296
Cha tenay-Malabry, France6 Universite Paris-Sud, Groupe Chimiothe rapie Antiparasitaire, UMR 8076 CNRS, Faculte de Pharmacie, 5 rue Jean-Baptiste
Cle ment, 92296 Cha tenay-Malabry, France
Advances in lipid separation methods and mass spec-trometry technologies allow the fine characterization ofthe lipidome of parasites, ranging from unicellular pro-tists to worms, which cause threatening infections invertebrates, including humans. Specific lipid structuresor lipid metabolic pathways can inspire the developmentof novel antiparasitic drugs. Changes in the lipid balancein membranes of parasites can also provide clues on thedynamics of drugs and some mechanisms of drug resis-tance. This review highlights recent trends in parasitelipidomics, combined with functional analyses, for thediscovery of novel targets and the development of noveldrugs.
Why target the membrane lipidome of parasites andhow?The life cycles of parasites imply numerous and rapidcellular divisions to invasively proliferate in their hostsand compensate losses when going from one host to anoth-er. In infected vertebrates, the populations of Kinetoplas-tida or Apicomplexa unicellular parasites are nearlyinnumerable. A Leishmania population can count morethan 100 millions of parasites in a patient’s spleen andcirculating blood [1], and Plasmodium falciparum canreach population levels of hundreds of millions [2] tohundreds of trillions [3] in human patients with normalhematocrit. In the case of parasitic helminths, such asTrematoda circulating in blood vessels and colonizingthe liver, pancreas or lung, female worms can producefrom 300 eggs/day for Schistosoma mansoni [4] to thou-sands for Schistosoma japonicum. Intestinal Nematodasuch as roundworms (Ascaris, Haemonchus), whipworms(Trichuris) and hookworms (Necator) produce 3000 to200 000 eggs/day and per female [5].
One issue raised by this dynamic proliferation of para-sites is the very rapid propagation of mutants resisting adrug. Therefore, resistance triggered by chemotherapy
Review
Glossary
Atmospheric pressure chemical ionization (APCI): technique used in mass
spectrometry (MS) to produce ions at atmospheric pressure. APCI is adapted to
the ionization of non-polar (e.g. triacylglycerol) to very polar molecules, with a
molecular weight ranging from around 50 to 1000 kDa.
Atmospheric pressure photo ionization (APPI): technique used in MS to
produce ions after exposure of analytes to UV light photons. APPI is adapted to
the ionization of non-polar (e.g. triacylglycerol) to polar molecules, with a
molecular weight ranging from around 100 to 2000 kDa.
Biomimetic membrane: in vitro reconstitution of a biological membrane, based
on the self-organization of polar lipids in mono- or bilayers.
Electrospray ionization (ESI) and nanoESI: technique used in MS to produce
ions, generating ions in a gas phase without fragmentation of the sprayed
sample. ESI is adapted to the ionization of polar to very polar molecules, with a
molecular weight ranging from around 100 to 100 000 kDa. ESI is currently the
most widely used ionization technique in lipidomics. Disadvantages include
the cost and a phenomenon known as ion suppression in the case of low-
abundance species in complex mixtures. Absolute quantification of lipids
requires class and mass independent internal standards. Miniaturized ESI,
known as nanoelectrospray or nanoESI, allows the automated analysis of very
large samples sets.
Gas chromatography (GC) and GC-MS: method separating and analyzing
compounds that can be vaporized without decomposition, based on their
selective retention in the matrix of specific columns. GC coupled to flame
ionization detection (GC-FID) allows the detection of micrograms to lower
nanograms of vaporizable lipids. GC is very widely used for the determination
of fatty acid profiles in glycerolipids. GC can be coupled to a mass spectro-
meter (GC-MS).
Glycolipid: a glycerolipid or a phospholipid, with a polar head containing a
sugar.
High-performance TLC (HPTLC): automated thin layer chromatography (TLC)
system allowing the parallel analysis of a large number of samples, with higher
throughput and more sensitive detection of lipids. Most recent HPTLC lines
include devices that automatically couple TLC with MS (TLC-MS).
KEGG: database of metabolic pathways developed by the Kyoto Encyclopedia
of Genes and Genomes consortium.
Langmuir monolayers: type of biomimetic membrane made by the self-
assembly of lipids in a monolayer at the air–water interface.
Lipid extraction: procedure by which lipids are extracted from a mixture, based
on their specific solubility in organic solvents. Usually after addition of
solvents, a biphasic organic–water system is obtained and lipids are purified in
the organic phase.
Lipid profile: proportions of each lipid class detected and quantified in a
sample. A lipid profile is usually shown as a histogram.
Lipid raft: membrane microdomains characterized by a specific combination of
glycosphingolipids, sterols and protein receptors, more ordered and tightly
packed than the surrounding bilayer and involved in important biological
functions. In the plasma membrane of parasites, lipid rafts are believed to be
important for pathogenic processes.
Liquid chromatography (LC) and LC-MS: method separating and analyzing
compounds in a liquid solution, based on their selective retention in the matrixCorresponding author: Marechal, E. ([email protected]).* CAP: Consortium anti-Parasitaire/Chemotherapy against Parasites.
496 1471-4922/$ – see front matter � 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.pt.2011.07.002 Trends in Parasitology, November 2011, Vol. 27, No. 11

Review Trends in Parasitology November 2011, Vol. 27, No. 11
of specific columns. High-pressure liquid chromatography (HPLC) allows the
separation of lipids solubilized in organic solvents and can be coupled to a
mass spectrometer (LC-MS).
Mass spectrometry (MS): analytical technique that measures the mass-to-
charge ratio of charged (ionized) molecules. A mass spectrometer is composed
of an ion source, a mass analyzer and an ion detector. The mass analyzer can
be a quadrupole (Q), a time-of-flight analyzer (TOF) or an ion trap. TOF has a
very high mass accuracy. MS data are eventually represented by a spectrum
composed of peaks corresponding to mass-to-charge ratios (m/z). MS peaks
are specific for fragments of ionized lipid molecules. MS sensitivity allows the
detection of picograms to fentograms of lipids.
Matrix-assisted laser desorption/ionization (MALDI) and MALDI-TOF: versatile
technique used in MS to produce ions, allowing the analysis of biomolecules
which tend to be fragile and fragment when ionized by other ionization
methods. MALDI is often used to ionize biomolecules upstream a time-of-flight
analyzer (MALDI-TOF).
Mass-to-charge ratio (m/z): charged particles move in electric and magnetic
fields following a path which is strictly dependent on their mass-to-charge ratio.
MS peaks therefore correspond to the m/z quantities of the charged fragments
hitting the detector. Neutral fragments cannot be detected. When the fragment is
positively charged, this quantity is positive and vice versa. When containing only
one charge, the m/z quantity corresponds to the mass of the fragment.
