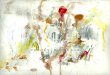Embed Size (px)
Citation preview

Sexologies 15 (2006) 241–249
ava i lab le at www.sc ienced i rect .com
journa l homepage: ht tp://f rance.e l sev ier.com/di rect/sexo l
ARTICLE ORIGINAL / ORIGINAL ARTICLE
Modèles précliniques du désir sexuel
Preclinical models of sexual desire
T. Spiteri (MSc), A. Ågmo (PhD) *
Institut de psychologie, université de Tromsø, 9037 Tromsø, Norvège
Disponible sur internet le 22 juin 2006
11do
MOTS CLÉSDésordre sexuel ;Désir sexuel ;Motivation sexuelle ;Modèle préclinique ;IRSS ;Bupropion
* Auteur correspondant.Adresse e-mail : andersa@
58-1360/$ - see front mattei:10.1016/j.sexol.2006.05.0
psyk.uit
r © 20001
Résumé Dans le cadre de l’analyse des modèles précliniques du comportement sexuel nousproposons que le concept de désir sexuel chez l’humain soit équivalent à celui de la motiva-tion sexuelle tel qu’il est utilisé dans la littérature scientifique. Beaucoup de tests de la moti-vation sexuelle chez les animaux ont été décrits au cours des années. La plupart d’entre euxsont fondés sur l’évaluation du taux ou de la vitesse d’exécution de réponses motrices appri-ses lors d’un conditionnement opérant. Pour de nombreuses raisons évoquées dans cet article,ce ne sont pas des mesures idéales pour évaluer l’intensité de la motivation sexuelle. Nousprésentons un test de la motivation sexuelle qui a été employé chez les rats mâles et femel-les. Ce test a l’avantage d’être peu sensible aux variations de l’activité motrice, de ne pasutiliser de mesure de taux de réponse et de ne nécessiter aucun apprentissage. Le manquede traitements établis et médicalement efficaces pour des désordres du désir sexuel rend dif-ficile l’évaluation de la validité prédictive et corrélationnelle de n’importe quel modèle.Néanmoins, le modèle proposé ici pourrait être isomorphe et homologue au désir sexuel chezl’humain.© 2006 Elsevier Masson SAS. Tous droits réservés.
KEYWORDSSexual disorders;Sexual desire;Sexual motivation;Preclinical models;SSRI;Bupropion
Abstract Within the context of preclinical models of sexual behavior we suggest that the hu-man concept of sexual desire is equivalent to the concept of sexual motivation as used in theexperimental literature. Many tests of sexual motivation in non-human animals have been de-scribed. Most of these are based on the speed or rate of execution of learned responses, par-ticularly learned operants. For several reasons outlined in this article, such procedures are notideal for the determination of the intensity of sexual motivation. We describe here a simpletest for sexual motivation that has been employed both in male and female rats. It has the ad-vantage of being rather insensible to variations in motor activity or general arousal, it doesnot employ measures of response speed and it does not require learning. The absence of es-tablished treatments for human sexual desire disorders makes it difficult to determine thepredictive or correlational validity of any preclinical model. Nevertheless, the model proposedhere may be both isomorph and homologous to human sexual desire.© 2006 Elsevier Masson SAS. Tous droits réservés.
.no (A. Ågmo).
6 Elsevier Masson SAS. Tous droits réservés.

T. Spiteri, A. Ågmo242
Abridged version
Human sexual dysfunctions are increasingly attractingattention within the medical profession. The success obtai-ned with pharmacological treatments of erectile deficien-cies has certainly contributed to this, but there are alsoother important reasons. Among these are a shift in focusfrom male to female dysfunctions, and recognition of thebasic role of sexual desire. It is probably so that some ormany problems of arousal (erection, vaginal lubrication)may be related to low or absent sexual desire. Further-more, epidemiological data have shown that hypoactivesexual desire is by large the most common sexual dysfunc-tion in women. Indeed, its prevalence is above that of erec-tile dysfunction, even when compared to the prevalence ofthat disorder in older men. At the same time, there aredata suggesting that male sex offenders may have an unu-sually high level of sexual desire. In view of the concernthat many societies show about sexual offenses, an effi-cient and humane treatment reducing that high desire isbeing sought for.
Because of the reasons outlined in the preceding para-graph, the clinical usefulness of treatments modifyingsexual desire is regarded as beyond doubt. An attractivetreatment alternative appears to be drugs with prosexualor antisexual effects. For the development of clinically effi-cient drugs, animal models predictive of drug actions in thehuman are essential. Thus, the enhanced interest in poten-tial treatments for human sexual desire disorders has led toa demand for animal models of desire with established pre-dictive validity. In addition, some believe that studies ofnon-human sexual behavior could reveal the mechanismsdetermining the intensity of sexual desire, and that suchknowledge could facilitate the discovery of efficient treat-ment. This supposition does not appear to be entirelyunreasonable.
Most research on non-human sexual behavior has tradi-tionally focussed on the act of copulation. This act is highlystereotyped and often species specific, and consists in therepeated activation of somatic and/or autonomic reflexes.In rodents, the favored subject for animal sex behavior stu-dies, the male performs a series of mounts with pelvicthrusting, associated or not with vaginal penetration andeventually ejaculation. The motor pattern displayed isextremely stereotyped. Likewise, the essential element ofthe female rodent’s sexual behavior, the lordosis, is a fixedaction pattern showing very little variation. In contrast,human copulatory behavior is quite flexible, includingpenetration of orifices other than the vaginal and whereefficient genital stimulation can be produced by almostany part of the body. The stereotyped and species specificnature of copulatory behavior makes it unsuitable as amodel.
At difference from copulation, there is another part ofsexual behavior that is completely arbitrary, and variablewithin the individual and among individuals: Initialapproach to a potential mate. Copulation is not possibleat a distance, making proximity a requisite for any copula-tory behavior. The intensity of approach behaviors is suppo-sed to be determined by the intensity of the urge to esta-blish sexual interaction. This urge can conveniently be
called sexual motivation, and we maintain that the conceptof sexual motivation, as used in the animal literature, isequivalent to the concept of desire. An animal model offe-ring reasonable estimations of the intensity of approachbehaviors could, then, offer valuable information aboutsexual motivation, hence desire.
One procedure for evaluating the intensity of approachbehaviors have been employed in both male and femalerats, and data have been obtained suggesting that it indeedquantifies sexual motivation in a reliable way. Whether ithas predictive validity or not is an open question, butsome coincidence between drug effects in the human anddrug effects in the model is suggestive. For example, speci-fic serotonin reuptake inhibitors have sometimes delete-rious effects on sexual desire in humans, and they havesimilar effects in the model. Treatments interfering withgonadal hormones are known to reduce sexual desire inhumans, and they are doing the same in the model. Thelack of established prosexual drugs in the human has madeit impossible to determine if the model also is sensitive tomanipulations enhancing sexual motivation.
In addition to physiologically or pharmacologically indu-ced changes, we have determined how learning may modifysexual motivation in the procedure. The association ofsexual activity with an aversive event completely elimina-tes sexual motivation. Although data on learned modifica-tions of human sexual desire are scarce, the potentialimportance of learning for human sexual desire disordersshould not be underestimated, and the procedure outlinedhere could provide some information about basic learningmechanisms.
