Embed Size (px)
Citation preview

PDFlib PLOP: PDF Linearization, Optimization, Protection
Page inserted by evaluation versionwww.pdflib.com – [email protected]

93
Physiological Importance of the T3 Mitochondrial Pathway
CHANTAL WRUTNIAK, PIERRICK ROCHARD, FRANÇOIS CASAS,ANDRÉ FRAYSSE, JACQUES CHARRIER, AND GÉRARD CABELLO
Laboratoire de Différenciation Cellulaire et Croissance, Unité d'Endocrinologie Cellulaire, INRA-ENSA, 2 Place Viala, 34060 Montpellier Cedex, France
Numerous data have provided strong evidence that mitochondrial activity is hor-monally regulated. In particular, the influences of thyroid hormone,1
glucocorticoids2 and vitamin D33 have been reported. In addition, peroxisomes pro-liferators such as clofibrate, acting via the PPAR (peroxisome proliferator activatedreceptor), also stimulate mitochondrial activity.4 These results clearly suggest thatnuclear receptors belonging to the steroid receptor superfamily are important for theregulation of mitochondrial activity. The influence of triiodothyronine (T3) has beenparticularly studied. In this field, the initial proposition by Sterling’s group5 of a di-rect T3 mitochondrial pathway involving specific receptors has raised a controversyin which scientific objectivity was not always the rule. After a brief description ofhow T3 influences mitochondrial function, this review will focus on the recent dis-covery of truncated c-Erb Aα1 receptors in the organelle. This has led to new in-sights in the knowledge of mitochondrial regulation.
THE INFLUENCE OF T3 UPON MITOCHONDRIAL ACTIVITY
The influence of T3 upon mitochondrial activity is well documented. The differ-ent effects can be classified into three groups in relation to their latency period.1 Thework of Sterling’s group pointed out a very rapid stimulation of oxidative phospho-rylations, mitochondrial O2 consumption and translocator activity occurring in lessthan 2 minutes in vitro or 15 minutes in vivo after T3 administration,6,7 referred toas “immediate” influences. Although these actions are well described, they have notyet received any convincing explanation.
Short-term influences have also been reported. These include a decrease in mito-chondrial membrane potential occurring within several hours of treatment. Thisprobably results from important changes in the area and in the phospholipid compo-sition of the inner membrane induced by T3, leading to an increase in proton perme-ability.8,9 In addition, an increase in the inner membrane cardiolipin contentstimulates cardiolipin-sensitive mitochondrial carriers such as translocase or AT-Pase and could activate organelle function.10
Another short-term influence is T3 stimulation of the mitochondrial genome tran-scription. While the maximal influence requires several hours, this action is initiatedafter a latency period of several minutes in isolated mitochondria.11 As T3 increasesthe steady-state level of mt-TFA mRNA,12 a constitutive mitochondrial transcrip-tion factor13 encoded by a nuclear gene, several investigators consider this action to

94 ANNALS NEW YORK ACADEMY OF SCIENCES
be induced at the nuclear level. However, this hypothesis is not fully supported bythe very short-term effect recorded in isolated mitochondria.11
Last, a general stimulation of mitochondriogenesis occurring several days afterthe onset of a T3 treatment is observed.14 Because mitochondriogenesis needs a co-ordinated rise in the expression of the mitochondrial genome and of nuclear genesencoding mitochondrial proteins, this influence probably involves both intramito-chondrial and nuclear T3 mechanisms.
Despite the controversy about the occurrence of a direct T3 mitochondrial path-way, it should be underlined that the “immediate” effects of T3 are not explain-able by mechanisms initiated at the nuclear level: in addition to their rapidity, theyoccur in isolated mitochondria and therefore without nuclear influence. In addition,they are not affected by protein synthesis inhibitors.7
Moreover, the short-term T3 influence on mitochondrial genome transcription,observed in isolated mitochondria,11 also raises the hypothesis of the occurrence ofa mitochondrial T3-dependent transcription factor. All these data are therefore inagreement with the possibility that a T3 receptor could be located in the mitochon-dria.
