Embed Size (px)
Citation preview

438 BIOCHIMICA ET IHOPHYSI(~A A( "I'A
BBA 97447
POLYADENYLATE SYNTH1~'TASE DES CELLULES 1)'ASCITE DE SOURIS
I. PURIFICATION ET CARACTt~RISATION DE L'ENZYME
MARIE-LOUISE GIRON ET J, HUPPERT
Laboratoire de Biologie Moldculaire, Institut Guslave Roussy, Groupe de RecheYches No. ~' d~ C.N.R.S.. Villejuif (94) (France)
(Regu le 14 juillet, 1972 )
SUMMARY
Poly (A ) syntheh~se from mouse ascites cells. I. Puri[ication and characterizatio~ o / the enzyme
I. Tile poly(A) synthetase from mouse ascites cells which incorporates i14Ci - AMP into a preexisting polyribonucleotide chain is a weakly acidic protein (isoelectric point = 6 ~: 0.05) with a molecular weight, determined by sucrose gradient centrifu- gation, close to 80 ooo. Its exclusion from Sephadex G-2oo would indicate that it can exist in a complex form. Incubation of the enzyme in a medium of high ionic strength (o.5 M KC1) results in a reversible dissociation. However, centrifugation in a high gravitational field (284 ooo × g) in presence of 0.5 M KC1 leads to an irreversible dissociation of the enzyme.
2. The enzyme is heat labile and loses 50 °/0 of its activity in 4 rain at 45 %;- The polyribonueleic acid primer and ATP have distinct roles in the stabilisation of the enzyme.
3. The enzyme was I5o-fold purified. Nevertheless, it still contains trace amounts of nucleic acids and has slight nuclease activity. Purification of the enzyme results in the appearance of new properties, which are discussed in the second paper.
INTRODUCTION
En 1963, nous avons mis en 6vidence une activit6 enzymatique responsable de la polym6risation de I 'ATP dans les cellules d'ascite Krebs I I de souris, en pr6sence d'un amorceur polyribonud6otidique et d'ions divalents. Cette activit6 enzymatique a 6td signal6e depuis par Horton et al. 1 (1964) comme contaminant au cours de l'6tude de la RNA polym6rase RNA dfpendante induite par le virus de l 'enc@halomyocardite dans ces cellules. Le r61e des enzymes qui semblent diriger spdcifiquement la poly- m6risation d'un seul nucl6otide, notamment les poly(A) synth6tases, reste mal connu. Cependant l'activit6 ATP polym~rasique a 6t6 d6crite dans des tissus tr6s divers: noyaux de thymus de veau 2, membrane chorioallantoide de l 'embryon de poulet a, foie de rat 4 et chez les bact6ries ~,6. Nous avons d'ailleurs montr6 apr&s une 6tude syst6matique de sa distribution, que l'expression de l'activit6 enzymatique semble devoir ~tre reli6e aux processus de la diff6renciation cellulaire v. Depuis la d~couverte en 1966 de poly(A) dans le foie de rat s, puis en 1969 darts le foie de souris 9, une s6rie de
Biochim. Biophys. Acta, 287 (1972) 438-447

POLYADENYLATE SYNTHETASE I 439
travaux a montr6 l'existence de s6quences de poly(A) associ6es aux RNA messagers et aux RNA nucl6aires h4t6rog6nes dans les eellules animales 1°-14. Du poly(A) a 6t6 trouv6 li6 aux RNA messagers de eertains virus /t DNA 16,16 et tout derni6rement, associ6 spdcifiquement au RNA des virus oneog6nes lv,~8.
Lorsque nous avons entrepris l'6tude de la poly(A) synthdtase des cellules d'ascite, nous nous proposions: (I) D'apporter des renseignements sur la structure et les propri6t6s de la prot6ine enzymatique sur lesquelles aucunes donn6es n'avaient 6t6 publi6es. Depuis, des rdsultats concernant la structure de l 'enzyme bact~rien ont 6t6 apport6s (Terzi el al. ~9, 197o ). (2) De faire une 6tude approfondie de la r6aetion enzymatique.
Nous allons d6erire maintenant la purification et earactdrisation de l'enzyme. La communication suivante d6crira la r6action enzymatique et le m6canisme d'action de l'enzyme.
MAT1~RIELS ET MI~THODES
Matdriel cellulaire Cellules d'ascite tumorale de la souris, ascite Krebs II (Klein el a/. 2°) et ascite
FLS (Lacour et al.21). Les cellules d'ascite pr61ev6es le 74me jour apr6s l'inoculation et lav6es, 6taient conserv6es sous forme de culot cellulaire ~t --20 °C.
Produits ugilisds Les nucl6osides triphosphates marqu6s au J4C: Schwarz Bioresearch, Inc., puis
New England Nuclear Corporation; nucl6osides mono- di- et triphosphates non radio- actifs: Sigma Chemical Company; les polynucl6otides ainsi que les oligonuclfiotides: Miles Chemical Company. Les RNA de diff6rentes origines ont 6t6 pr6parfs au labora- toire par extractions r6p6t6es au ph6nol et pr6cipitations /t l'alcool (Harel et a1.22).