Version longue
Introduction
Durant l’histoire de la recherche sur le comportementsexuel animal, il n’y a eu que de rares efforts sérieux visantà généraliser, de manière explicite, les résultats des sujetsnon humains à l’humain (voir Ågmo, 1999 ; Ågmo et Elling-sen, 2003 pour une discussion plus approfondie). Une desraisons à ce constat est, sans doute, que l’un des cher-cheurs les plus influents dans l’étude du comportementsexuel, Frank Beach, considérait le comportement sexuelnon humain comme très dépendant des hormones sexuelleset fortement stéréotypé alors que le comportement sexuelhumain l’était beaucoup moins (Beach, 1947a, b). Dans cesconditions les généralisations de l’animal à l’humain n’ontpas pu facilement être faites. En outre, la recherche sur lecomportement sexuel animal s’est concentrée sur les réfle-xes hormonodépendants tels que la lordose chez les femel-les et la monte avec poussée pelvienne (habituellementassociée à la pénétration et, par la suite, à l’éjaculation)chez les mâles. De tels réflexes sont, en effet, tous deuxfortement stéréotypés et spécifiques, rendant toute géné-ralisation extrêmement difficile voire impossible. Toutefois,cela n’a jamais été réellement considéré comme un pro-blème car une grande partie du travail visait et vise tou-jours à comprendre la manière dont les hormones agissentsur le cerveau aussi bien au niveau cellulaire que molécu-laire, et à savoir dans quelles structures du cerveau elles le

Modèles précliniques du désir sexuel 243
font. L’élucidation de la façon dont les estrogènes régissentle réflexe de lordose en est un bel exemple (résumé dansPfaff, 1999).
Cette situation a changé durant les dernières années.L’intérêt croissant pour les désordres sexuels humains etleurs traitements a créé un besoin en modèles animaux fia-bles de la sexualité humaine. L’attention commence égale-ment à s’éloigner des éléments réflexes du comportementsexuel, comme l’érection, pour s’intéresser à des formesplus complexes de comportement volontaire comme ledésir sexuel et ses manifestations. Dans cette revue, nousprouverons que l’intensité de certains comportements pré-cédant la copulation est déterminée par un mécanisme quenous pouvons facilement appeler la motivation sexuelle etqu’elle peut être considérée comme équivalente auconcept de désir sexuel utilisé chez l’homme. Nous montre-rons alors que l’intensité de la motivation sexuelle peutêtre expérimentalement évaluée en présentant quelquesdonnées chez les rats mâles et femelles. Cependant, lefait qu’elle puisse être mesurée chez l’animal et qu’ellepuisse être équivalente au désir sexuel chez l’hommen’est pas suffisant pour supposer que les données animalespuissent être généralisées sans ambiguïté aux humains. Leproblème devient toujours plus complexe si nous voulonsexaminer, chez les animaux, des traitements pharmacologi-ques ou non, destinés aux humains. Ainsi, les modèles ani-maux des désordres humains doivent satisfaire un certainnombre de critères que nous étudierons précisément danscet article.
Pourquoi a-t-on besoin d’un modèle préclinique ?
Le succès énorme autant qu’inattendu des médicamentsdans le traitement des déficiences érectiles, à savoir du sil-dénafil (Viagra®) et de ses successeurs, du verdanafil(Levitra®) et du tadalafil (Cialis®), a été un facteur décisif.Ces produits ont, non seulement, rendu évident l’efficacitéde certains médicaments dans l’amélioration des fonctionssexuelles mais aussi que de nombreuses personnes sont dis-posées à payer des sommes élevées pour les obtenir. Les2,5 milliards de dollars américains de ventes cumulées desildénafil, de verdanafil et de tadalafil en 2004, l’illustrentbien. Si des médicaments améliorant l’érection peuventavoir un tel succès, il n’est pas impossible qu’il en soit demême pour des médicaments modifiant le désir sexuel. Eneffet, les études épidémiologiques (Arnal et al., 1995 ;Fugl-Meyer et Sjögren-Fugl-Meyer, 1999 ; Laumann et al.,1999) ont démontré que la prévalence du désordre dedésir sexuel hypoactif chez les femmes est largement supé-rieure à celle de l’impuissance chez les hommes, indiquantainsi un marché potentiel considérable. De plus, il existe uncertain nombre d’hommes manifestant une activitésexuelle qui n’est acceptable, ni socialement, ni personnel-lement. On trouve parmi eux les paraphiliaques, pour les-quels un traitement pharmacologique visant à la réductiond’une motivation sexuelle trop élevée pourrait donc êtresouhaitable. Ce type de raisonnement a stimulé une recher-che sur des médicaments pouvant augmenter ou bloquersélectivement la motivation sexuelle. Cette rechercheexige des modèles animaux fiables.
Critères pour un bon modèle préclinique
La corrélationUn des critères corrélationnels est que le modèle animaldevrait être sensible aux composés standard connus pourêtre thérapeutiquement efficace chez l’homme. D’ailleurs,l’activité relative des médicaments connus devrait êtresemblable chez l’humain et dans le modèle animal. Ildevrait également être sélectif de sorte que les médica-ments inefficaces chez l’humain devraient également êtreinefficaces dans le modèle animal. Cet ensemble de critè-res est fortement utile dans les modèles animaux de l’an-goisse, de la schizophrénie, de la dépression, etc., là oùune grande quantité de médicaments efficaces existe etest validée. Malheureusement tel n’est pas le cas pour ledésir sexuel hypoactif où il n’existe aucun médicamentavec un effet clinique prouvé. Comme le suggèrent certai-nes données préliminaires, le bupropion pourrait être clini-quement actif chez la femme mais c’est loin d’être établi.Même si cela s’avérait exact, il serait, d’abord, très diffi-cile d’analyser des corrélations à partir de ce seul médica-ment, et ensuite, de démontrer l’activité et la sélectivitérelatives. L’inévitable conclusion est que des critères corré-lationnels traditionnels ne sont pas facilement appliquésaux modèles animaux du désir sexuel.
Cependant, il existe quelques manipulations expérimen-tales qui réduisent la motivation sexuelle chez les mammi-fères non-humains : l’élimination des hormones gonadiqueset/ou de leurs actions cellulaires. Chez l’homme, de tellesmanipulations ont des effets forts semblables (voir Ågmo etEllingsen, 2003 pour des références). Cela offre une possi-bilité d’établir une certaine validité corrélationnelle.
L’isomorphismeL’adéquation d’un modèle préclinique peut également êtrejugée par l’isomorphisme, c’est-à-dire la similitude entreles comportements chez l’homme et l’animal. Par simili-tude, on ne veut, évidemment, pas dire que les patronsmoteurs sont exactement identiques ou même équivalentsmais qu’ils visent le même résultat ou le même but. Dans lecas du désir sexuel ou de la motivation sexuelle, le résultatou le but serait l’approche réussie d’un compagnon et l’éta-blissement d’un contact menant, par la suite, à la copula-tion. Les procédures pour évaluer l’intensité des comporte-ments d’approche et la transition de l’approche à lacopulation sont aisément disponibles chez les animaux.Ainsi, on peut prétendre qu’au moins certains modèles pré-cliniques du désir sexuel sont isomorphes.