TRUNCATED FORMS OF A T3 NUCLEAR RECEPTOR ARELOCATED IN THE MITOCHONDRIA
Sterling's group provided the first evidence for the existence of mitochondrial T3receptors. The organelle was shown to be a major T3 accumulation compartment15
and specific T3 binding sites were described.5 This latter result was confirmed bytwo other groups,16,17 but other data concluded that the binding observed was notsaturable.18 Therefore, to date, the occurrence of T3 mitochondrial receptors re-mains controversial.
Recently, using a photoaffinity labeling technique, our laboratory identified twoT3 binding proteins in highly purified rat liver mitochondrial extracts. One was lo-cated in the matrix (MW 43 kDa) and the other, in minute amounts, in the innermembrane (MW 28 kDa).19 These data are in partial agreement with Sterling's work.Using two different antibodies raised against the T3 nuclear receptor c-Erb A, wedetected by Western blot two proteins (p43 and p28) displaying identical locationand molecular weights as T3 binding proteins (FIG. 1).19 In addition the T3-labeled43 kDa protein (p43) was immunoprecipitated by a c-Erb A antibody. Only c-Erb Aα forms were detected in mitochondria despite the fact that c-Erb Aβ was themajor nuclear receptor form in rat liver.20
Bigler et al.21 reported that truncated c-Erb Aα1 proteins are synthesized fromthe c-Erb A mRNA encoding the T3 nuclear receptor by using internal AUG codon.Transfection in CV1 cells of a c-Erb Aα1 construct expressing a major 43 kDaprotein21 demonstrated the mitochondrial location of this protein.19
We have identified five sequences of the rat mitochondrial genome, each highlyrelated to thyroid hormone response elements (T3RE) well-characterized in nuclearT3 target genes (TABLE 1). Interestingly, we reported that p43 specifically binds toone of these sequences, located in the D-loop, which contains the promoters of themitochondrial genome.19 Our present data also indicate that p43 binds to at least

95WRUTNIAK et al.: THE T3 MITOCHONDRIAL PATHWAY
three other sequences. These results raise the hypothesis that p43 could act as a T3-dependent mitochondrial transcription factor.
PHYSIOLOGICAL IMPORTANCE OF P43
Tissue-specific expression of p43 was studied in newborn rabbits.19 Intramito-chondrial amounts of this protein were positively correlated with the number of mi-tochondria in several tissues. In particular, brown adipose tissue, exhibiting a veryimportant number of organelles, displayed the highest amount of p43 in the mito-chondria. In addition, adult rat brain organelles were devoid of p43. In contrast tothe important responsiveness of brown adipose tissue,22 the brain is not sensitive tothe thermogenic T3 influence; this observation suggests that p43 could be involvedin this action.
Expression of p43 in CV1 cells led to a stimulation of mitochondrial activity asassessed by an increase in membrane potential and cytochrome oxidase activity.19
However, enzymatic complexes such as citrate synthase and malate dehydrogenase,which are encoded only by the nuclear genome, are less influenced (unpublished da-ta). These results underline a specific influence of p43 upon an enzymatic complexincluding subunits encoded by the mitochondrial genome, in agreement with a pos-sible function as a mitochondrial transcription factor.
More surprising was the observation that p43 overexpression in a myoblast avianline strongly stimulated myogenic differentiation (FIG. 2), thus suggesting an in-volvement of the mitochondrial T3 pathway in the regulation of cell differenti-ation. This hypothesis is well supported by works reporting that mitochondrialfunction affects terminal differentiation. In line with data obtained inerythrocytes23 and neurons,24 several reports indicate that mitochondrial functionstrongly influences myogenic differentiation. First, acute changes in mitochondrialactivity occur before the onset of myoblast differentiation and are not observed indifferentiation-deficient myoblasts.25 Secondly, expression of the v-erb A oncogeneinduces significant changes in mitochondrial activity25 before exhibiting its myo-
TABLE 1. Nucleotide Sequences in Rat Mitochondrial Genome
Nucleotide Sequence TRE Type Gene Position
AGGTCAAGGTGT DR0a ARN 12S 748-760
CGACCTatttaagAGTTCA Ipal7a ARN 16S 2374-2392
TGACCTaaaaccAGGTGA Ipal6 COX II 7367-7384
TGCCTTcctcaacatagccgtcAAGGCA TRE RSVa D-loop 15909-15936
AGGCATgaAGGTCA DR2a D-loop 15932-15945
NOTE: Several sequences similar to nuclear T3REs are located in the rat mitochondrialgenome. The number of the first and last nucleotide of each sequence is given according toGadaletta et al. 33
DR, direct repeat, Ipal, inverted palindrome; TRE RSV, TRE identified in the genome of theRous sarcoma virus.
aSequences tested in gel-retardation experiments showing specific binding of p43.