MetaCyc: database of metabolic pathways developed by the BioCyc con-
sortium.
Neutral loss scan: a tandem MS mode used when a molecular ion, produced
after fragmentation of a lipid class, loses a specific uncharged fragment.
Because uncharged fragments cannot be detected, the specific loss of the
corresponding neutral fragment is recorded. The lipid class can thus be
characterized in a complex mixture by selecting only the initial molecular ions
that lose this specific neutral fragment. Phosphatidylserine species can, for
instance, be detected by constant neutral loss scans of m/z 87.
Phospholipid: a glycerolipid or a sphingolipid, which has a polar head
containing a phosphate.
Polar head group: hydrophilic moiety of polar lipids.
Polar lipid: general term defining lipids having a hydrophobic moiety and a
hydrophilic polar head. Polar lipids make up the bulk of biological membranes.
Precursor ion scan: a tandem MS mode used when a molecular ion, produced
after fragmentation of a lipid class, loses a specific charged fragment, which is
then easily detected. The lipid class can thus be characterized in a complex
mixture by selecting only the initial molecular ions that give this specific
charged fragment. In negative ion mode, the dissociation of lipids induced in
the collision chamber of a mass spectrometer can thus allow the detection of
phosphatidic acid, phosphatidylethanolamine, phosphatidylinositol, phospha-
tidylglycerol and phosphatidylcholine by precursor ion scans of m/z –153, –196,
–241, –153 and +184.
Quadrupole mass analyzer (QMS) and triple quadrupole (QQQ): a QMS is a
part of a mass spectrometer, consisting of four parallel circular rods, filtering
sample ions based on their mass-to-charge ratio (m/z). Ions are thus separated
based on the stability of their trajectories in the oscillating electric fields
applied to the rods. Owing to the linear trajectory of ions, a QMS can be
aligned in series of quadrupoles in so-called triple quadrupoles (QQQ). In a
QQQ, the first QMS allows an initial analysis of ions (based on their m/z ratio)
and a selection of ions of interest for subsequent fragmentation and refine
analysis. The second quadrupole is then used as a collision chamber
generating fragments of the ions filtered in the first QMS. The final refined
MS analysis is eventually achieved in the third QMS. QQQ are tandem MS
instruments that are very useful for the analysis of complex lipid mixtures.
Raft-like structures (RLS): membrane microdomains in which structure and
composition is similar to that of lipid rafts.
Tandem MS: also known as MS/MS, tandem MS involves multiple steps of MS
selection. Between each step, a fragmentation and selection of molecular ions
occurs. Tandem MS can be carried out ‘in space’ when the two MS steps are
physically separated in specific instruments (triple quadrupoles or QQQ, or a
quadrupole and a time-of-flight analyzer in QTOF instruments). Tandem MS
can be carried out ‘in time’ in ion trap analyzers. Different modes of analyses
are classically achieved with tandem MS, including the precursor ion scan and
neutral loss scan modes.
Thin layer chromatography (TLC): method of separating and analyzing
molecules such as lipids on a sheet of glass, plastic or aluminum foil, coated
with a thin layer of absorbent material, usually silica gel. Typically inexpensive
and very well established, this technique has a low resolution in direct
semiquantification using lipid specific dyes; however, it allows the detection of
low micrograms of lipids when coupled to GC, and in most recent techniques,
it is efficient in the picogram range, when automatically coupled to MS.
Time-of-flight (TOF) and quadrupole time-of-flight (QTOF) mass analyzers: a
TOF is part of a mass spectrometer, in which the mass-to-charge ratio (m/z) of
ions is determined after their acceleration by an electric field of known strength
and record of the time it takes the ion to reach a detector. TOF MS has a very
high mass accuracy. A QTOF is a tandem MS instrument in which an initial
quadrupole filters sample ions based on their mass-to-charge ratio (m/z). After
fragmentation, the filtered ions are then separated in a TOF analyzer.
spreads rapidly, resulting in a seemingly never-endingchallenge for medical and veterinary research. Efforts todevelop novel drugs must consequently be sustained.
Additionally, membrane expansion for cell division anddifferentiation, for the elaboration of extracellular barriers,requires a huge resource of lipids. Impairment of lipidbiosynthesis thus appears as a potential strategy of choiceto fight against parasites. Is the membrane lipidome thentargetable for therapeutic purposes? In its simplest defini-tion, a drug target is a component of the parasite, of whichimpairment is lethal or prevents the ability of a pathogen toproceed in its infectious cycle. Basically, one searches for avital enzyme, metabolic pathway or dynamic process, absentfrom the host or sufficiently different, so that developeddrugs have little to no adverse effects [6].
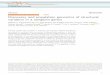
The membrane lipidome is constituted of three majorcategories of lipids, that is glycerolipids, sphingolipids andsterols (Figure 1). To make the bulk of a membrane in aeukaryotic cell, few sterol structures and no more than eightclasses of glycerolipids are required, mainly phosphatidyl-choline (PC), phosphatidylethanolamine(PE), phosphatidyl-serine (PS), phosphatidylinositol (PI), phosphatidylglycerol(PG) and mitochondrial diphosphatidylglycerol (DPG). Dif-ferential phosphorylation of the inositol polar head (seeGlossary) generates seven distinct phosphoinositides, whichare quantitatively minor, but essential determinants of theidentity of membrane compartments of the endomembranesystem. In addition, some membranes, and particularly theplasma membrane, contain sphingolipids and sterol-richdomains, known as rafts, and high proportions of glycolipidswith polar heads harboring species-specific glycosylatedmoieties.
To identify specific targets, a first objective of the lipi-dome mining is thus to identify novel lipid structures anddemonstrate that blocking their synthesis provokes severeeffects. Search of unique glycolipids has thus been veryprolific. Ubiquitous lipids, such as PC, would naively ap-pear as unlikely targets; nevertheless, PC biosynthesis isprecisely one of the current promising targets for a novelseries of antimalarials [7]. A second objective of membranelipidome mining is to link lipid profiles with metabolicpathways, because identical lipid structures can be syn-thesized by distinct pathways. Eventually, lipids are notrestricted to membranes; they can serve to store carbon orenergy, such as triacylglycerol, or act as signal molecules,such as diacylglycerol, phosphatidic acid or oxygenatedfatty acids. Lipidome mining should therefore link mem-brane lipid profiles with any non-lipid metabolic or signaltransduction processes that would be indirectly related tothe membrane lipidome and be essential for the life cycle ofa parasite.