Essai d'incorporation de I ' A M P Le m61ange r6actionnel contenait (sauf modifications indiqu6es dans le texte)
dans 0.4 ml: 5o/,moles Tris (pH 8.4); 0.5/,moles MnC12; I / ,mole fl-mercapto6thanol; 50/,g polyribonucl6otide; 0.2/,moles E14CIATP (radioactivit6 sp6cifique = 500 ~ IOOO epm/nmole; prot6ine enzymatique, concentration indiqude pour chaque expdrience; de m6me pour MgC12. Les incubations 6taient effectu6es ~ 37 °C.
La r6action d'incorporation de [14CLAMP 6tait arr~t6e par pr6cipitation du polym~re form6 par HC104 3.5 % froid, en pr6sence de I mg de s6rum albumine. Le
O/ pr6cipitd 6tait lay6 avec "~ × 4 ml HC104 3.5 /o et dissous dans 0.22 ml NH4OH 0.25 M. La radioactivit6 6tait mesur6e dans un compteur Tracerlab ~t fen~tre mince (rendement environ 25 %).
Dosage des protdines La quantit6 de prot6ine 6tait d6terminfie par mesure de l'absorption /~ 28o et
26o nm, en faisant intervenir une correction pour la teneur en acides nuel6iques 23, oll par la m6thode de Lowry et al.24).
Biochim. Biophys. Acta, 287 (1972) 438-447
44 ° M.-L. GIRON, J. HUPPERT
Electro / ocal isat ion La m6thode d6velopp6e par Svensson zs pour la s6paration des prot6ines par
61ectrophor6se dans un gradient combin6 de pH et de saccharose, a 6t6 utilis6e. La colonne ~t 61ectrophor6se, de marque LKB, avait une contenance de IiO ml. La concentration finale des ampholines 6tait 6gale ~ 1 % . L'61ectrofocalisation 6tait r6alis6e en 5 jours sous 800 V ~t 4 °C. Le contenu de la colonne 6tait ensuite collect6 en fractions de 15o gouttes ~ l'aide d'une pompe Technicon. Sur chaque fraction, 6taient d6termin6es: la teneur en plot6ine (par la m6thode de Lowry et al. 2., pr6c6d6e d'une pr6cipitation pr6alable des protdines par de l'acide trichloroac6tique 7 °'o) ; la valeur du pH, ~ 4 et 2o °C, avec un pH m6tre Radiometer, type TTT IC. ; l'activit6 enzymatique.
Electrophor}ses en gel de polyacrylamide Les 61ectrophorbses 6taient r6alis6es selon la mdthode de Davis ~6 dans un appa-
reil Quickfit. Les produits utilis6s provenaient de Eastman Kodak Company.
R~SULTATS
Purification de l'enzyme Les diff6rentes 6tapes de la purification ont 6td effectu6es ~t 4 °C. L'unit6 d'enzyme a 6t6 ddfinie comme suit: Une unit6 d'enzyme est la quantit6
d'enzyme permettant l 'incorporation de I nmole de ~1~C~AMP apr~s 3 h d'incubation 37 °C,
L'activit6 sp6cifique est repr6sent6e par le nombre de nmoles de E14C~AMP incorpor6 par I mg de prot6ine pendant 3 h ~. 37 °C. Le bilan complet d'une purification est rdsum6 dans le Tableau I.
T A B L E A U 1
BILAN D'UNE PURIFICATION D'•NZYME
Fractions d'enzyme
Extrait Surnageant Surnageant Sulfate Sephadex D EA E- brut Spinco sulfate de d'ammo- G-75 cellulose dialysd strepto- nium KC1 o.z M
mycine 25-4 ° %
t~ = 28o[26o 0.758 o,82o 0.95 ° 1.48 1.58 1.35
Quan t i t~ to ta le de pro t6 ines (mg) 5060 263 ° 888 151 141 15.7
R e n d e m e n t pa r r a p p o r t ioo °/o 52 °/o 17. 5 % 2.98 % 2.8 % 0.30 % a u x pro t6 ines ini t iales
Uni tds to ta les d ' e n z y m e 19 700 I6 900 13 800 14 2oo 12 600 9720
iRendement pa r r a p p o r t a u x un i tds d ' e n z y m e ini t iales ioo % 86 % 7 ° % 72 % 64 % 49.4 %
Act iv i t6 spdcif ique 3.91 7.8 lO.3 lO4 135 585
Pur i f i ca t ion 2 2.6 26 34 15 °
Biochim. Biophys. Acta, 287 (1972) 438-447

POLYADENYLATE SYNTHETASE I 441
(a) Prdparation de l'extrait brut. Les cellules d'ascite gard~es congel~es en culot, sont mises en suspension dans deux volumes de Tris o.ooi M (pH 7-4), MgC1, o.ooi M et fl-mercapto6thanol o.ooi M e n pr6sence de bentonite (3 mg/ml), puis elles sont broyfes avec des billes de verre pendant 15 min/ t 4 °C (2000 secousses/min.). Apr~s une centrifugation de 30 rain & 45 ooo × g dans une centrifugeuse Sorvall, le surna- geant est dialys~ contre un tampon Tris o.oi M (pH 7.4), MgC12 o.oi M et fl-mercapto- 6thanol o.ooi M. Lorsque la purification est poursuivie, la dialyse est effectu6e contre le tampon prfc6dent comprenant MgCI, o.ooi M, afin d'6viter la pr6cipitation de nucl~oprot~ines. Apr~s ~limination des ribosomes par centrifugation (I h 5 15o ooo × g. Spinco rotor L 5o), les acides nucl6iques sont 61imin6s de la pr6paration par pr6cipi- ration au sulfate de streptomycine (sulfate de streptomycine Specia--solution/t 5 % pr~par6e extemporan6ment et amen6e ~ pH neutre avec NH4OH 0.25 M). La pr6cipi- tation est effectu6e sous agitation pendant IO min avec une concentration finale en sulfate de streptomycine 6gale ~ 0.75 %. Le surnageant recueilli apr~s une centrifu- gation de IO rain /t 24 ooo × g est dialys6 contre un tampon Tris o.oi M (pH 7.4), MgC12 o.oi M, fl-mercapto6thanol o.ooi M.