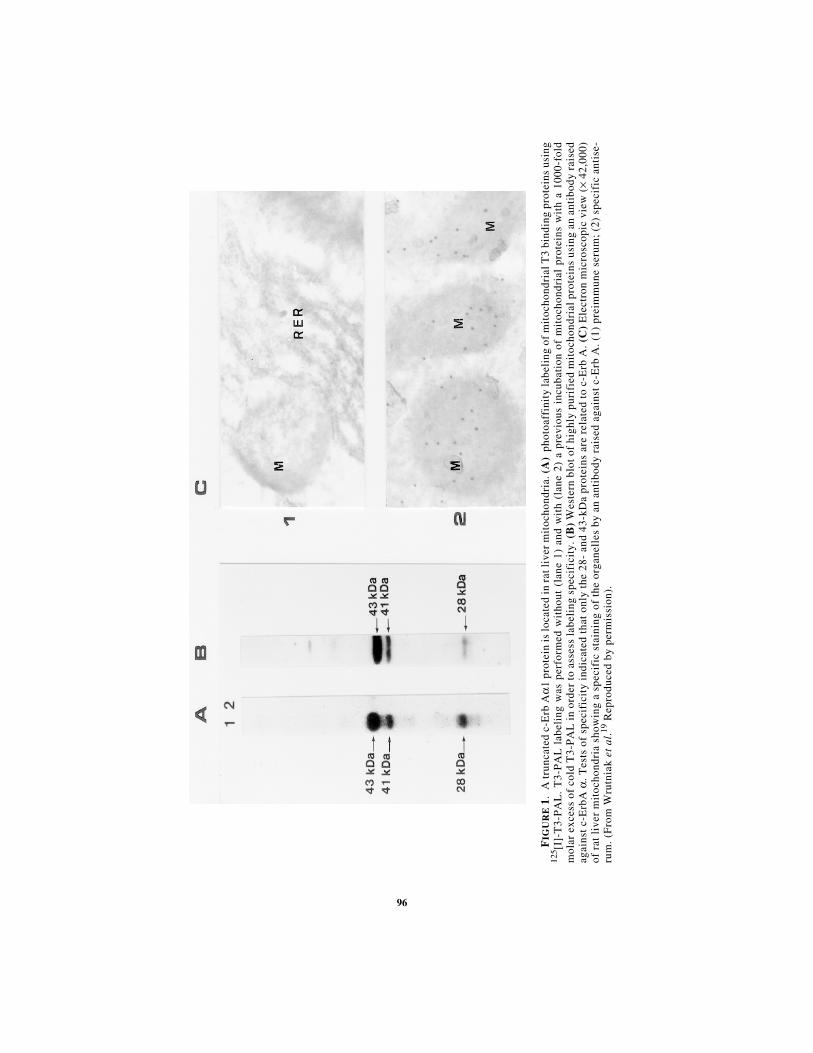
96
FIG
UR
E 1
.A
trun
cate
d c-
Erb
Aα
1 pr
otei
n is
loca
ted
in r
at li
ver
mit
ocho
ndri
a. (
A)
phot
oaff
init
y la
beli
ng o
f m
itoc
hond
rial
T3
bind
ing
prot
eins
usi
ng12
5 [I]-
T3-
PA
L.
T3-
PA
L l
abel
ing
was
per
form
ed w
itho
ut (
lane
1)
and
wit
h (l
ane
2) a
pre
viou
s in
cuba
tion
of
mit
ocho
ndri
al p
rote
ins
wit
h a
1000
-fol
dm
olar
exc
ess
of c
old
T3-
PA
L i
n or
der
to a
sses
s la
beli
ng s
peci
fici
ty. (
B)
Wes
tern
blo
t of
hig
hly
puri
fied
mit
ocho
ndri
al p
rote
ins
usin
g an
ant
ibod
y ra
ised
agai
nst
c-E
rbA
α. T
ests
of
spec
ific
ity
indi
cate
d th
at o
nly
the
28-
and
43-k
Da
prot
eins
are
rel
ated
to
c-E
rb A
. (C
) E
lect
ron
mic
rosc
opic
vie
w (
× 42
,000
)of
rat
liv
er m
itoc
hond
ria
show
ing
a sp
ecif
ic s
tain
ing
of t
he o
rgan
elle
s by
an
anti
body
rai
sed
agai
nst
c-E
rb A
. (1)
pre
imm
une
seru
m;
(2)
spec
ific
ant
ise-
rum
. (F
rom
Wru
tnia
k et
al .