L’homologieL’idéal, pour qu’un modèle préclinique soit considérécomme tel, est qu’il soit homologue. C’est-à-dire que lescauses fondamentales du comportement ou les processuscomportementaux sont semblables chez l’humain et dansle modèle animal. Il y a de nombreuses raisons de croireque les mécanismes neuraux et comportementaux de basedéterminant le désir ou la motivation sexuelle sont sembla-bles chez les rongeurs et chez l’homme (voir Ågmo et al.,2004 pour une discussion). De ce point de vue les modèlesde rongeurs peuvent être considérés comme homologues.Cependant, la catégorie du désir sexuel hypoactif n’estpas facilement définie chez les animaux. Néanmoins, une

T. Spiteri, A. Ågmo244
faible motivation sexuelle peut facilement être produitechez les animaux grâce à des procédures expérimentales(voir ci-dessous). Il y a également une proportion substan-tielle de rats montrant un faible désir sexuel toute leur vie,à savoir qu’ils ne réalisent aucun comportement copula-toire dans des conditions de test standard (Whalen, 1964 ;Whalen et al., 1961).
L’isomorphisme et le choix du comportementà observer
Chez la plupart des animaux dont les rongeurs, la copula-tion est essentiellement limitée à l’activation répétéed’une série de réflexes. Au contraire, la copulation chezl’humain n’est que partiellement déterminée par des mou-vements réflexes. Évidemment, le réflexe autonome del’érection est nécessaire pour des rapports vaginaux et lalubrification vaginale est utile mais le comportement copu-latoire réel peut être fortement variable. D’ailleurs, lesactivités sexuelles autres que des rapports vaginaux, nedépendent pas obligatoirement de l’érection ou de la lubri-fication vaginale. Il semble donc, que le comportementcopulatoire n’est pas nécessairement isomorphe chez leshumains et chez les rats. Cela signifie que l’étude de lacopulation chez l’animal n’est pas toujours la plus appro-priée pour l’extrapolation de l’animal à l’humain.
Le comportement d’approcheAu lieu d’étudier le comportement copulatoire, nous pou-vons nous intéresser aux événements précédant la copula-tion. Le comportement copulatoire ne peut avoir lieu quelorsque deux (ou plus) individus sont très proches. Dans lestests standard l’expérimentateur assure donc, cette proxi-mité en confinant des individus dans une petite cage.Cependant, dans un environnement naturel, la premièrecondition pour l’interaction sexuelle est la localisation etl’approche au partenaire potentiel. Les comportementsd’approche ne sont pas stéréotypés mais fortement varia-bles et déterminés par le contexte immédiat. Ainsi, l’ap-proche ne se compose pas d’un modèle moteur spécifiquemais de mouvements arbitraires menant à une réduction dela distance avec ce partenaire potentiel. L’intensité decette approche au partenaire est une mesure de l’intensitéde la motivation incitative. Nous avons déjà argué du faitque la motivation sexuelle est un cas prototypique de lamotivation incitative (Ågmo, 2003 ; Pfaff et Ågmo, 2002).D’ailleurs, c’est parce que ces comportements d’approchesont arbitraires et, par conséquent, les plus variables, maisayant le but commun chez les animaux et chez les humains,d’établir la proximité avec le partenaire, qu’ils peuventêtre considérés comme isomorphes. Les études de l’inten-sité de l’approche du partenaire sont donc, idéales pourévaluer la motivation sexuelle.
Problèmes des tests demandant une réponse motriceBeaucoup de tests de la motivation sexuelle incitative utili-sent des réponses induites d’un conditionnement opéranttelles que courir sur une piste ou appuyer sur un levierdans une boîte de Skinner pour accéder au partenaire (parexemple, López et al., 1999 ; Matthews et al., 1997). Detelles réponses peuvent être employées pour étudier lamotivation uniquement lorsque l’apprentissage est asymp-
totique car, dans le cas contraire, l’effet de la manipulationexpérimentale pourrait concerner autant l’apprentissageque la motivation. Et, même dans ce cas, les effets desvariables expérimentales sur la motivation peuvent facile-ment être confondus avec les effets sur la mémoire. Le ratcourt-il plus lentement sur la piste parce que sa motivationa été réduite ou parce qu’il a oublié qu’il y avait unefemelle réceptive dans la boîte à atteindre ? La réponse àcette question pourrait peut-être être obtenue par quel-ques expériences de contrôle mais il peut également êtretrès difficile d’éliminer l’explication mnésique. Cette ambi-guïté peut être évitée par l’utilisation d’une réponse nedépendant pas d’un apprentissage.
Un autre problème, sans doute bien plus sérieux encore,vient du fait que ces procédures infèrent l’état motivation-nel à partir du taux ou de la vitesse de la réponse opérante.Or, les fonctions motrices sont susceptibles d’être affectéesdès que l’on utilise des manipulations expérimentales, enparticulier avec des traitements pharmacologiques et, afortiori, avec de grandes doses. Les effets moteurs se trou-vent alors confondus avec ceux sur la motivation. La confu-sion est encore plus importante lorsque les procédures uti-lisent les activités locomotrices conditionnées commeindicateurs de la motivation sexuelle (Mendelson et Pfaus,1989 ; Pfaus et al., 1999). Pour éviter ce biais, il est doncessentiel d’avoir une mesure la plus indépendante possibledes fonctions motrices.
Un test de la motivation sexuelle chez le ratpouvant servir de modèle préclinique
Mesure de la motivation sexuelleLa mesure de la motivation sexuelle, obtenue par ce pro-cédé, est le temps passé dans le secteur à côté d’un incita-teur sexuel inaccessible comparé au temps passé dans lesecteur à côté d’un incitateur social et non sexuel égale-ment inaccessible (Fig. 1). Ainsi, plutôt que de mesurer lavitesse ou le taux de réponse, le procédé utilise la perma-nence dans un secteur particulier comme indicateur demotivation. Les capacités motrices nécessaires pour resterdans un endroit particulier de la cage ne sont pas exigean-tes mais le sujet doit montrer un certain niveau d’activitéambulatoire pour atteindre et explorer chacun des sec-teurs. Par conséquent, les variables telles que la distancede déplacement pendant le test, la vitesse des déplace-ments et le temps d’immobilité sont toujours enregistrés.Au cas où le traitement expérimental aurait des effets surl’activité motrice, il sera nécessaire d’y accorder uneattention particulière pour interpréter les résultats. Alter-nativement, les indicateurs de l’activité ambulatoire peu-vent être employés comme covariants dans les analyses,réduisant ou éliminant la confusion provoquée par deseffets sur le fonctionnement moteur. Le fait que l’activitéambulatoire puisse être évaluée simultanément mais indé-pendamment des paramètres de motivation est un avantageimportant du procédé.