(b) Prdcipitation ]ractionnde au suljate d'ammonium. La prot6ine enzymatique pr6cipite dans la fraction 25-4 ° ~/o de saturation en sulfate d'ammonium .Le pr6cipit~ recueilli par centrifugation (15 rain /t 27 ooo × g) est dissous dans un tampon Tris 0.025 M (pH 7.4), MgC12 o.oi M e t fl-mercapto~thanol o.ooi M, puis dialys6 contre ce mSme tampon pendant 2/~ 3 h.
(c) Filtration sur gel de Sephadex G-75. La fraction active est plac6e sur une colonne de Sephadex G-75 (1.5 cm × 22 cm) pr6alablement 6quilibrde avec un tampon Tris 0.025 M (pH 7.4) MgCI~ o.oi M, /q-mercapto~thanol o.ooi M. La prot6ine enzy- matique est exclue du gel (Fraction G-75 ).
(d) Chromatographie sur colonne de DEAE-cellulose. La fraction G-75 est dia- lys6e contre un tampon Tris 0.05 M (pH 8.4), fl-mercapto6thanol o.ooi M e t placde sur une colonne de DEAE-cellulose (I.5 cm × 23 cm) 6quihbr6e dans le m~me tampon. La prot~ine enzymatique est 61u6e par du KC1 o.I Me t pr6cipitde par addition d'un volume 6gal d'une solution satur6e de sulfate d'ammonium amen~e ~ pH neutre. Le pr6cipit6 recueilli par centrifugation (15 rain/t 27 ooo ×g) est dissous dans un Tam- pon Tris. o.oi M (pH 7.4), fl-mercapto~thanol o.ooi M contenant du glyc6rol 50 %.
(e) Rdsultats de la puri/ication. Le taux de la purification est d'environ 15o lois (Tableau I). Le rapport A2so/A26 o a progress6 de o.718 h 1.35. Mais la valeur relative- ment peu 61ev6e de ce rapport montre que la pr6paration n'est pas enti~rement d6pourvue d'acides nucl6iques. De m6me, elle n'est pas exempte de route activit6 nucl6asique. La fraction DEAE-cellulose est instable mais la pr6sence de glycerol permet une conservation satisfaisante de l'activit6 enzymatique ~ --20 °C.
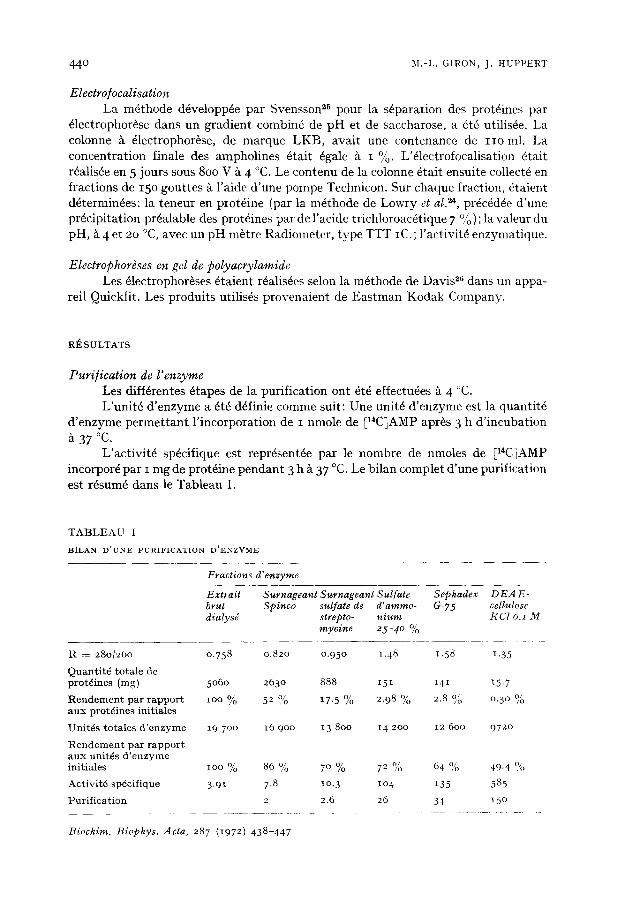
Caractdrisation de l'enzyme (a) Ddtermination de son point isodectrique. La valeur du point iso~lectrique de
la poly(A) synth6tase a 6td d6termin~e par 61ectrofocalisation d'une fraction enzyma- tique partiellement purifi6e (Fraction S. Ammo) (Fig. I). Cette valeur est 6gale /~ 6 i 0.05. Au cours de l'61ectrofocalisation, une perte importante d'activit6 enzy- matique a lieu (30 % d'activit6 initiale retrouv6e). L'exposition prolong6e a 4 °C peut en 6tre la cause ainsi que le processus 61ectrophor~tique lui-m~me (certaines alt6rations mol6culaires ont 6t6 signal~es dans le cas de la c6ruloplasmine27).