Once a biosynthetic pathway has been validated as anantiparasitic target, the next question is whether it issusceptible to any drugs. Some enzymes involved in syn-thesizing or modifying lipids are soluble proteins, but mostare membrane bound. These enzymes can be surroundedby biological membranes which the drug would need to beable to cross. For instance, the fatty acid synthase of type II(FASII), discussed below in the case of Apicomplexa, islocated inside an organelle known as the apicoplast, in thecytosol of the parasites. A drug candidate targeting a
497

OH OH
OH OH
RONH
β-galactose
OH O
RONH
CH2OH
HO
HH
P
C
CH2CH2 CH2-
H
CH2OOCR 1
CH2OH
CH2OOCR 1 CH2OOCR 1
R2COOR2COO R2COO C H
P
C
- -- -
H
P
NHO
O
OH OH
+
P
N
OOC(18:2)
C
-
H(18:2)COO (18:2)COO (20:3)COO-
N+
P
C
-
H-
CH-COO+
P
C
-
H-
-
--
OOC(18:3)
C H(16:0)COO
P
H
H
- 2C
C
- 2C
H OH
-
C
OOC(18:3)
H(16:0)COO
P-
P
C
-
H(18:3)COO-
inositol
P
HOH2C
C
-OH2C
H OH
OOC(18:3)
C
-
H(16:0)COO-
Phosphatidicacid (PA)
Diacylglycerol(DAG)
Cytidine diphosphatediacylglycerol (CDP-DAG)
Phosphatidyl-choline (PC)
Phosphatidyl-ethanolamine (PE)
Phosphatidyl-serine (PS)
Phosphatidyl- inositol (PI)
Phosphatidyl- glycerol (PG)
Diphosphatidyl-glycerol (DPG)
Sphingoid base (e.g. sphingosine)
Ceramide = sphingosine + amide-acyl (R)
Phosphocholine ceramide
Phosphoethanol ceramide
Cerebroside (e.g. monogalactosylcerebroside)
Squalene
Sterols (e.g. ergoserol occuringin Leishmania)
Glycerolipids
Main precursors:
Sphingolipids Sterols
Main precursors:Main precursors:
POH
RONH
OH
RONH
CH2 CH 2
CH2 CH 2
NH3+
NH3+
N+
CH3
CH3
CH3
CH3
P
P
: Fatty acid
Key:
: Phosphate
: Phospholipids (glycerolipids and sphingolipids)
CH3
CH3
CH3
H3C
H3C
N
CH2 OOC(18:2) OOC(18:3)CH2 OOC(18:0)CH2
CH2
CH2
CH2
CH2
CH2
CH2
CH3
H3 N H3CH3
H3C
CH2
CH2
CH2
CH2
CH2
CH2
CH2
CH2
CH2
CH2
TRENDS in Parasitology
O
Figure 1. Major classes of lipids of the membrane lipidome. The three main classes of membrane lipids (i.e. glycerolipids, sphingolipids and sterols) are shown with their
polar heads on the right. Acyls are shown in grey boxes. The main glycerolipids are assembled by esterification of the sn-1 and sn-2 positions of a glycerol backbone with
acyls and a polar head at position sn-3. These glycerolipids are neosynthesized from three precursors (PA, DAG and CDP-DAG). In some cases, fatty acid chains can be
linked via ether or vinyl ether linkages. Examples of acyl structures with various numbers of carbon and double bonds are indicated. Sphingolipids are assembled by
linkage of a fatty acid by an amide bond on a long-chain base, also called sphingoid base, such as sphingosine. Simple structures are shown but a variety of unique
Review Trends in Parasitology November 2011, Vol. 27, No. 11
498

Review Trends in Parasitology November 2011, Vol. 27, No. 11
soluble subunit of FASII, such as haloxyfop targetingacetyl-CoA carboxylase, cerulenin targeting FabF, thiolac-tomycin targeting FabF and FabH or triclosan developed totarget FabI would have to cross the plasma membrane ofthe host cell, the parasitophorous vacuole membrane, theplasma membrane of the parasite and the four membranessurrounding the apicoplast. As a result, in targeting thelipidome the chemical properties of the screened ordesigned drug candidates must be considered, which en-able the compound to circulate into hydrophilic and lipo-philic environments.
In this review, we introduce lipidomic strategies cur-rently applied to a large variety of parasites. Unique lipidstructures and biosynthetic pathways are pointed out thatare promising targets for novel drugs. Eventually, thequestion of the ability of a drug to find a target embeddedin a membrane and how lipid composition can alter theefficacy of developed drugs is addressed.
Technologies to characterize the lipidome of parasitesThe preparation of parasite samples for lipidomic
analyses
Lipids are small organic compounds characterized by theirhydrophobicity and molecular diversity. Lipidomicsaddresses their comprehensive analysis [8–14], requiringappropriate separation and analytical methods. The firstrequirement for the analysis of the parasite lipidome is toavoid contamination by lipids of the host. Pioneering lipi-domic analyses were based on the comparison of uninfectedand infected host cells [15]. Owing to the technologicalprogress in sensitivity, it is now possible to analyze sam-ples obtained after refined purification of parasites, despitea low yield [16,17]. Recent advances include the analyses ofparasite membrane organelles [18], subdomains such asrafts [17,19] or minor lipid classes such as glycosylpho-sphatidylinositols [20]. The presence of peripheral struc-tures protecting the parasites, such as cysts, eggshells andcuticles [21,22], require special procedures for lipid extrac-tion. Extraction of lipids from nematodes thus combinesfreezing and mechanical grinding before treatment bysolvents. The homogenates can then be analyzed by oneor a combination of the following methods, usually per-formed in specific technological platforms.
Sensitive analyses based on mass spectrometry
Lipidomic analyses heavily rely on the advances in massspectrometry (MS), either by coupling MS with anotherseparation method such as gas chromatography (GC-MS)or liquid chromatography (LC-MS) or by aligning massspectrometers (tandem MS) [23]. The detection and quan-tification of all the lipids in a series of samples is known aslipid profiling and can thus be performed with highthroughput using state-of-the-art, but still expensive,MS instruments [24]. The identification of molecular spe-cies that would vary between different sample groups isachieved by multivariate statistics to determine group-specific signals. Leishmania lipid profiling has thus been
glycosylated polar heads occurs in parasites. Sterols derive from the squalene structure
sterol acyl-esters. Phospholipids contain a phosphated polar head and can be glycero
diacylglycerol (DAG); phosphatidic acid (PA).
achieved with a very high-resolution and mass accuracy[24–26]. To characterize selected sets of lipid classes orsubclasses (e.g. phospholipidomics [27], sphingolipidomics[28]), tandem MS with a triple quadrupole is the method ofchoice, because after collision of analyzed molecules theloss of a polar-head fragment can generate a specific ionused as a diagnostic signature of a complete lipid class orsubclass, which can then be profiled in precursor ion scanmode. Alternatively, neutral loss scanning permits thedetection of lipids that have lost the same fragment follow-ing collision.