Un rat peut s’approcher d’un autre rat pour de nombreu-ses raisons. Il peut, par exemple, avoir froid et vouloir seréchauffer, chercher une protection contre un prédateur,chercher le contact social ou des rapports sexuels. En labo-ratoire, la température est telle que les rats n’ont pas

Figure 1 Photo du dispositif utilisé pour le test de motivationsexuelle. Les boîtes des animaux incitateurs (marqués avec unA sur la photo) se trouvent à l’extérieur d’un compartimentouvert en forme d’arène (100 * 50 cm). Elles sont facilementdémontables permettant ainsi de changer la position des inci-tateurs aléatoirement entre chaque test. Le côté adjacent à cecompartiment ouvert est constitué d’un grillage qui permet ausujet expérimental de voir, de sentir et d’entendre l’incitateur.Une zone virtuelle de 21 * 29 cm (marquée avec un B sur laphoto) est définie devant chaque boîte d’incitateur. Un sys-tème de videotrack détermine la position du sujet expérimen-tal. Il calcule le temps passé et le nombre de visites dans leszones virtuelles ; la distance parcourue, la vitesse de déplace-ment et le temps d’immobilité durant le test.A photograph of the sexual incentive motivation test arena.The incentive animal cages (marked with an A on the photo-graph) are located on the outside of the oval arena (100 *50 cm). They are detachable from the outside of the wall sothat the position of the incentive animals can be changed ran-domly. The side facing the arena is made of a wire mesh thatallows the experimental subject to see, smell and hear theincentives. A virtual zone of 21 * 29 cm (marked with a B onthe photograph) is defined outside each incentive animalcage. A computerized videotrack system determines the expe-rimental subject’s position, the time spent in the incentivezones, the number of visits to them, the distance moved duringthe test, the mean speed of movement while moving, and theimmobility time.
Modèles précliniques du désir sexuel 245
besoin de se réchauffer, de fuir un prédateur, etc. Dansnotre dispositif, nous supposons donc que les causes princi-pales de l’approche entre animaux sont, soit la recherchedu contact social, soit celle de rapports sexuels. Le rat estun animal grégaire qui aime bien se retrouver proche de sescongénères (Latané, 1969 ; Latané et Glass, 1968 ; Sloan etLatané, 1974) et ils s’engagent volontiers dans des compor-tements sexuels lorsqu’ils trouvent un partenaire adéquat.Pour ces raisons, nous proposons toujours deux incitateursau sujet expérimental : un incitateur social (rat ne mon-trant aucune activité sexuelle lorsqu’il en a l’occasion) etun incitateur sexuel (rat montrant une activité sexuellelorsque l’occasion se présente). Ce dernier est, en réalité,un double incitateur car, en plus d’être un possible parte-naire sexuel, il (ou elle) est aussi un incitateur social. La
différence entre l’approche de l’incitateur social et cellede l’incitateur sexuel et social constitue une mesure del’intensité de la motivation sexuelle.
Lorsque le sujet expérimental est une femelle, l’incita-teur social et sexuel est un mâle intact sexuellement actifet l’incitateur social est un mâle castré. Lorsqu’un mâle estle sujet d’expérience, l’incitateur social est un autre mâleet une femelle sexuellement réceptive est l’incitateursocial et sexuel. Dans ce contexte, il faut remarquer queles animaux incitateurs n’ont aucune propriété aversive.Curieusement, certains chercheurs du monde anglo-saxons’imaginent que l’interaction entre les rats est toujoursinfluencée ou carrément déterminée par les comporte-ments de peur ou d’agressivité. Cela est loin de la réalité.Comme nous l’avons déjà mentionné, les rats sont des ani-maux grégaires. Si on laisse un sujet mâle choisir entre uncompartiment vide et un compartiment contenant un autremâle, il se trouve qu’il préfère largement passer son tempsprès du compartiment du mâle plutôt qu’à côté du compar-timent vide (Ågmo et al., 2004). Comme prévu, le mâlefonctionne comme incitateur social sans activer de compor-tement d’évitement. On obtient des résultats identiqueschez la femelle, c’est-à-dire, qu’elles préfèrent toujoursun compartiment avec un autre rat à un compartimentvide. En conséquence, il semble tout à fait raisonnable deparler d’incitateur social aussitôt qu’on parle d’un rat sansactivité sexuelle.
La validation expérimentale
Un autre animal peut fonctionner comme incitateur sexueluniquement si l’animal de l’expérience cherche le contactsexuel. Il est bien connu que les femelles ovariectomiséesne montreront aucun comportement sexuel et qu’un mâlecastré ne montrera, de même, aucun comportement copu-latoire à condition que l’on attende un certain temps aprèsla castration. Donc, une femelle ovariectomisée ou un mâlecastré ne montreront jamais de motivation due à une incita-tion sexuelle. Au contraire, la motivation due à l’incitationsociale ne devrait pas changer quand les hormones gonadi-ques sont absentes. En conséquence, un mâle castré ou unefemelle ovariectomisée devrait passer un temps à peu prèsidentique à proximité des incitateurs sexuels et sociaux.
Comme nous pouvons le voir sur les Fig. 2A et B, letemps que les mâles ont passé à proximité d’une femelleréceptive est réduit après la castration alors que le tempspassé à proximité d’un autre mâle n’est pas modifié. Deplus, l’intérêt pour la femelle réceptive regagne le niveauprécédant la castration après l’administration de testosté-rone. Nos prévisions s’avèrent donc exactes puisque laréponse expérimentale du sujet à l’incitation sexuelle estmodifiée dans le sens envisagé et que celle à l’incitationsociale ne change pas.
Pour pousser plus loin la validation du procédé, nousavons réalisé un test de motivation dans lequel les mâlesont eu libre accès aux femelles réceptives durant quatreheures. Cette activité sexuelle prolongée juste avant letest devrait réduire la motivation sexuelle. La Fig. 3 nousmontre que la motivation sexuelle a bien été réduitecomme nous l’avions prévu.