Biochim. Biophys. Acta, 287 (I972) 438-447
442 M.-L. G1RON, J, HUPPKRT
pH c p m
.1 000
- 800
.600
.400
-200 /
1"0 Z'0 3"0 4"0 50 6"0 f rac t ions
Fig. I. D6terminat ion du point iso6lectrique de l 'enzyme par ~lectrofocalisation. 20 mg d 'une fraction S. Amino (AS ~ I9O), on t 6t6 soumis ~ une ~lectrophor6se dans un gradient de p H de 5-8, pendan t 5 jours sous 800 V. Aprgs arr~t de l'61ectrophor~se, des fractions de 15o gout tes sont recueillies. On mesure sur chaque fraction: Le pH, ~t 4 °C (m • . . m) et k 2o °C ( O - O I ; et l 'activit6 enzymat ique sur une part ie aliquote de 0.05o ml, dans les conditions habituelles ((2)-@).
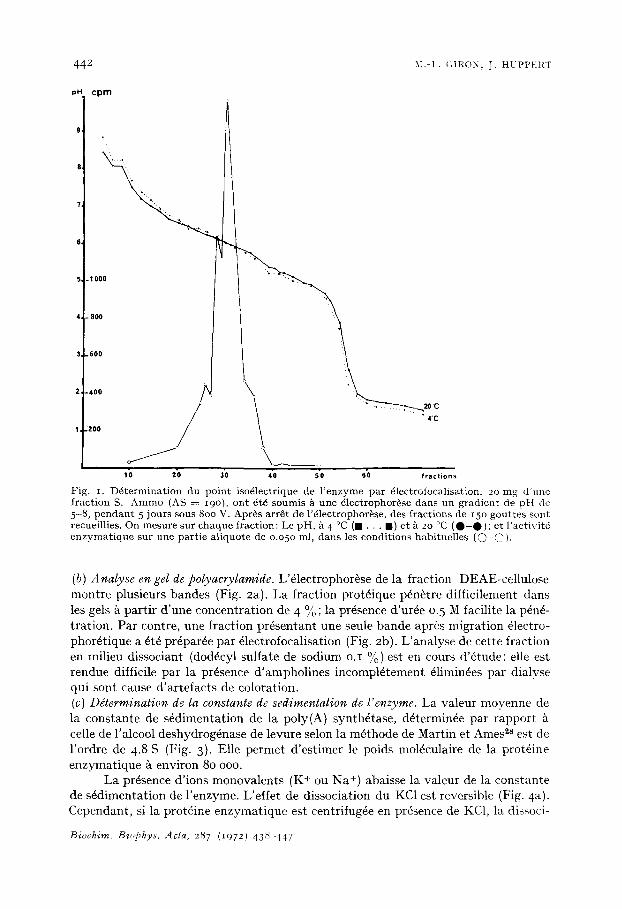
(b) Analyse en gel de polyacrvlamide. L'61ectrophor~se de la fraction DEAE-cellulose montre plusieurs bandes (Fig. 2a). La fraction prot6ique p6n~tre difficilement dans les gels k partir d'une concentration de 4 %; la pr6sence d'ur6e 0.5 M facilite la pdn6- tration. Par contre, une fraction pr6sentant une seule bande aprbs migration 61ectro- phor6tique a 6t6 pr6par6e par 61ectrofocalisation (Fig. 2b). L'analyse de cette fraction en milieu dissociant (dod6cyl sulfate de sodium o.I °/o ) est en cours d'dtude; elle est rendue difficile par la pr6sence d'ampholines incompl6tement 61imin6es par dialyse qui sont cause d'artefacts de coloration. (c) Ddtermination de la constante de sedimentation de/'enzyme. La valeur moyenne de la constante de s6dimentation de la poly(A) synthdtase, d6termin6e par rapport celle de l'alcool deshydrog6nase de levure selon la m6thode de Martin et Ames ~8 est de l'ordre de 4.8 S (Fig. 3). Elle permet d'estimer le poids mol6culaire de la prot6ine enzymatique ~t environ 80 ooo.