Although GC-MS is still used for fatty acids or sterols,LC-MS or direct infusion of crude lipid extracts into thespectrometer interface has become prominent in recentstudies [29]. The most common ionization techniques areelectrospray ionization (ESI) [30] and atmospheric pres-sure chemical ionization (APCI), although MALDI-TOF(matrix-assisted laser desorption/ionization coupled totime-of-flight mass spectrometry) is emerging in the lipi-domic field [31]. Shotgun lipidomics [32] is carried out bydirectly infusing crude lipid extracts in the ESI source.Separation of lipid classes is then based on intrinsic elec-trical properties. Shotgun lipidomics is inherently biasedtowards the most abundant and easily ionized lipids [26].The recent atmospheric pressure photo ionization (APPI)source was found of particular interest for non-polar lipids[33]. The efficiency of LC separation prior to the introduc-tion in the MS ion source plays an important role in thedetection of minor lipid species. The blood plasma lipidomewas thus assessed by ultra high pressure LC coupled withhigh-resolution fast-scanning MS, generating information-rich profiles [34]. This technique will undoubtedly be cru-cial for the analyses of parasites proliferating in bloodvessels.
High-performance thin-layer chromatography for
screens and metabolic labeling of parasites
To complement MS, thin layer chromatography (TLC)remains a robust technique to analyze lipids of knownstructures and screen for phenotypic variations [35]. Au-tomated high-performance TLC (HPTLC) [36], combinedwith the use of radiolabeled precursors, is a straightfor-ward method to analyze lipid syntheses inside parasiticcells, for a large array of functional studies [37–41], fromin vitro to in vivo conditions [40]. Radioactive precursorsmay thus allow the identification of essential lipid biosyn-thetic pathways that could be targeted. For example,radiolabeled fatty acids (FAs) have been used to showFA scavenging by Toxoplasma gondii [41], whereas radi-olabeled acetate could be incorporated into T. gondii acyl-lipids indicating the possibility of de novo FA biosynthesis[39]. Distinct conclusions can hence be drawn based onexperimental design. Accuracy of precursors for de novobiosynthesis of lipids should be selected with caution.Acetate has, for instance, been widely used as a precursorfor FA de novo synthesis in apicomplexan parasites [39,40]or apicomplexa-infected rodents [40]. In apicomplexans,
. A variety of sterols exist, including compounds linked to fatty acids in the case of
l or sphingolipids. Abbreviations: cytidine diphosphate-diacylglycerol (CDP-DAG);
499

Review Trends in Parasitology November 2011, Vol. 27, No. 11
the FA synthase is located inside an organelle called theapicoplast, but in this organelle, acetate cannot be activat-ed into acetyl-CoA, and no acetyl-CoA transporter is cur-rently known in the apicoplast. It is now considered thatphosphoenolpyruvate would be a more accurate precursorand that this metabolite should be used in future studies ofthis essential process in apicomplexans [42]. HPTLC isthus a rapid and efficient method to follow lipid syntheticfluxes, but results strongly depend on initial hypotheses,and the drawn conclusion should be supported by comple-mentary lines of evidence. Once a metabolic hypothesis hasbeen fully validated, HPTLC is also a method of choice toscreen for mutations or drugs impairing the lipidomicfluxes.
Imaging lipids and lipid domains in parasites
The subcellular localization of lipids can be analyzed bythree methods: (i) in Toxoplasma, fluorescent compounds, inthe form of fluorophore-conjugated lipids, have been used tomimic lipids diverted from host cells [41]. It is, however,difficult to assess whether the observed relocation of fluo-rescence reflects an in vivo process. (ii) The second method isbased on fluorescent proteins harboring lipid-bindingdomains [43,44]. Phosphatidylinositol 3-monophosphatewas studied in Toxoplasma using such a probe [45]. It shouldbe noted that the overexpression of a protein with a lipid-binding domain might result in changes in vivo, resulting inaltered interactions between the studied lipid and its natu-ral receptors. (iii) The third method is based on immuno-staining with specific antibodies, such as those raisedagainst glycolipid polar heads [17]. It is then important toassess the specificity of the antibody and take into accountthe possible aggregation of the visualized lipids that mightbe triggered by the antibody.
Imaging the lipid domains is also essential becausethese lipid structures, enriched in sterols, sphingolipidsand specific proteins, are suspected to play crucial roles inpathogenesis and drug resistance. Large proportions ofactive P-glycoproteins (Pgp), responsible for drug efflux,have been localized in raft-like structures (RLS) in nema-todes by confocal microscopy, and a relationship has beenestablished among RLS–Pgp complexes, xenobiotic trans-port and drug resistance [46].
Mining the lipidome of parasites to search for uniquelipid structuresIn the search for parasite-specific lipids, the analyses ofglycolipids has been one of the most prolific, pointing outglycosylglycerolipids such as phosphatidylinositols (GPIs)[47,48] or glycosylsphingolipids such as inositolphosphorylceramide (IPC) [49] and sulfoglycosphingolipids [50].Glycero- or sphingolipids terminated by non-mammaliana-Gal(1!6)b-Gal in Apicomplexans [17,38] or b-Gal(1!6)b-Gal in Trematoda parasites can thus inspire novel che-motherapeutic strategies as long as the precise structuresof the specific lipids and their biosynthetic pathways can beeventually elucidated [49]. In the shorter term, glycolipidsare excellent candidates for immunotherapeutic strategies[48,51]. Glycolipids seem ideal structures to search forparasite-specific lipids. With the gain in sensitivity ofanalytic methods, future research will have to focus on
500
other sources of molecular diversity, in particular at thelevel of the hydrophobic moiety of membrane lipids, such asthe fatty acid (which length can vary from 12 to more than26 carbon atoms, be even or odd numbered and be modifiedby addition of double bonds, oxygen atoms, cycles, etc.), thelong-chain base of glycerolipids and sphingolipids (whichlength can also vary), and sterols, which have not beensystematically characterized up to now.
Mining lipidome metabolism to search for unique lipidbiosynthetic pathwaysLimits of bioinformatic resources to mine lipid metabolic
pathways of parasites
Parasite lipid homeostasis results from four processes: (i)diversion from host; (ii) de novo syntheses; (iii) lipid con-versions; and (iv) trafficking (example in Figure 2 for theglycerolipidome). Regardless of the difficulty in annotatinggenomes of parasites [52], it is crucial to have access to themost complete metabolic maps to select probable targets.Unfortunately, databases such as KEGG [53] and MetaCyc[54] are far from being exploitable. KEGG provides globalmetabolic schemes, including metabolites, enzymes andlinks to drugs [53]. KEGG metabolic maps are available for14 Apicomplexa, one pathogenic Ciliate, five Kinetoplas-tida, one Trematoda and one pathogenic Nematoda. Com-paring maps of different organisms highlights differencesthat should be helpful to find target candidates. However,maps are designed based on enzymatic reactions and, forinstance, the map for FA synthesis is identical for FAsynthase of type I (FASI, a multiprotein complex) in thecytosol of mammalian cells and of type II (FASII, dissoci-ated enzymes) in the apicoplast of Apicomplexa. Further-more, FA synthesis in mammals combines the cytosolicFASI producing 16- to 18-carbon acyls, with mitochondrialFASII components producing 8-carbon precursors of lipo-ate. In the case of T. gondii containing both a cytosolicFASI and an apicoplast FASII, only the latter is shown.The molecular diversity of FAs (carbon length and desa-turations) is not shown. Trafficking is not represented, anda lipid generated in a given subcellular membrane can bemapped together with an enzyme localized in anothermembrane.