De même chez les femelles, la préférence pour le mâlesexuellement actif par rapport à celui qui est castré est
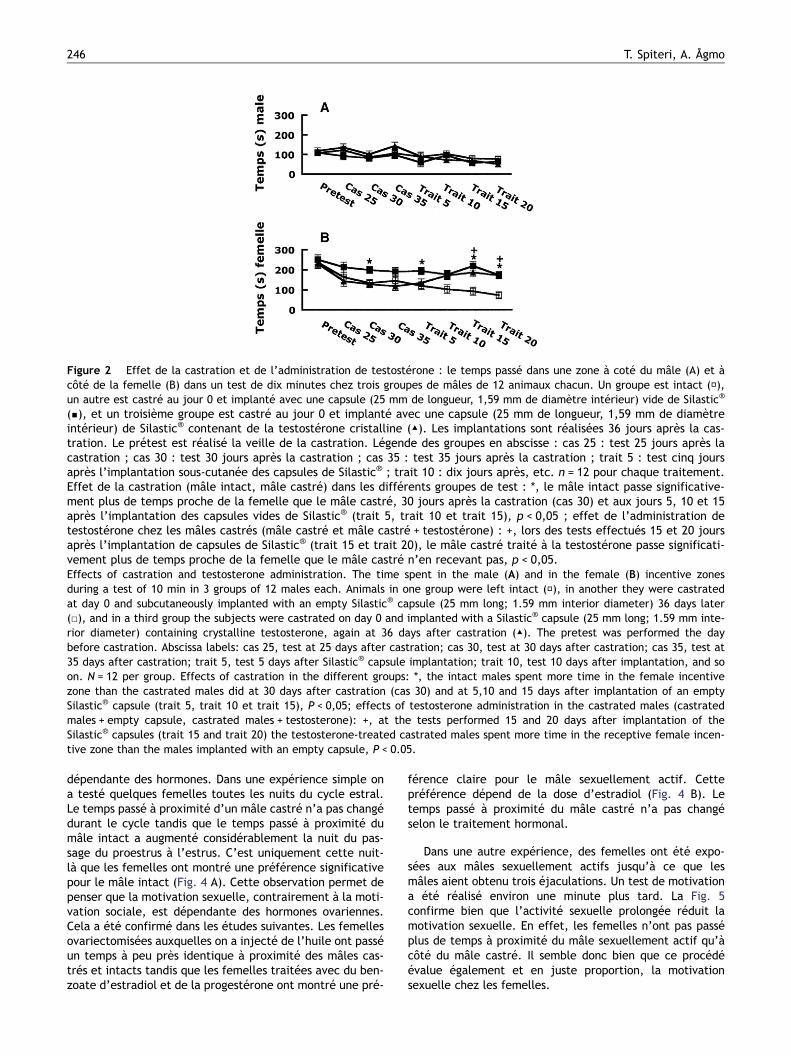
Figure 2 Effet de la castration et de l’administration de testostérone : le temps passé dans une zone à coté du mâle (A) et àcôté de la femelle (B) dans un test de dix minutes chez trois groupes de mâles de 12 animaux chacun. Un groupe est intact (■),un autre est castré au jour 0 et implanté avec une capsule (25 mm de longueur, 1,59 mm de diamètre intérieur) vide de Silastic®
(▪), et un troisième groupe est castré au jour 0 et implanté avec une capsule (25 mm de longueur, 1,59 mm de diamètreintérieur) de Silastic® contenant de la testostérone cristalline (▲). Les implantations sont réalisées 36 jours après la cas-tration. Le prétest est réalisé la veille de la castration. Légende des groupes en abscisse : cas 25 : test 25 jours après lacastration ; cas 30 : test 30 jours après la castration ; cas 35 : test 35 jours après la castration ; trait 5 : test cinq joursaprès l’implantation sous-cutanée des capsules de Silastic® ; trait 10 : dix jours après, etc. n = 12 pour chaque traitement.Effet de la castration (mâle intact, mâle castré) dans les différents groupes de test : *, le mâle intact passe significative-ment plus de temps proche de la femelle que le mâle castré, 30 jours après la castration (cas 30) et aux jours 5, 10 et 15après l’implantation des capsules vides de Silastic® (trait 5, trait 10 et trait 15), p < 0,05 ; effet de l’administration detestostérone chez les mâles castrés (mâle castré et mâle castré + testostérone) : +, lors des tests effectués 15 et 20 joursaprès l’implantation de capsules de Silastic® (trait 15 et trait 20), le mâle castré traité à la testostérone passe significati-vement plus de temps proche de la femelle que le mâle castré n’en recevant pas, p < 0,05.Effects of castration and testosterone administration. The time spent in the male (A) and in the female (B) incentive zonesduring a test of 10 min in 3 groups of 12 males each. Animals in one group were left intact (■), in another they were castratedat day 0 and subcutaneously implanted with an empty Silastic® capsule (25 mm long; 1.59 mm interior diameter) 36 days later(□), and in a third group the subjects were castrated on day 0 and implanted with a Silastic® capsule (25 mm long; 1.59 mm inte-rior diameter) containing crystalline testosterone, again at 36 days after castration (▲). The pretest was performed the daybefore castration. Abscissa labels: cas 25, test at 25 days after castration; cas 30, test at 30 days after castration; cas 35, test at35 days after castration; trait 5, test 5 days after Silastic® capsule implantation; trait 10, test 10 days after implantation, and soon. N = 12 per group. Effects of castration in the different groups: *, the intact males spent more time in the female incentivezone than the castrated males did at 30 days after castration (cas 30) and at 5,10 and 15 days after implantation of an emptySilastic® capsule (trait 5, trait 10 et trait 15), P < 0,05; effects of testosterone administration in the castrated males (castratedmales + empty capsule, castrated males + testosterone): +, at the tests performed 15 and 20 days after implantation of theSilastic® capsules (trait 15 and trait 20) the testosterone-treated castrated males spent more time in the receptive female incen-tive zone than the males implanted with an empty capsule, P < 0.05.
T. Spiteri, A. Ågmo246
dépendante des hormones. Dans une expérience simple ona testé quelques femelles toutes les nuits du cycle estral.Le temps passé à proximité d’un mâle castré n’a pas changédurant le cycle tandis que le temps passé à proximité dumâle intact a augmenté considérablement la nuit du pas-sage du proestrus à l’estrus. C’est uniquement cette nuit-là que les femelles ont montré une préférence significativepour le mâle intact (Fig. 4 A). Cette observation permet depenser que la motivation sexuelle, contrairement à la moti-vation sociale, est dépendante des hormones ovariennes.Cela a été confirmé dans les études suivantes. Les femellesovariectomisées auxquelles on a injecté de l’huile ont passéun temps à peu près identique à proximité des mâles cas-trés et intacts tandis que les femelles traitées avec du ben-zoate d’estradiol et de la progestérone ont montré une pré-
férence claire pour le mâle sexuellement actif. Cettepréférence dépend de la dose d’estradiol (Fig. 4 B). Letemps passé à proximité du mâle castré n’a pas changéselon le traitement hormonal.
Dans une autre expérience, des femelles ont été expo-sées aux mâles sexuellement actifs jusqu’à ce que lesmâles aient obtenu trois éjaculations. Un test de motivationa été réalisé environ une minute plus tard. La Fig. 5confirme bien que l’activité sexuelle prolongée réduit lamotivation sexuelle. En effet, les femelles n’ont pas passéplus de temps à proximité du mâle sexuellement actif qu’àcôté du mâle castré. Il semble donc bien que ce procédéévalue également et en juste proportion, la motivationsexuelle chez les femelles.