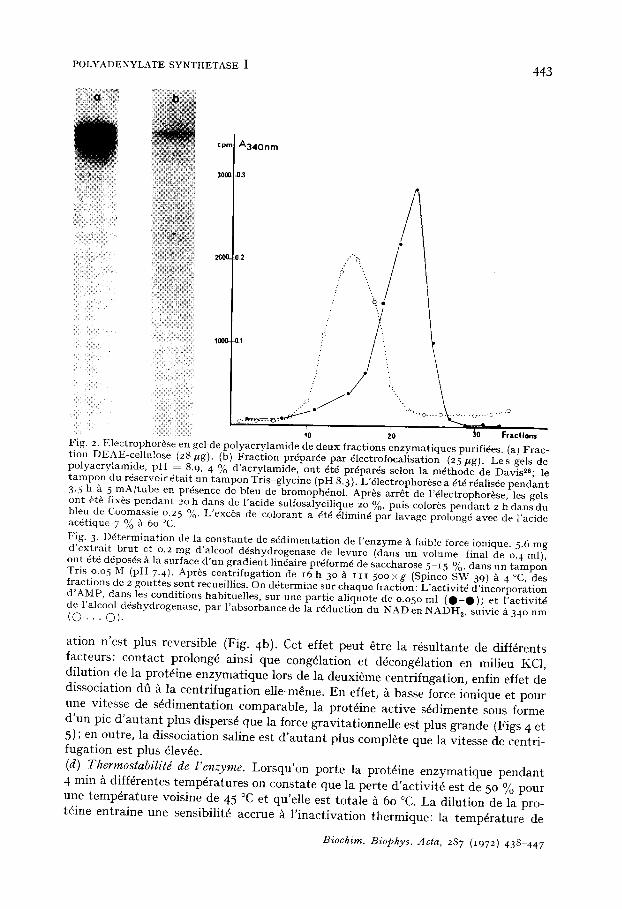
La pr6sence d'ions monovalents (K+ ou Na +) abaisse la valeur de la constante de s6dimentation de l'enzyme. L'effet de dissociation du KC1 est reversible (Fig. 4a). Cependant, si la prot6ine enzymatique est centrifug6e en p%sence de KC1, la dissoci-
Biochim. Biophys. Acta, 287 (1972) 438 -447
POLYADENYLATE SYNTHETASE I
ili~ii]i~!ii;iiii: ~
~!ii!iii!iiiiiii!~iiiii~i~iiill
ii! i i!iii~i ~i~ii!~! ~
~ii ~ iliiii ~ ~
i~i i~ill I~
~iiiiiiTiiiiiiiiiiiiiii~"~'~'~iiiiiiiiiiiiiiiiiiii
:tiiiiiii~ ~,5
iiiiiiiii~ili:il~;!~ d)!!!!
i'!i!ii iiiiii!~i iii ~!~!!!iiii fli!l![iii!ii!iiii;]i~!!ii!iii!i!iiii!i~!:!i~
~!iliiiiii!ii!iiii!iiiii~ii!ii~ii~iiiili!iiiiiiii~ii
~i!iiiiii diiiiiiii~i~ii;ii:i~i
!
cpm A 3 4 0 n m
3000 -0.3
2~00. 0,2
1000- 0.1
443
©
......... o .MI j ' /
/ / . " ' b
..c;" '..
:" ",.
b ".1
""" 'J" "• o . . . . o i , : , - o II
~0 20 10 Fractions - - Fig. 2. Electrophor~se en gel de po lyacry lamide de deux fractions e n z y m a t i q u e s purifi~es. (a) Frac- t ion D E A E - c e l l u l o s e (28/2g). (b) Fract ion pr~par6e par 61ectrofocalisation (25 t'g). L e s gels de po lyacry lamide , p H = 8.9, 4 % d'acrylamide, ont 6t6 pr6par6s selon la m6thode de Davis26; le tanlpon du r6servoir ~tait un tampon Tr i s -g lyc ine (pH 8.3). L'61ectrophor~se a ~t6 r~alis6e pendant 3.5 h /L 5 m A / t u b e en pr6sence de bleu de bromoph~nol . Apr~s arrSt de l '~lectrophor~se, les gels ont ~t~ fixes pendant 2o h dans de l'acide sulfosalyci l ique 2o %, puis color , s pendant 2 h dans du bleu de Coomassie o.25 %. L'excbs de colorant a 6t~ 61imin~ par lavage prolong6 avec de l 'acide ac~t ique 7 % 5 6o °C.
Fig. 3- D~terminat ion de la cons tante de s6dimentat ion de l ' e n z y m e / t faible force ionique. 5.6 mg d'extrai t brut et o.2 mg d'alcool d~shvdrogenase de levure (dans un vo lume final de o. 4 ml), ont ~t6 d6pos6s/~ la surface d'un gradient lin6aire pr6form6 de saccharose 5 -15 %, dans un tampon Tris o.o 5 M (pH 7-4). Apr~s centri fugat ion de I 6 h 3o ~ I i i 5 o o × g (Spinco SW 39) /t 4 °C, des fract ions de 2 gout tes sont recueillies. On d6termine sur chaque fraction: L'act ivi t~ d' incorporat ion &, dans les condit ions habituel les , sur une partie a l iquote de o .o5o ml ( ~ - ~ ) ; et l 'activit6 de l'alcool d~shydrogenase , par l 'absorbance de la r~duction du N A D en NADH~, suivie/~ 34o nm ( 0 . . . 0 ) .