In contrast to KEGG, the MetaCyc view of metabolism isfragmented, reflecting knowledge gaps, and allowing thedesign of pathways that are unique to some phyla [54],including seven Apicomplexa, three pathogenic Ciliatesand one pathogenic Amoebozoa. Similar to KEGG, mem-brane compartmentalization is missing and molecular di-versity of FAs (carbon-chain length, unsaturation level,oxidation, etc.) is not shown. Alternative databases takinginto account the uniqueness of the metabolism of parasitesare thus badly needed. The best, if not sole example, is theMalaria Parasite Metabolic Pathways database [55]. Thequality is high, missing data are documented and enzymesubcellular localizations are shown. Nevertheless, similar toKEGG or MetaCyc, the in silico search of potential targets isdifficult in the context of lipid metabolism, because themolecular diversity of lipids, compartmentalization of path-ways and existence of alternative routes to generate identi-cal structures cannot be easily compared. Experimentalmetabolomic studies are therefore necessary.

FFA or glycerolipids from host
PLA?
Fatty acids
PA
G3P
LPA
FA scavenging from host?
Neosynthesis by a FASI?
Scavengingfrom host?
Neosynthesis by a FASII?
Neosynthesis by a FAE?
and / or
and / or
and / or
Pool of acyl-CoA/ACP
acyl(R1COO) – CoA/ACP
acyl(R1COO) – CoA/ACP AT1?
AT2?
Glycerolipidsfrom host
Glycerolipidsfrom host
Endomembranephospholipids
Polar headprecursor(s)
from host
*
*
* **
**
?
CDP – DAGDAG
PG
DPG
PIPSPEPC(MGDG)
(DGDG)
(SQDG) MT?
Scavengingfrom host?
PLC?
Scavengingfrom host?
Mitochondrialipids
Plastidlipids?
PPolar head-precursortransporter(s)?
***
***
Direct import?
Phospholipids from host
Glycerol or glycerolipidsfrom host
PLA?
Glycerol transporter?
PLD?
DAG transporter?
FA transporter?
PA transporter?
TRENDS in Parasitology
Figure 2. Targeting the membrane glycerolipidome of a unicellular parasite. Summary of the redundant pathways that a parasitic cell could harbor to synthesize a category
of lipids needed to build its cellular membranes, here the acyl-glycerolipids. The metabolism of glycerolipids is highly compartmentalized and if blocked at one point,
redundant routes might be used by the parasite to bypass the interrupted pathway. Redundant pathways combine scavenging of more or less elaborated precursors,
de novo syntheses and lipid conversions. The scavenging processes allow precursors to be imported into the parasite, probably due to transporters, generally after the
hydrolysis of host lipids by specific enzymes, the lipases, into free FAs, glycerol, diacylglycerol or phosphatidic acid. FAs can also be obtained by neosynthesis due to fatty
acid synthases or elongases. Phospholipids or phospholipid polar heads could also be diverted from the host. All these products feed several key pools to provide four main
groups of lipids (FAs, plastids lipids, endomembrane phospholipids and mitochondrial lipids). These pools include acyl-CoA or acyl-ACP that can be combined to glycerol-
3-phosphate by the action of two acyltransferases thus generating phosphatidic acid. Phosphatidic acid and diacylglycerol are the precursors for all diacyl-glycerolipids,
either by the DAG or by the CDP-DAG pathways, producing phospholipids (PS, PC, PE, PI, PG, DPG), glycolipids (MGDG, DGDG) or sulfoquinovosyldiacylglycerol found in
some plastid-containing cells. Once a specific feature has been detected in the lipidome of a parasite, it is thus essential to characterize the corresponding metabolic process
and assess that this process can be targeted and that the parasite will not bypass it, when chemically impaired. (*) FA scavenging from host has been demonstrated in
Apicomplexa, but parasites such as Plasmodium falciparum and Toxoplasma gondii also contain a FASII system that could also neosynthesize FAs; the presence of a FASI
system in T. gondii could be a third route to generate FAs. (**) Acyl transferases of the apicoplast might be potential targets for drugs to fight against malaria. (***) Import
of polar head precursors and methyltransferases converting ethanolamine into choline are currently used as targets to develop novel antimalarials (see text). Abbreviations:
acyl carrier protein (ACP); acyl-coenzyme A (Acyl-CoA); acyltransferases (AT1 and AT2); cytidine diphosphate-diacylglycerol (CDP-DAG); diacylglycerol (DAG);
diphosphatidylglycerol (DPG); fatty acid (FA); fatty acid elongases (FAE); fatty acid synthases I or II (FASI or FASII); free fatty acid (FFA); galactosyldiacylglycerol, mono
and di- (MGDG, DGDG); glycerol-3-phosphate (G3P); lysophosphatidic acid (LPA); methyl-transferases (MT); phosphatidic acid (PA); phosphatidylcholine (PC);
phosphatidylethanolamine (PE); phosphatidylglycerol (PG); phosphatidylinositol (PI); phosphatidylserine (PS); phospholipase A (PLA); phospholipase C (PLC);
phospholipase D (PLD); sulfoquinovosyldiacylglycerol (SQDG).
Review Trends in Parasitology November 2011, Vol. 27, No. 11
Absolute requirement of experimental validation: the
lessons of acyl-lipid drug development in Apicomplexa
The discovery of a plant-like plastid FASII in Apicomplex-ans such as Plasmodium or Toxoplasma [56] has stimulat-ed an intense search for possible drugs among herbicidesand antibiotics, such as thiolactomycin [56,57] or triclosan[40,58]. However, ambiguous and non-reproducible in vitroand in vivo results raised questions on the target validity.It was thus shown by genetic knockout that FASII could bedispensable in the blood stages of Plasmodium [59] andthat off-targets were probably responsible for measureddrug effects [60,61]. By contrast, the metabolism ofa ubiquitous glycerolipid (i.e. phosphatidylcholine) inPlasmodium has been dissected by various biochemical,bioinformatic and genetic approaches [7,62,63] and provedto be an ideal target for novel drugs [7,64], even inspiringthe design of drugs for other parasites in the Kinetoplas-
tida phylum [65]. Genomic clues should therefore be com-pleted by genetic and biochemical evidence.