19 R
epro
duce
d by
per
mis
sion
).

97WRUTNIAK et al.: THE T3 MITOCHONDRIAL PATHWAY
genic potency.26 Lastly, inhibition of mitochondrial protein synthesis by chloram-phenicol induces a strong differentiation block.27
THE MITOCHONDRIA: A DIRECT TARGET OF RECEPTORS RELATED TO THE STEROID RECEPTOR SUPERFAMILY?
Receptors of thyroid hormone, glucocorticoids, and vitamin D3 all belong to thesteroid receptor superfamily. In addition, peroxisome proliferators such as clofibratealso act via the PPAR, another member of this family. Strikingly, all these factorshave been shown to stimulate mitochondrial activity.1–4 Therefore, the identifica-tion of truncated forms of c-Erb A T3 receptors in the organelle, raises the questionof the presence of other nuclear receptors in the mitochondrion.
The presence of the native form of the nuclear glucocorticoid receptor in the or-ganelle has already been reported, as has the occurrence of glucocorticoid responseelements in the mitochondrial genome.28 Therefore, to date, two members of the ste-roid receptor superfamily display a dual location in the nucleus and in the mitochon-drion.
The observation that, in contrast to c-Erb Aα, the β form of the T3 nuclear recep-tor is not addressed in the mitochondrion19 could be helpful to understand the mito-chondrial import of these receptors. Taking into account the high homology betweenthe α and β proteins, an amino acid sequence located in the less conserved domains(N-terminal and hinge region) is probably involved in mitochondrial targeting. Iden-tification of this sequence will allow prediction of whether other nuclear receptorscould enter the mitochondrion.
CONCLUSIONS
The data presented in this review suggest that mitochondria could be an importanttarget of hormonal action. First, mitochondrial activity is under a well established
FIGURE 2. p43 overexpression stimulates avian myoblast differentiation. Cytoimmu-nofluorescence study (× 100). Staining by an antibody raised against connectin, a muscle-specific protein used as a marker of differentiation. (1) quail myoblasts stably transfectedwith an “empty vector”; (2) quail myoblasts transfected with a p43 expression vector. Thenumber of myotubes is strongly enhanced in p43-overexpressing myoblasts.

98 ANNALS NEW YORK ACADEMY OF SCIENCES
T3 control. Moreover, some T3 actions upon the organelle are obviously not inducedat the nuclear level. Finally, identification of truncated forms of the nuclear c-ErbAα1 T3 receptor located in the mitochondria is in agreement with the occurrence ofa direct T3 mitochondrial pathway first proposed by Sterling’s group. p43 is an at-tractive candidate to play the role of a T3 receptor in the organelle. This proteinbinds to specific nucleotidic sequences of the mitochondrial genome D-loop, thussuggesting a role as a possible T3-dependent transcription factor.
This direct mitochondrial T3 pathway is well supported by the observation thatp43 overexpression stimulates mitochondrial activity. In addition, it also potientatesmyoblast differentiation, thus suggesting that this pathway could have a significantimportance for cell biology.
Recent data raise another question. Besides p43, a mitochondrial location of thenuclear glucocorticoid receptor has been reported.28 This observation suggests thatother nuclear receptors could be present in the organelle. Therefore numerous ques-tions remain to be solved. What nuclear receptors can be imported into the or-ganelle? How can receptors displaying a nuclear localization signal enter themitochondrion? What molecular mechanisms are involved in their intramitochondri-al action? If they act as hormone-dependent transcription factors of the mitochondri-al genome, do they need interactions with proteins similar to nuclearcorepressors29,30 and coactivators31 involved in the transcriptional activity of nucle-ar receptors? New molecular tools such as p43 may provide several clues in answersome of these questions in the near future.