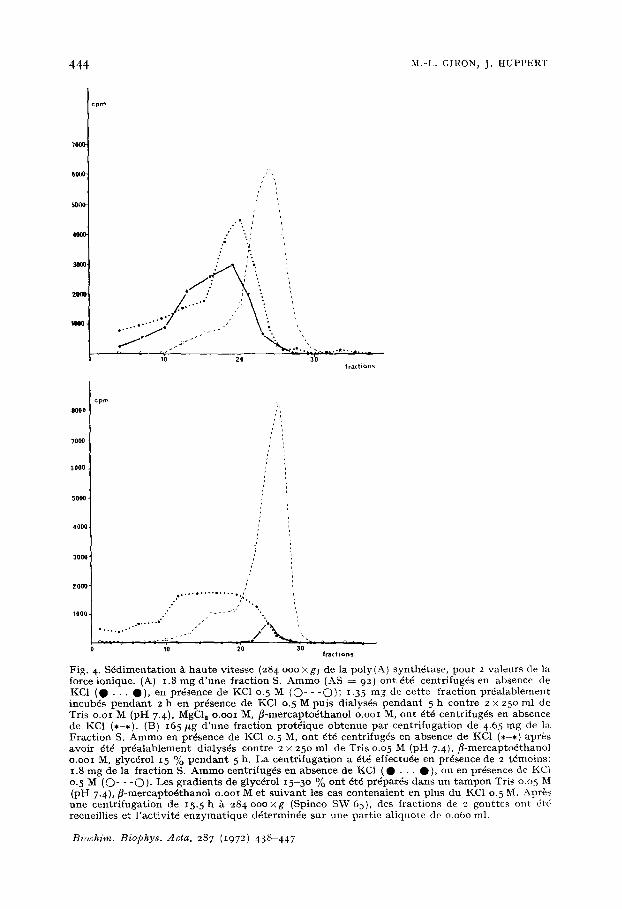
ation n'est plus reversible (Fig. 4b). Cet effet peut 8tre la r6sultante de diff6rents facteurs: contact prolong~ ainsi que cong61ation et d6cong61ation en milieu KC1, dilution de la prot~ine enzymatique lors de la deuxi~me centrifugation, enfin effet de dissociation dfi ~ la centrifugation elle-mSme. En effet, 5. basse force ionique et pour une vitesse de s6dimentation comparable, la prot6ine active s~dimente sous forme d'un pie d'autant plus dispers6 que la force gravitationnelle est plus grande (Figs 4 et 5); en outre, la dissociation saline est d'autant plus complete que la vitesse de centri- fugation est plus 61ev6e. (d) Thermostabilild de l'enzyme. Lorsqu'on porte la prot6ine enzymatique pendant 4 min/t diffdrentes temp6ratures on constate que la perte d'activit6 est de 5o % pour une temp6rature voisine de 45 °C et qu'elle est totale/~ 6o °C. La dilution de la pro- t6ine entraine une sensibilit~ accrue ~ l'inactivation thermique: la temp@ature de
Biochim. Biophys. Acta, 287 (1972) 4 3 8 - 4 4 7
444 M.-L. GIRON, j . HUPPERT
60~0-
5000-
4000"
3O0O
2 0 0 0
c p m
. . ' .
. ; • ,
:" 7".
. o: . "°''''' "'. .:.]],, . . . . . ~ : . . . . . . .
~o io ~'o f r a c t i o n s
I
0ooo i
7oo~ "
5 0 0 0 .
5000 -
4 0 0 0
3000"
2000"
1000
cpo~
?
/ ', / i
. . . . . . . . . . . . . . . "/--.o ', • . . . . . /
......... / , . . , _ } • . . . . o . . ° ~ ,
. . . . . : --- , ? e.~, . lO ~2'o 30
f ract ions
Fig. 4. S4dimentat ion ~ hau te vitesse (284 ooo xg) de la poly(A) synth4tase, pour 2 valeurs de la force ionique. (A} 1.8 mg d 'une fract ion S. Amino (AS ~ 92) ont 4t4 centrifug4s en absence de KC1 ( 0 • • • O) , en pr4sence de KC1 0. 5 M (O- - - O ) ; 1.35 mg de cette fraction pr4alablement incubds pendan t 2 h e n pr4sence de KC1 o. 5 M puis dialys4s pendan t 5 h contre 2 x 250 ml de Tris o.oi M (pH 7.4), MgC12 o.ooi M, f l-mercaptodthanol o.ooi M, ont 4t4 centrifug4s en absence de KC1 (*-*). (B) i65/2g d 'une fraction prot4ique obtenue par centr i fugat ion de 4.65 mg de la Fract ion S. A m m o en pr4sence de KCI 0.5 M, ont ~t4 centrifug4s en absence de KC1 (*-*) apr~s avoir 4t4 pr4alablement dialysds contre 2 x 250 ml de Tris 0.05 M (pH 7.4), f l-mercaptodthanol o.ooi M, glycdrol 15 % pendan t 5 h. La centrifugation a 6t4 effectu4e en pr4sence de 2 t4moins: 1.8 mg de la fraction S. Amino centrifug4s en absence de KC1 ( • . . . • ) , ou en pr6sence de KC| 0-5 M (O- - - O ) . Les gradients de glyc4rol 15-3o % ont 4t~ pr4par6s dans un t a m p o n Tris 0.05 M (pH 7.4), f l -mercapto6thanol o.ooI IV[ et su ivan t les can contenaient en plus du KC1 0.5 M. Apr~s une centr i fugat ion de 15.5 h ~ 284 o o o x g (Spinco SW 65), des fractions de 2 gout tes out 4t4 recueillies et l 'activit6 enzymat ique d4terminde sur une part ie aliquote de 0.060 ml.