Relative proportions of ubiquitous phospholipids suchas PC and PE also determine the general properties ofdrugs in membranes, a point addressed in the followingsection.
The question of drug dynamics in the membraneenvironmentDrug dynamics analyzed in membrane models
The interaction of a drug with the parasite plasma mem-brane is a prerequisite for its action. Accordingly, it isessential to investigate whether the specific properties ofthe parasite limiting membranes can alter the drug effi-ciency. Vesicles can be prepared from parasite plasmamembranes to measure salt permeability variations thatmight be provoked by a drug to assess the possibility of
501

Review Trends in Parasitology November 2011, Vol. 27, No. 11
drug-induced pore formation [66]. Drug–membrane inter-actions can, however, be dissected more precisely usingbiomimetic membrane models, including Langmuir mono-layers that are spontaneously formed at the air–waterinterface or immobilized artificial membranes in chro-matographic systems. A drug–lipid interaction can, forinstance, be studied using a monolayer composed of asingle lipid species [67]. However, based on a comprehen-sive lipidomic profiling, a biomimetic lipid monolayerreflecting a plasma membrane or a lipid raft compositioncan be analyzed, with a particular focus on differencesbetween wild type and drug-resistant parasites [67].
Amphotericin B was the first antiparasitic drug forwhich the interaction with lipids, and particularly sterols,was investigated using the Langmuir film [67]. The am-photericin B-induced membrane pore is responsible forion leakage, contributing to Leishmania and fungi death.Modeling studies of the amphotericin B–sterol channel,comparing relative affinity of amphotericin B for choles-terol and ergosterol, have shown a better stability and ahigher diameter of the pore when amphotericin B boundto ergosterol, present in fungi and Leishmania mem-branes and absent in mammals [68,69]. Miltefosine isan antileishmanial lipid-like drug for which an affinity forsterols was demonstrated by the Langmuir monolayertechnique, suggesting that membrane lipid rafts could bea miltefosine reservoir [70]. A miltefosine analog, edelfo-sine, was studied on the Langmuir monolayer, in combi-nation with amphotericin B, and this drug combinationresulted in a strong interaction between drugs, explain-ing a decrease in the antileishmanial activity of thiscombination [71]. The Langmuir monolayer techniquewas also useful in demonstrating that sitamaquine,an antileishmanial drug in development for which notransporter was found [72]; the drug accumulated inLeishmania donovani through passive diffusion, relyingon an electrical gradient [73].
Drug dynamics analyzed in living parasites
Nematodes are complex organisms that possess very thick,specific protective barriers (eggshells and cuticles) forwhich the exact biochemical composition is unknownand cannot be reconstructed. In addition to model mem-branes, the use of living cells or living nematodes repre-sents a primary and essential approach in understandingthe role of lipids in drug dynamics in these parasites. Allparasite membranes and barriers analyzed thus far innematodes include sterols. In vertebrates, cholesterol isthe main component of membranes and is known to mod-ulate the activity of pumps such as Pgp which are thoughtto be involved in drug efflux. Sterols could thus be used tomodulate resistance to antiparasitic drugs.
The role of cholesterol can be studied in living cells/organisms due to cholesterol/sterol acceptors such asmethyl-b-cyclodextrin which remove cholesterol frommembranes. The adaptation of this technique to nema-todes showed that sterols could also alter eggshells andmembranes, consequently changing the activity of Pgpand the susceptibility to drugs in nematodes [22,35,74]or unicellular parasites [70,75]. Depletion of cholesterol inLeishmania or nematodes consequently increases drug
502
resistance while cholesterol loading decreases it [22,75].The decrease in cholesterol concentration was associatedwith the highest drug efflux. Moreover, a significant dif-ference in sterol concentration was found between nema-todes susceptible or resistant to anthelmintics, the latterhaving a naturally lower sterol concentration [22,35].
The interplay between cholesterol and phospholipids isalso crucial for drug dynamics. Phospholipids classes arethe same in nematodes and in vertebrates. Regardless ofthe fact that FAs within the same classes of lipids can varygreatly between the phyla, a significant relationship be-tween lipid class composition and drug resistance wasnevertheless found to depend on the equilibrium betweenfree cholesterol and phospholipids with PC, PE and phos-phatidic acid having the most significant impacts [35].
Manipulating lipid composition of nematodes and elu-cidating sterol pathways and the balance with phospholi-pids can thus help improve drug efficiency and the designof more potent antiparasitic strategies. Results obtainedwith several nematode species, parasites of plants oranimals, showed that plant oils can be antiparasitic oradjuvants to antiparasitic drugs [74], supporting this per-spective for future drug developments.
Concluding remarksThe spectacular technological advances in high sensitivityMS, HPTLC and high-resolution imaging has acceleratedthe structural and functional characterization of the lipi-dome of parasites, ranging from unicellular protists toworms, which cause threatening diseases in vertebrates,including humans. The search for specific lipid structureshas proven efficient to determine unique glycosylated polarheads in parasite membrane lipids, and the current chal-lenge is to expand this search to identify specific substruc-tures in hydrophobic moieties of membrane lipids such asacyls, long-chain bases and sterols. The next step is theassessment that the specific feature detected in the para-site lipidome is indeed vital for the pathogen. When vali-dated by metabolic and functional studies, the specificity ofthe parasite lipidome can then inspire the development ofnovel drugs. The definition of precise lipidomic profiles alsoallow the reconstitution of biomimetic systems to study thedynamics and bioavailability of developed drugs in para-site limiting membranes, giving clues concerning the re-sistance that could arise from membrane lipid remodeling.Future developments will therefore strongly depend on theprogresses in sensitivity and throughput of lipidomic tech-nologies made available to the scientific community, main-ly by the access to specific technological platforms.