REFERENCES
1. WRUTNIAK C. & CABELLO G. 1996. La voie d'action mitochondriale directe de la tri-iodothyronine: Mythe ou réalité. Mèdecine-Science 12: 475–484.
2. VAN ITALLIE M.C. 1992. Dexamethasone treatment increases mitochondrial RNAsynthesis in a rat hepatoma cell line. Endocrinology 130: 567–576.
3. CHOU S.H., HANNAH S.S., LOWE K.E., NORMAN A.W. & HENRY H.L. 1995. Tissue-specific regulation by vitamin D status of nuclear and mitochondrial gene expres-sion in kidney and intestine. Endocrinology 136: 5520–5226.
4. EL KEBBAJ M.S., CHERKAOUI MALKI M. & LATRUFFE N. 1995. Effects of peroxi-somes proliferators and hypolipemic agents on mitochondrial inner membranelinked D-3-hydroxybutyrate dehydrogenase (BDH). Biochem. Molec. Biol. Intern.35: 65–77.
5. STERLING K. & MILCH P.O. 1975. Thyroid hormone binding by a component ofmitochondrial membrane. Proc. Natl. Acad. Sci. USA 72: 3225–3229.
6. STERLING K., MILCH P.O., BRENNER M.A. & LAZARUS J.H. 1977. Thyroid hormoneaction: The mitochondrial pathway. Science 197: 996–999.
7. STERLING K. & BRENNER M.A. 1995. Thyroid hormone action: Effect of triiodothy-ronine on mitochondrial adenine nucleotide translocase in vivo and in vitro. Metab-olism 44: 193–199.
8. HAFNER R.P., NOBES C.D., MCGOWN A.D. & BRAND M.D. 1988. Altered relation-ship between protonmotive force and respiration rate in non-phosphorylating livermitochondria isolated from rats of different thyroid hormone status. Eur. J. Bio-chem. 178: 511–518.
9. BRAND M.D., STEVERDING D., KADENBACH B., STEVENSON P.M. & HAFNER R.P.1992. The mechanism of the increase in mitochondrial proton permeability inducedby thyroid hormones. Eur. J. Biochem. 206: 775–781.

99WRUTNIAK et al.: THE T3 MITOCHONDRIAL PATHWAY
10. PARADIES G. & F.M. RUGGIERO. 1989. Decreased activity of pyruvate translocatorand changes in the lipid composition in heart mitochondria from hypothyroid rats.Arch. Biochem. Biophys. 269: 595–602.
11. MARTINO G., COVELLO C., DE GIOVANNI R., FILIPELLI R. & G. PITRELLI. 1986.Direct in vitro action of thyroid hormones on mitochondrial RNA-polymerase.Molec. Biol. Rep. 11: 205–211.
12. GARTSKA H.L., FACKE M., ESCRIBANO J.R. & WIESNER R.J. 1994. Stoichiometry ofmitochondrial transcripts and regulation of gene expression by mitochondrial tran-scription factor A. Biochem. Biophys. Res. Commun. 200: 619–626.
13. FISHER R.P. & D.A. CLAYTON. 1988. Purification and characterization of humanmitochondrial transcription factor 1. Mol. Cell. Biol. 8: 3496–3509.
14. JAKOVILCIC S., SWIFT H.H., GROSS N.J. & RABINOWITZ R. 1978. Biochemical andstereological analysis of rat liver mitochondria in different thryoid states. J. CellBiol. 77: 887–901.
15. STERLING K., CAMPBELL G.A., TALIADOUROS G.S. & NUNEZ E.A. 1984. Mitochon-drial binding of triiodothyronine (T3). Demonstration by electron microscopicradioautography of dispersed liver cells. Cell Tissue Res. 236: 321–325.
16. HASHIZUME K. & ICHIKAWA K. 1982. Localization of 3,5,3′-triiodothyronine recep-tor in rat mitochondrial membrane. Biochem. Biophys. Res. Commun. 106: 920–926.
17. GOGLIA F., TORRESANI J., BUGLI P., BARLETTA A. & LIVERINI G. 1981. In vitrobinding of triiodothyronine to rat liver mitochondria. Pflugers Arch. 390: 120–124.