Biochirn. Biophys. Acta, 287 (1972) 438 447
POLYADENYLATE SYNTHETASE I
A280nm;cpm
0,I00
I000
.500 /
2'o
/
P
~t
/t . \ .. \
4'0
445
A28O n m c p m . ~ ~
/
.IODO i I
/
f o.loo 500 ,,.,, f\\ I, ,,' /
l, ~ " F.p / 2~0 4 ~0
e-e. o
6~ |ractions
i ' . . f
%activite °
100 ~ o ">'- 4t
°•o 8 ~
50. ~' " ' ' ' J l t 2
" ~- '~34 o
temps ( r rm)
~o activit~ 100.
50-
".'~
- 0 3
W-.. "•.o
6'0 f ract ions t :~ t emps (rr'~ )
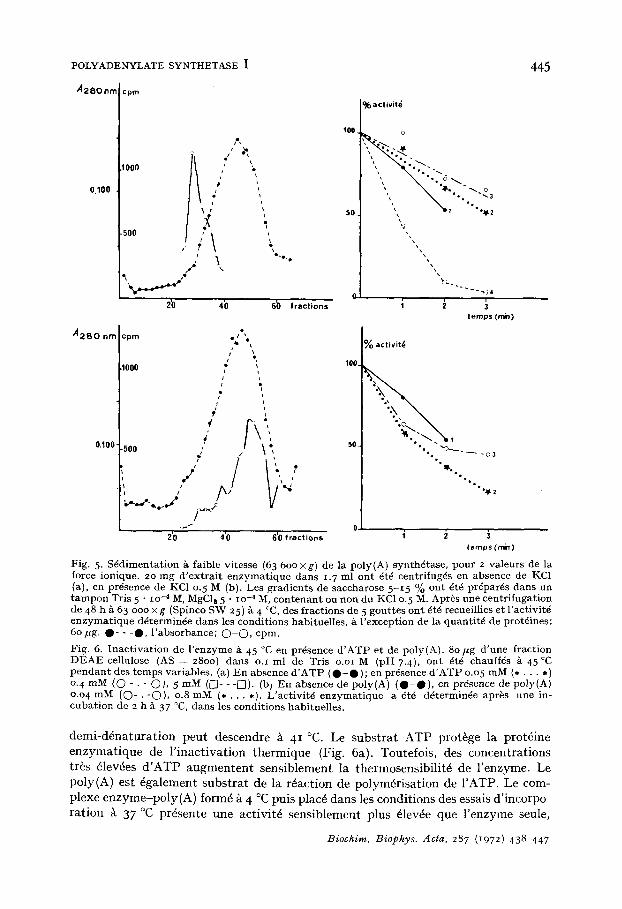
Fig. 5. S6dimentat ion g faible vitesse (63 600 xg ) de la poly(A) synth6tase, pour 2 valeurs de la force ionique. 20 ing d ' ex t ra i t enzymat ique darts 1. 7 ml ont 6t~ centrifug6s en absence de KC1 (a), en pr6sence de KC1 o. 5 M (b). Les gradients de saccharose 5-15 % ont 6t6 pr6par6s darts un t a m p o n Tris 5 " lO-4 M, MgCle 5 " lO-4 M, con tenant ou non du KC10. 5 M. Apr6s une centr ifugat ion de 48 h ~ 63 ooo x g (Spinco SW 25) a- 4 °C, des fract ions de 5 gout tes ont 6t6 recueillies et l 'activit6 enzymat ique d6termin6e darts les condit ions habituelles, ~ l 'exception de la quant i t6 de prot6ines: 6o/ tg. 0 - - - 0 , l ' absorbance; O - O , cpm.