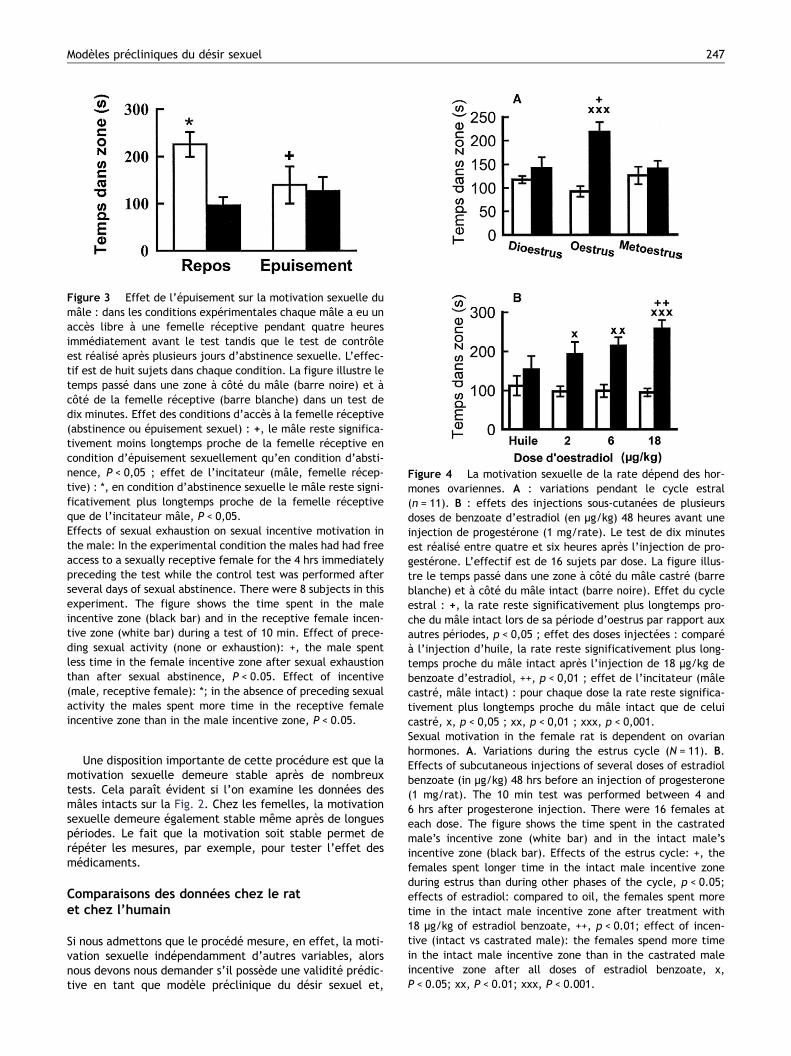
Figure 3 Effet de l’épuisement sur la motivation sexuelle dumâle : dans les conditions expérimentales chaque mâle a eu unaccès libre à une femelle réceptive pendant quatre heuresimmédiatement avant le test tandis que le test de contrôleest réalisé après plusieurs jours d’abstinence sexuelle. L’effec-tif est de huit sujets dans chaque condition. La figure illustre letemps passé dans une zone à côté du mâle (barre noire) et àcôté de la femelle réceptive (barre blanche) dans un test dedix minutes. Effet des conditions d’accès à la femelle réceptive(abstinence ou épuisement sexuel) : +, le mâle reste significa-tivement moins longtemps proche de la femelle réceptive encondition d’épuisement sexuellement qu’en condition d’absti-nence, P < 0,05 ; effet de l’incitateur (mâle, femelle récep-tive) : *, en condition d’abstinence sexuelle le mâle reste signi-ficativement plus longtemps proche de la femelle réceptiveque de l’incitateur mâle, P < 0,05.Effects of sexual exhaustion on sexual incentive motivation inthe male: In the experimental condition the males had had freeaccess to a sexually receptive female for the 4 hrs immediatelypreceding the test while the control test was performed afterseveral days of sexual abstinence. There were 8 subjects in thisexperiment. The figure shows the time spent in the maleincentive zone (black bar) and in the receptive female incen-tive zone (white bar) during a test of 10 min. Effect of prece-ding sexual activity (none or exhaustion): +, the male spentless time in the female incentive zone after sexual exhaustionthan after sexual abstinence, P < 0.05. Effect of incentive(male, receptive female): *; in the absence of preceding sexualactivity the males spent more time in the receptive femaleincentive zone than in the male incentive zone, P < 0.05.
Figure 4 La motivation sexuelle de la rate dépend des hor-mones ovariennes. A : variations pendant le cycle estral(n = 11). B : effets des injections sous-cutanées de plusieursdoses de benzoate d’estradiol (en μg/kg) 48 heures avant uneinjection de progestérone (1 mg/rate). Le test de dix minutesest réalisé entre quatre et six heures après l’injection de pro-gestérone. L’effectif est de 16 sujets par dose. La figure illus-tre le temps passé dans une zone à côté du mâle castré (barreblanche) et à côté du mâle intact (barre noire). Effet du cycleestral : +, la rate reste significativement plus longtemps pro-che du mâle intact lors de sa période d’oestrus par rapport auxautres périodes, p < 0,05 ; effet des doses injectées : comparéà l’injection d’huile, la rate reste significativement plus long-temps proche du mâle intact après l’injection de 18 μg/kg debenzoate d’estradiol, ++, p < 0,01 ; effet de l’incitateur (mâlecastré, mâle intact) : pour chaque dose la rate reste significa-tivement plus longtemps proche du mâle intact que de celuicastré, x, p < 0,05 ; xx, p < 0,01 ; xxx, p < 0,001.Sexual motivation in the female rat is dependent on ovarianhormones. A. Variations during the estrus cycle (N = 11). B.Effects of subcutaneous injections of several doses of estradiolbenzoate (in μg/kg) 48 hrs before an injection of progesterone(1 mg/rat). The 10 min test was performed between 4 and6 hrs after progesterone injection. There were 16 females ateach dose. The figure shows the time spent in the castratedmale’s incentive zone (white bar) and in the intact male’sincentive zone (black bar). Effects of the estrus cycle: +, thefemales spent longer time in the intact male incentive zoneduring estrus than during other phases of the cycle, p < 0.05;effects of estradiol: compared to oil, the females spent moretime in the intact male incentive zone after treatment with18 μg/kg of estradiol benzoate, ++, p < 0.01; effect of incen-tive (intact vs castrated male): the females spend more timein the intact male incentive zone than in the castrated maleincentive zone after all doses of estradiol benzoate, x,P < 0.05; xx, P < 0.01; xxx, P < 0.001.
Modèles précliniques du désir sexuel 247
Une disposition importante de cette procédure est que lamotivation sexuelle demeure stable après de nombreuxtests. Cela paraît évident si l’on examine les données desmâles intacts sur la Fig. 2. Chez les femelles, la motivationsexuelle demeure également stable même après de longuespériodes. Le fait que la motivation soit stable permet derépéter les mesures, par exemple, pour tester l’effet desmédicaments.