References1 Maurya, R. et al. (2010) Evaluation of blood agar microtiter plates for
culturing Leishmania parasites to titrate parasite burden in spleen andperipheral blood of patients with visceral leishmaniasis. J. Clin.Microbiol. 48, 1932–1934
2 Dietz, K. et al. (2006) Mathematical model of the first wave ofPlasmodium falciparum asexual parasitemia in non-immune andvaccinated individuals. Am. J. Trop. Med. Hyg. 75, 46–55
3 Geraghty, E.M. et al. (2007) Overwhelming parasitemia withPlasmodium falciparum infection in a patient receiving infliximabtherapy for rheumatoid arthritis. Clin. Infect. Dis. 44, e82–e84
4 Loverde, P.T. and Chen, L. (1991) Schistosome female reproductivedevelopment. Parasitol. Today 7, 303–308

Review Trends in Parasitology November 2011, Vol. 27, No. 11
5 Bethony, J. et al. (2006) Soil-transmitted helminth infections:ascariasis, trichuriasis, and hookworm. Lancet 367, 1521–1532
6 Saidani, N. et al. (2009) Potential and limits of in silico target discovery– case study of the search for new antimalarial chemotherapeutictargets. Infect. Genet. Evol. 9, 359–367
7 Ben Mamoun, C. et al. (2010) Targeting the lipid metabolic pathwaysfor the treatment of malaria. Drug Dev. Res. 71, 44–55
8 Shevchenko, A. and Simons, K. (2010) Lipidomics: coming to grips withlipid diversity. Nat. Rev. Mol. Cell Biol. 11, 593–598
9 Dennis, E.A. (2009) Lipidomics joins the omics evolution. Proc. Natl.Acad. Sci. U.S.A. 106, 2089–2090
10 Kita, Y. et al. (2006) Lipidomics approaches for a comprehensive analysisof fatty acids and metabolites by LC/ESI-MS. Prostaglandins Other LipidMediat. 79, 150–151
11 Wenk, M.R. (2005) The emerging field of lipidomics. Nat. Rev. DrugDiscov. 4, 594–610
12 McKee, D.L. (2003) Lipidomics. Scientist 17, 2013 Lagarde, M. et al. (2003) Lipidomics is emerging. Biochim. Biophys.
Acta 1634, 6114 Feng, L. et al. (2003) Lipidomics: probing lipid-protein interactions
using immobilized lipids. FASEB J. 17, A993–A99415 Hsiao, L.L. et al. (1991) Modification of host cell membrane lipid
composition by the intra-erythrocytic human malaria parasitePlasmodium falciparum. Biochem. J. 274, 121–132
16 Welti, R. et al. (2007) Lipidomic analysis of Toxoplasma gondii revealsunusual polar lipids. Biochemistry 46, 13882–13890
17 Botte, C. et al. (2008) Subcellular localization and dynamics of adigalactolipid-like epitope in Toxoplasma gondii. J. Lipid Res. 49,746–762
18 Besteiro, S. et al. (2009) Export of a Toxoplasma gondii rhoptry neckprotein complex at the host cell membrane to form the moving junctionduring invasion. PLoS Pathog. 5, e1000309
19 Azzouz, N. et al. (2006) Toxoplasma gondii grown in human cells usesGalNAc-containing glycosylphosphatidylinositol precursors to anchorsurface antigens while the immunogenic Glc-GalNAc-containingprecursors remain free at the parasite cell surface. Int. J. Biochem.Cell Biol. 38, 1914–1925
20 Azzouz, N. et al. (2005) Removal of phospholipid contaminants throughprecipitation of glycosylphosphatidylinositols. Anal. Biochem. 343,152–158
21 Riou, M. et al. (2005) Immunolocalisation of an ABC transporter,P-glycoprotein, in the eggshells and cuticles of free-living andparasitic stages of Haemonchus contortus. Parasitol. Res. 96, 142–148
22 Riou, M. et al. (2003) Modulation of the multidrug resistance (MDR)system in the nematode Haemonchus contortus by changing cholesterolcontent: effects on resistance to anthelmintics. J. Antimicrob.Chemother. 52, 180–187
23 Navas-Iglesias, N. et al. (2009) From lipids analysis towardslipidomics, a new challenge for the analytical chemistry of the 21stcentury. Part II: analytical lipidomics. Trends Anal. Chem. 28, 393–403
24 Schwudke, D. et al. (2007) Top-down lipidomic screens by multivariateanalysis of high-resolution survey mass spectra. Anal. Chem. 79,4083–4093
25 Zheng, L. et al. (2010) Profiling of lipids in Leishmania donovani usinghydrophilic interaction chromatography in combination with Fouriertransform mass spectrometry. Rapid Commun. Mass Spectrom. 24,2074–2082
26 Griffiths, W.J. and Wang, Y. (2009) Mass spectrometry: from proteomicsto metabolomics and lipidomics. Chem. Soc. Rev. 38, 1882–1896
27 Willmann, J. et al. (2008) Hyphenated tools for phospholipidomics.J. Biomol. Tech. 19, 211–216
28 Haynes, C.A. et al. (2009) Sphingolipidomics: methods for thecomprehensive analysis of sphingolipids. J. Chromatogr. B: Anal.Technol. Biomed. Life Sci. 877, 2696–2708
29 Roberts, L.D. et al. (2008) A matter of fat: an introduction to lipidomicprofiling methods. J. Chromatogr. B: Anal. Technol. Biomed. Life Sci.871, 174–181
30 Lehtonen, M. et al. (2011) Determination of endocannabinoids innematodes and human brain tissue by liquid chromatographyelectrospray ionization tandem mass spectrometry. J. Chromatogr.B: Anal. Technol. Biomed. Life Sci. 879, 677–694
31 Schiller, J. et al. (2007) MALDI-TOF MS in lipidomics. Front. Biosci.12, 2568–2579
32 Han, X. and Gross, R.W. (2005) Shotgun lipidomics: electrosprayionization mass spectrometric analysis and quantitation of cellularlipidomes directly from crude extracts of biological samples. MassSpectrom. Rev. 24, 367–412
33 Cai, S.S. et al. (2007) Comparison of atmospheric pressurephotoionization and atmospheric pressure chemical ionization fornormal-phase LC/MS chiral analysis of pharmaceuticals. Anal.Chem. 79, 2491–2498
34 Sandra, K. et al. (2010) Comprehensive blood plasma lipidomics byliquid chromatography/quadrupole time-of-flight mass spectrometry.J. Chromatogr. A 1217, 4087–4099
35 Riou, M. et al. (2007) Relationships between sterol/phospholipidcomposition and xenobiotic transport in nematodes. Parasitol. Res.100, 1125–1134
36 Fuchs, B. et al. (2010) Lipid analysis by thin-layer chromatography – areview of the current state. J. Chromatogr. A 1218, 2754–2774
37 Azzouz, N. et al. (2008) Metabolic labeling and structural analysis ofglycosylphosphatidylinositols from parasitic protozoa. Methods Mol.Biol. 446, 183–198
38 Marechal, E. et al. (2002) Synthesis of chloroplast galactolipids inapicomplexan parasites. Eukaryot. Cell 1, 653–656