18. GREIF R.L. & SLOANE D. 1978. Mitochondrial binding sites for triiodothyronine.Endocrinology 103: 1899–1902.
19. WRUTNIAK C., CASSAR-MALEK I., MARCHAL S., RASCLE A., HEUSSER S., KELLER
J.M., FLECHON J., DAUCA M., SAMARUT J., GHYSDAEL J. & CABELLO G. 1995. A43-kDa Protein related to c-erb A α1 is located in the mitochondrial matrix of ratliver. J. Biol. Chem. 270: 16347–16354.
20. RODD C., SCHWARTZ H.L., STRAIT K.A. & OPPENHEIMER J.H. 1992. Ontogeny ofhepatic nuclear triiodothynonine receptor isoforms in the rat. Endocrinology 131:2559–2564.
21. BIGLER J., HOKANSON W. & EISENMAN R.N. 1992. Thyroid hormone receptor tran-scriptional activity is potentially autoregulated by truncated forms of the receptor.Mol. Cell. Biol. 12: 2406–2417.
22. BIANCO A.C. & SILVA J.E. 1987. Intracellular conversion of thyroxine to triiodothy-ronine is required for the optimal thermogenic function of brown adipose tissue. J.Clin. Invest. 79: 295–300.
23. KANEKO T., WATANABE T. & OISHI M. 1988. Effect of mitochondrial protein syn-thesis inhibitors on eythroid differentiation of mouse erythroleukemia (Friend)cells. Mol. Cell. Biol. 8: 3311–3315.
24. CORDEAU-LOSSOUARN L., VAYSSIERE J.L., LARCHER J.C., GROS F. & CROIZAT B.1991. Mitochondrial maturation during neuronal differentiation in vivo and invitro. Biol. Cell 71: 57–65.
25. ROCHARD P., CASSAR-MALEK I., MARCHAL S., WRUTNIAK C. & CABELLO G. 1996.Changes in mitochondrial activity during avian myoblast differentiation: Influenceof triiodothyronine or v-erb A expression. J. Cell. Physiol. 168: 239–247.
26. CASSAR-MALEK I., MARCHAL S., ALTABEF M., WRUTNIAK C., SAMARUT J. &CABELLO G.. 1994. V-erb A stimulates quail myoblast differentiation in a T3 inde-pendent, cell-specific manner. Oncogene 9: 2197–2206.
27. KOROHODA W., PIETRZKOWSKI Z. & REISS K. 1988. Chloramphenicol, an inhibitorof mitochondrial protein synthesis, inhibits myoblast fusion and myotube differen-tiation. Folia Histochem. Cytobiol. 31: 9–13.

100 ANNALS NEW YORK ACADEMY OF SCIENCES
28. DEMONACOS C., R. DJIORDJEVIC-MARKOVIC, N. TSAWDAROGLOU & C. E. SEKERIS.1995. The mitochondrion as a primary site of action of glucocorticoids: The inter-action of the glucocorticoid receptor with mitochondrial DNA sequences showingpartial similarity of the nuclear glucocorticoid responsive element. J. Steroid Bio-chem. Molec. Biol. 55: 43–55.
29. HORLEIN A.J., NAAR A.M., HEINZEL T., TORCHIA J., GLOSS B., KUROKAWA R.,RYAN A., KAMEI Y., SODERSTROM M., GLASS C.K. & ROSENFELD M.G. 1995.Ligand-independent repression by the thyroid hormone receptor mediated by anuclear receptor co-repressor. Nature 377: 397–404.
30. CHEN J.D. & EVANS R.M. 1995. A transcriptional co-repressor that interacts withhormone receptors. Nature 377: 454–457.
31. HORWITZ K.B., JACKSON T.A., BAIN D.L., RICHER J.K., TAKIMOTO G.S. & TUNG L.1996. Nuclear receptors coactivators and corepressors. Mol. Endocrinol. 10: 1167–1177.
32. GADALETA G, PEPE G., DE CANDIA G., QUAGLIARIELLO C., SBISA E. & SACCONE C.The complete nucleotide sequence of the rattus Norvegicus mitochondrial genome:cryptic signals revealed by comparative analysis between vertebrates. J. Mol. Evol.28: 497–516.





