Comparaisons des données chez le ratet chez l’humain
Si nous admettons que le procédé mesure, en effet, la moti-vation sexuelle indépendamment d’autres variables, alorsnous devons nous demander s’il possède une validité prédic-tive en tant que modèle préclinique du désir sexuel et,
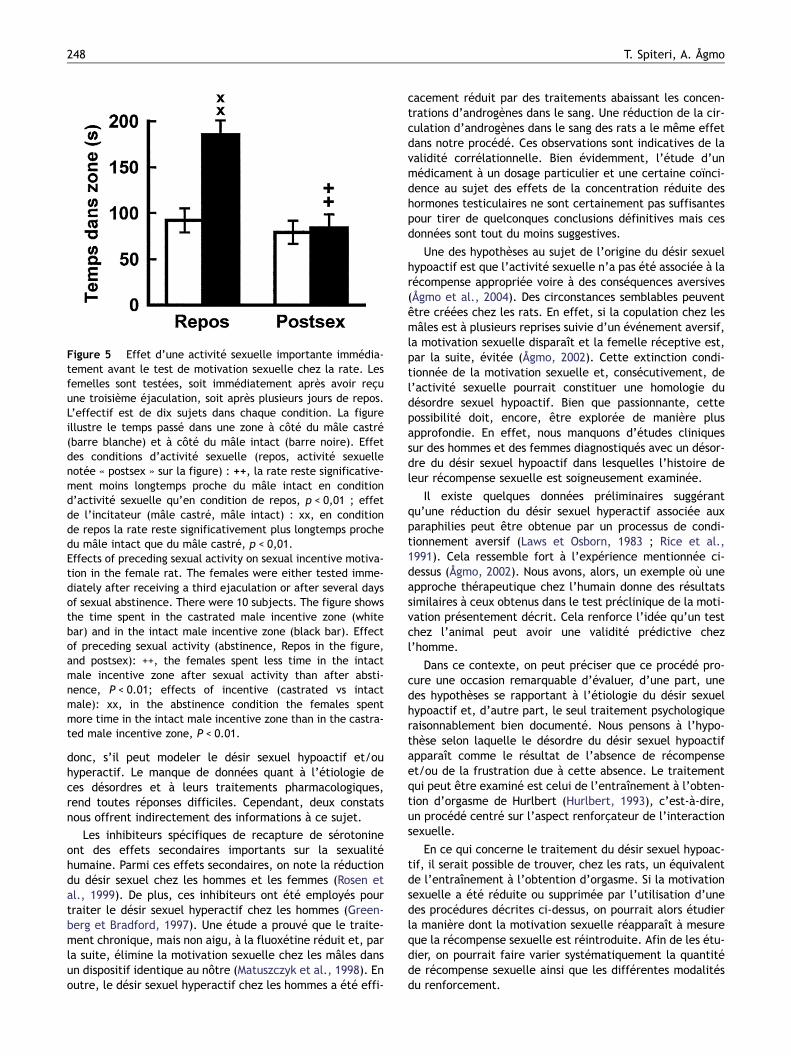
Figure 5 Effet d’une activité sexuelle importante immédia-tement avant le test de motivation sexuelle chez la rate. Lesfemelles sont testées, soit immédiatement après avoir reçuune troisième éjaculation, soit après plusieurs jours de repos.L’effectif est de dix sujets dans chaque condition. La figureillustre le temps passé dans une zone à côté du mâle castré(barre blanche) et à côté du mâle intact (barre noire). Effetdes conditions d’activité sexuelle (repos, activité sexuellenotée « postsex » sur la figure) : ++, la rate reste significative-ment moins longtemps proche du mâle intact en conditiond’activité sexuelle qu’en condition de repos, p < 0,01 ; effetde l’incitateur (mâle castré, mâle intact) : xx, en conditionde repos la rate reste significativement plus longtemps prochedu mâle intact que du mâle castré, p < 0,01.Effects of preceding sexual activity on sexual incentive motiva-tion in the female rat. The females were either tested imme-diately after receiving a third ejaculation or after several daysof sexual abstinence. There were 10 subjects. The figure showsthe time spent in the castrated male incentive zone (whitebar) and in the intact male incentive zone (black bar). Effectof preceding sexual activity (abstinence, Repos in the figure,and postsex): ++, the females spent less time in the intactmale incentive zone after sexual activity than after absti-nence, P < 0.01; effects of incentive (castrated vs intactmale): xx, in the abstinence condition the females spentmore time in the intact male incentive zone than in the castra-ted male incentive zone, P < 0.01.
T. Spiteri, A. Ågmo248
donc, s’il peut modeler le désir sexuel hypoactif et/ouhyperactif. Le manque de données quant à l’étiologie deces désordres et à leurs traitements pharmacologiques,rend toutes réponses difficiles. Cependant, deux constatsnous offrent indirectement des informations à ce sujet.
Les inhibiteurs spécifiques de recapture de sérotonineont des effets secondaires importants sur la sexualitéhumaine. Parmi ces effets secondaires, on note la réductiondu désir sexuel chez les hommes et les femmes (Rosen etal., 1999). De plus, ces inhibiteurs ont été employés pourtraiter le désir sexuel hyperactif chez les hommes (Green-berg et Bradford, 1997). Une étude a prouvé que le traite-ment chronique, mais non aigu, à la fluoxétine réduit et, parla suite, élimine la motivation sexuelle chez les mâles dansun dispositif identique au nôtre (Matuszczyk et al., 1998). Enoutre, le désir sexuel hyperactif chez les hommes a été effi-
cacement réduit par des traitements abaissant les concen-trations d’androgènes dans le sang. Une réduction de la cir-culation d’androgènes dans le sang des rats a le même effetdans notre procédé. Ces observations sont indicatives de lavalidité corrélationnelle. Bien évidemment, l’étude d’unmédicament à un dosage particulier et une certaine coïnci-dence au sujet des effets de la concentration réduite deshormones testiculaires ne sont certainement pas suffisantespour tirer de quelconques conclusions définitives mais cesdonnées sont tout du moins suggestives.
Une des hypothèses au sujet de l’origine du désir sexuelhypoactif est que l’activité sexuelle n’a pas été associée à larécompense appropriée voire à des conséquences aversives(Ågmo et al., 2004). Des circonstances semblables peuventêtre créées chez les rats. En effet, si la copulation chez lesmâles est à plusieurs reprises suivie d’un événement aversif,la motivation sexuelle disparaît et la femelle réceptive est,par la suite, évitée (Ågmo, 2002). Cette extinction condi-tionnée de la motivation sexuelle et, consécutivement, del’activité sexuelle pourrait constituer une homologie dudésordre sexuel hypoactif. Bien que passionnante, cettepossibilité doit, encore, être explorée de manière plusapprofondie. En effet, nous manquons d’études cliniquessur des hommes et des femmes diagnostiqués avec un désor-dre du désir sexuel hypoactif dans lesquelles l’histoire deleur récompense sexuelle est soigneusement examinée.
Il existe quelques données préliminaires suggérantqu’une réduction du désir sexuel hyperactif associée auxparaphilies peut être obtenue par un processus de condi-tionnement aversif (Laws et Osborn, 1983 ; Rice et al.,1991). Cela ressemble fort à l’expérience mentionnée ci-dessus (Ågmo, 2002). Nous avons, alors, un exemple où uneapproche thérapeutique chez l’humain donne des résultatssimilaires à ceux obtenus dans le test préclinique de la moti-vation présentement décrit. Cela renforce l’idée qu’un testchez l’animal peut avoir une validité prédictive chezl’homme.
Dans ce contexte, on peut préciser que ce procédé pro-cure une occasion remarquable d’évaluer, d’une part, unedes hypothèses se rapportant à l’étiologie du désir sexuelhypoactif et, d’autre part, le seul traitement psychologiqueraisonnablement bien documenté. Nous pensons à l’hypo-thèse selon laquelle le désordre du désir sexuel hypoactifapparaît comme le résultat de l’absence de récompenseet/ou de la frustration due à cette absence. Le traitementqui peut être examiné est celui de l’entraînement à l’obten-tion d’orgasme de Hurlbert (Hurlbert, 1993), c’est-à-dire,un procédé centré sur l’aspect renforçateur de l’interactionsexuelle.
En ce qui concerne le traitement du désir sexuel hypoac-tif, il serait possible de trouver, chez les rats, un équivalentde l’entraînement à l’obtention d’orgasme. Si la motivationsexuelle a été réduite ou supprimée par l’utilisation d’unedes procédures décrites ci-dessus, on pourrait alors étudierla manière dont la motivation sexuelle réapparaît à mesureque la récompense sexuelle est réintroduite. Afin de les étu-dier, on pourrait faire varier systématiquement la quantitéde récompense sexuelle ainsi que les différentes modalitésdu renforcement.