39 Bisanz, C. et al. (2006) Toxoplasma gondii acyl-lipid metabolism: denovo synthesis from apicoplast-generated fatty acids versusscavenging of host cell precursors. Biochem. J. 394, 197–205
40 Surolia, N. and Surolia, A. (2001) Triclosan offers protection againstblood stages of malaria by inhibiting enoyl-ACP reductase ofPlasmodium falciparum. Nat. Med. 7, 167–173
41 Charron, A.J. and Sibley, L.D. (2002) Host cells: mobilizable lipidresources for the intracellular parasite Toxoplasma gondii. J. CellSci. 115, 3049–3059
42 Mazumdar, J. et al. (2006) Apicoplast fatty acid synthesis is essentialfor organelle biogenesis and parasite survival in Toxoplasma gondii.Proc. Natl. Acad. Sci. U.S.A. 103, 13192–13197
43 Lemmon, M.A. (2003) Phosphoinositide recognition domains. Traffic 4,201–213
44 Backer, J.M. (2010) New methods for capturing the mystery lipid,PtdIns5P. Biochem. J. 428, e1–e2
45 Tawk, L. et al. (2010) Phosphatidylinositol 3-monophosphate isinvolved in toxoplasma apicoplast biogenesis. PLoS Pathog. 7,e1001286
46 Riou, M. et al. (2010) Drug resistance is affected by colocalizationof P-glycoproteins in raft-like structures unexpected in eggshellsof the nematode Haemonchus contortus. Biochem. Cell Biol. 88,459–467
47 Ferguson, M.A. et al. (1994) Glycosyl-phosphatidylinositol molecules ofthe parasite and the host. Parasitology 108 (Suppl.), S45–S54
48 Debierre-Grockiego, F. et al. (2010) Binding of Toxoplasma gondiiglycosylphosphatidylinositols to galectin-3 is required for theirrecognition by macrophages. J. Biol. Chem. 285, 32744–32750
49 Sonda, S. and Hehl, A.B. (2006) Lipid biology of Apicomplexa:perspectives for new drug targets, particularly for Toxoplasmagondii. Trends Parasitol. 22, 41–47
50 Landoni, M. et al. (2007) Plasmodium falciparum biosynthesizessulfoglycosphingolipids. Mol. Biochem. Parasitol. 154, 22–29
51 Boutlis, C.S. et al. (2002) Antibodies to Plasmodium falciparumglycosylphosphatidylinositols: inverse association with tolerance ofparasitemia in Papua New Guinean children and adults. Infect.Immun. 70, 5052–5057
52 Birkholtz, L.M. et al. (2006) Integration and mining of malariamolecular, functional and pharmacological data: how far are wefrom a chemogenomic knowledge space? Malar. J. 5, 110
53 Kanehisa, M. et al. (2010) KEGG for representation and analysis ofmolecular networks involving diseases and drugs. Nucleic Acids Res.38, D355–D360
54 Caspi, R. et al. (2010) The MetaCyc database of metabolic pathwaysand enzymes and the BioCyc collection of pathway/genome databases.Nucleic Acids Res. 38, D473–D479
55 Ginsburg, H. and Tilley, L. (2011) Plasmodium falciparum metabolicpathways (MPMP) project upgraded with a database of subcellularlocations of gene products. Trends Parasitol. 27, 285–286
56 Waller, R.F. et al. (2003) A type II pathway for fatty acid biosynthesispresents drug targets in Plasmodium falciparum. Antimicrob. AgentsChemother. 47, 297–301
503

Review Trends in Parasitology November 2011, Vol. 27, No. 11
57 Prigge, S.T. et al. (2003) The initiating steps of a type II fatty acidsynthase in Plasmodium falciparum are catalyzed by pfACP, pfMCAT,and pfKASIII. Biochemistry 42, 1160–1169
58 McLeod, R. et al. (2001) Triclosan inhibits the growth of Plasmodiumfalciparum and Toxoplasma gondii by inhibition of apicomplexan FabI. Int. J. Parasitol. 31, 109–113
59 Yu, M. et al. (2008) The fatty acid biosynthesis enzyme FabI plays a keyrole in the development of liver-stage malarial parasites. Cell HostMicrobe 4, 567–578
60 Baschong, W. et al. (2011) Triclosan is minimally effective in rodentmalaria models. Nat. Med. 17, 33–34 34–35 author reply
61 Surolia, A. and Surolia, N. (2011) Triclosan is minimally effective inrodent malaria models – reply. Nat. Med. 17, 34–35
62 Besteiro, S. et al. (2010) Exploring metabolomic approaches to analysephospholipid biosynthetic pathways in Plasmodium. Parasitology 137,1343–1356
63 Dechamps, S. et al. (2010) Rodent and nonrodent malaria parasitesdiffer in their phospholipid metabolic pathways. J. Lipid Res. 51, 81–96
64 Caldarelli, S.A. et al. (2010) Exploration of potential prodrug approachof the bis-thiazolium salts T3 and T4 for orally delivered antimalarials.Bioorg. Med. Chem. Lett. 20, 3953–3956
65 Ibrahim, H.M. et al. (2010) Symmetrical choline-derived dicationsdisplay strong anti-kinetoplastid activity. J. Antimicrob. Chemother.66, 111–125
66 Cohen, B.E. (1986) Concentration- and time-dependence ofamphotericin-B induced permeability changes across ergosterol-containing liposomes. Biochim. Biophys. Acta 857, 117–122
504
67 Rakotomanga, M. et al. (2005) Alteration of fatty acid andsterol metabolism in miltefosine-resistant Leishmania donovanipromastigotes and consequences for drug-membrane interactions.Antimicrob. Agents Chemother. 49, 2677–2686
68 Baginski, M. et al. (2002) Comparative molecular dynamicssimulations of amphotericin B-cholesterol/ergosterol membranechannels. Biochim. Biophys. Acta 1567, 63–78
69 Resat, H. and Baginski, M. (2002) Ion passage pathways andthermodynamics of the amphotericin B membrane channel. Eur.Biophys. J. 31, 294–305
70 Saint-Pierre-Chazalet, M. et al. (2009) Membrane sterol depletionimpairs miltefosine action in wild-type and miltefosine-resistantLeishmania donovani promastigotes. J. Antimicrob. Chemother. 64,993–1001
71 Hac-Wydro, K. et al. (2009) Langmuir monolayer study towardcombined antileishmanian therapy involving amphotericin B andedelfosine. J. Phys. Chem. B 113, 14239–14246
72 Lopez-Martin, C. et al. (2008) Sitamaquine sensitivity in Leishmaniaspecies is not mediated by drug accumulation in acidocalcisomes.Antimicrob. Agents Chemother. 52, 4030–4036
73 Duenas-Romero, A.M. et al. (2007) Interaction of sitamaquine withmembrane lipids of Leishmania donovani promastigotes. Biochim.Biophys. Acta 1768, 246–252
74 Kerboeuf, D. et al. (2010) Membrane drug transport in helminths. Anti-Infective Agents Med. Chem. 9, 113–129
75 Barratt, G. et al. (2009) Cellular transport and lipid interactions ofmiltefosine. Curr. Drug Metab. 10, 247–255