Be
Ell
Fug
Gre
Hu
Lat
Lat
Lau
Law
Lóp
Ma
Ma
Me
Me
Pfa
Pfa
Pfa
Ric
Ros
Slo
Wh
Wh
Modèles précliniques du désir sexuel 249
Conclusion
Les études épidémiologiques ont prouvé que les désordres dudésir sexuel sont communs, en particulier chez les femmes.Les traitements psychologiques prennent du temps et lesrésultats sont incertains à court et moyen terme. En ce quiconcerne leurs résultats à long terme nous ne savons prati-quement rien. D’ailleurs, la plupart des traitements psycho-logiques exigent non seulement la participation du patientmais aussi de son partenaire, et leur volonté d’investir dutemps et des efforts dans un traitement ayant des résultatsincertains n’est pas toujours présente. Ces problèmes sontencore plus importants pour les patients qui ne vivent pasen couple stable. Qui pourrait être alors, le partenaire dansl’entraînement à l’obtention d’orgasme, seul traitement surlequel on a fait des études plus ou moins acceptables d’effi-cacité ? Par conséquent, il est très probable que de nom-breux individus préfèrent un traitement pharmacologiquefiable et sans effet secondaire. Le développement systéma-tique de tels traitements exige assurément un bon modèlepréclinique. La recherche d’un traitement pharmacologiqueefficace tirerait un bénéfice considérable d’une meilleureconnaissance de l’étiologie des désordres du désir sexuel etun modèle animal qui pourrait contribuer à cette connais-sance serait non seulement souhaitable mais très utile.Dans le meilleur des cas, le modèle serait pertinent pourles mâles et pour les femelles aussi bien pour le faible quepour le fort désir sexuel. Nous avons présenté des donnéesproposant que le test de motivation sexuelle incitative déve-loppé à l’origine par Meyerson et Lindström (1973) et, plustard, adapté par Ågmo et al. (Ågmo, 2003 ; Ellingsen etÅgmo, 2004) puisse représenter un modèle acceptable.Notons qu’une évaluation finale de ce procédé n’est pas pos-sible actuellement en raison du manque de traitements phar-macologiques cliniquement efficaces.
L’importance de l’apprentissage dans l’étiologie desdésordres du désir sexuel n’est pas connue. Nous avons pro-posé que l’apprentissage puisse jouer un rôle central. Enconséquence, un bon modèle préclinique devrait pouvoir dis-tinguer les effets de divers apprentissages sur la motivationsexuelle. Le modèle que nous avons présenté à l’avantage depouvoir le faire.
Références
Ågmo A. Sexual motivation. An inquiry into events determining theoccurrence of sexual behavior. Behav Brain Res 1999; 105: 129-50.
Ågmo A. Copulation-contingent aversive conditioning and sexualincentive motivation in male rats: evidence for a two-stage pro-cess of sexual behavior. Physiol Behav 2002; 77: 425-35.
Ågmo A. Unconditioned sexual incentive motivation in the male Nor-way rat (Rattus norvegicus). J Comp Psychol 2003; 117: 3-14.
Ågmo A, Ellingsen E. Relevance of non-human animal studies to theunderstanding of human sexuality. Scand J Psychol 2003; 44: 291-9.
Ågmo A, Turi AL, Ellingsen E, Kaspersen H. Preclinical models ofsexual desire: Conceptual and behavioral analyses. PharmacolBiochem Behav 2004; 78: 379-404.
Arnal A, Llario R, Gil MD. Estudio epidemiológico sobre la prevalen-cia de disfunciones sexuales en la comunidad valenciana. Psico-thema 1995; 7: 95-104.
ach FA. A review of physiological and psychological studies ofsexual behavior in mammals. Physiol. Rev. 1947a;27:240–307.Beach FA. Evolutionary changes in the physiological control ofmating behavior in mammals. Psychol. Rev. 1947b;54:297–315.ingsen E, Ågmo A. Sexual incentive motivation and paced sexualbehavior in female rats after treatment with drugs modifyingdopaminergic neurotransmission. Pharmacol Biochem Behav2004; 77: 431-45.l-Meyer AR, Sjögren-Fugl-Meyer K. Sexual disabilities, problemsand satisfaction in 18–74 year old Swedes. Scand J Sexol 1999;2: 79-105.enberg DM, Bradford JMW. Treatment of the paraphilic disorders:a review of the role of the selective serotonin reuptake inhibi-tors. Sexual Abuse: J Treat Res 1997; 9: 349-60.rlbert DF. A comparative study using orgasm consistency trainingin the treatment of women reporting hypoactive sexual desire. JSex Marital Ther 1993; 19: 41-55.ané B. Gregariousness and fear in laboratory rats. J Exp Soc Psy-chol 1969; 5: 61-9.ané B, Glass DC. Social and nonsocial attraction in rats. J Pers SocPsychol 1968; 9: 142-6.mann EO, Paik A, Rosen RC. Sexual dysfunction in the United Sta-tes: prevalence and predictors. JAMA 1999; 281: 537-44.s DR, Osborn CA. How to build and operate a behavioral labora-tory to evaluate and treat sexual deviance. In: Greer JG,Stuart IR, eds. The sexual aggressor: current perspectives ontreatment. New York: Van Nostrand Reinhold, 1983: 293-335.ez HH, Olster DH, Ettenberg A. Sexual motivation in the malerat: the role of primary incentives and copulatory experience.Horm Behav 1999; 38: 176-85.tthews TJ, Grigore M, Tang L, Doat M, Kow LM, Pfaff DW. Sexualreinforcement in the female rat. J Exp Anal Behav 1997; 68: 399-410.tuszczyk J, Larsson K, Erikson E. The selective serotonin reuptakeinhibitor fluoxetine reduces sexual motivation in male rats. Phar-macol Biochem Behav 1998; 60: 527-32.ndelson SD, Pfaus JG. Level searching: a new assay of sexualmotivation in the male rat. Physiol Behav 1989; 45: 337-41.yerson BJ, Lindström LH. Sexual motivation in the female rat. Amethodological study applied to the investigation of the effect ofestradiol benzoate. Acta Physiol Scand 1973(Suppl. 389): 1-80.ff DW. Drive. Neurobiological and molecular mechanisms ofsexual motivation. Cambridge, MA: MIT Press, 1999.ff DW, Ågmo A. Reproductive motivation. In: Pashler H, GallistelR, eds. Steven’s handbook of experimental psychology. Volume 3,learning, motivation and emotion. New York: Wiley, 2002: 709-36.us JG, Smith WJ, Coopersmith CB. Appetitive and consummatorysexual behaviors of female rats in bilevel chambers. I. A correla-tional and factor analysis and the effects of ovarian hormones.Horm Behav 1999; 35: 224-40.e ME, Harris GT, Quinsey VL. Evaluation of an institution-basedtreatment program for child molesters. Can J Prog Eval 1991; 6:111-29.en RC, Lane RM, Menza M. Effects of SSRIs on sexual function: acritical review. J Clin Psychopharmacol 1999; 19: 67-85.an L, Latané B. Sex and sociability in rats. J Exp Soc Psychol1974; 10: 147-58.alen RE. Hormone-induced changes in the organization of sexualbehavior in the male rat. J Comp Physiol Psychol 1964; 57: 175-82.alen RE, Beach FA, Kuehn RE. Effects of exogenous androgen onsexually responsive and unresponsive male rats. Endocrinology1961; 68: 373-80.