Fig. 6. Inac t iva t ion de l ' enzyme ~ 45 °C en pr6sence d ' A T P et de poly(A). 80 yg d 'une fraction DEAE-cel lulose (AS = 28oo) dans o.i ml de Tris o.oi M (pH 7.4), ont 6t6 chauff6s A 45¢C p e n d a n t des t emps variables. (a) E n absence d ' A T P ( O - • ); en pr6sence d ' A T P o.o 5 mM (* . . . *) 0. 4 mM (O - • - O) , 5 mM ([]- - -F/). (b/ En absence de poly(A) ( O - O ) , en presence de poly(A) o.o4 mM (O- . -O) , o.8 mM ( . . . . . ) . L 'act ivi t6 enzymat ique a 6t~ d6termin6e apr~s une in- cubat ion de 2 h ~ 37 °C, dans les condit ions habituelles.
demi-d6naturation peut descendre ~ 41 °C. Le substrat ATP prot&ge la prot6ine enzymatique de l'inactivation thermique (Fig. 6a). Toutefois, des concentrations tr~s 61ev6es d'ATP augmentent sensiblement la thermosensibilit6 de l'enzyme. Le poly(A) est 6galement substrat de la r~action de polym6risation de I'ATP. Le com- plexe enzyme-poly(A) form6 ~ 4 °C puis plac6 dans les conditions des essais d'incorpo ration ~ 37 °C pr6sente une activit6 sensiblement plus 61ev~e que l'enzyme seule,
Biochim. Biophys. Acta, 287 (I972) 438-447

446 M.-1.. (;IRON, .I. HUPPERT
toutes conditions ~tant ~gales par ailleurs (notainment en ce qui con,'erne la concen- tration finale cn poly(A)). Touteiois, le COlnplexe enzyme poly(A) pr6sente une courbe d'inactivation thermique plus accentude que l 'enzyme seule (Fig. 6b).
DISCUSSION
La poly(A) synth6tase des cellules d'ascite telle que nous l 'avons caraet6ris~e est une prot6ine faiblement acide, de poids mol6culaire d'environ 8o 000. Mais cer- taines de ses propri6t6s (exclusion du Sephadex G-200, difficult6 de p~n6tration dans les gels de polyacrylamide, effet de la force gravitationnelle sur son profil de s6dimen- tation ~ basse force ionique) tendent ~ prouver qu'elle peut exister sous forme plus complexe. La vitesse de sddimentation de la protdine est abaiss6e en pr6sence de KC1; cette dissociation est reversible. La dissociation de la prot6ine peut 6tre accen- tu~e sous l 'action de la force gravitationnelle et devenir irr6versible. Mais d 'autres facteurs, notamment la dilution de la protdine, peuvent dgalement en 8tre responsa- bles. Actuellement, la composition en sous-unitds de la prot6ine ne peut cependant pas 8tre affirinde d'apr~s l 'analyse 61ectrophor6tique en gel de polyacrylamide.
Les pr@arations d 'enzyme semi-purifi~es sont stables bien que certaines modifi- cations des propri~t~s cin6tiques apparaissent durant la conservation ~ --2o °C (M. L. Giron et J. Huppert , ~ paraitre). Au tours de la purification la prot~ine enzymatique devient instable/~ partir du stade de la prdcipitation au sulfate d 'ammonium. Cette dtape correspond h l 'dlimination de nombreuses prot6ines inactives et d'une grande partie des acides nucl6iques. Cependant, la valeur relativement peu 61evde du rapport A28o/Ai6o indique la pr6sence d'acides nucl6iques rdsiduels. Ceux-ci ne suppriment pas la n6cessit6 d'un anlorceur polyribonucl6otidique exog~ne pour l 'incorporation d'AMP, mais ils pourraient ~tre n6cessaires/~ la stabilisation de la prot6ine enzyma- tique. L'existence d'acides nucl6iques intimement lids /t la prot6ine enzymatique emp&he actuellement d'affirmer qu'elle est compos~e de sous-unitds. La dissociation observ6e 5 force ionique 6lev6e ou par centrifugation, pourrait en effet correspondre
une dissociation acides nucl6iques prot6ine. L'dtude de la thermostabilit6 de la poly(A) synth6tase a montr6 que celle-ci
commence ~ ~tre inactiv6e par chauffage /~ des temp6ratures relativement basses. Elle a mis en dvidence l'influence de la concentration en protdine et le r61e distinct des deux substrats de la rdaction, ATP et poly(A), dans la stabilisation de la structure de la prot6ine enzymatique.
A notre connaissance il n 'y a pas de donn6es concernant les propri6tfis physico- chimiques et la structure des poly(A) synth6tases d'origine animale ou v6g6tale. I1 faut noter cependant le cas de l 'enzyme ad6nylant des plants de mais 29, enzyme qui utilise prdf&entiellement le tRNA ou les d6oxyoligonucl6otides comme amorceurs de la polym6risation d 'ATP. L'hypoth~se a 6t6 avanc6e ~ part ir de donn6es indirectes que la prot6ine enzymatique ferait partie d'un complexe prot~ique poss~dant une activit6 RNA polym6rasique. En ce qui concerne les bact6ries, la poly(A) synthdtase serait form6e de plusieurs sous-unit6s dont certaines seraient communes i~ la RNA poly- mdrase (Terzi et al.~9). I1 a dt6 sugg&~ qu la poly(A) synth6tase bact~rienne ne serait qu'une iorme modifide de la RNA polym6rase, dont le r61e serait distinct de la simple production de poly(A), ceci d 'au tant plus que la prdsence de poly(A) n 'a pu fitre raise en 6vidence chez les bactdries,
Biochim. Biophys. Acta, 287 (r972) 438-447

POLYADENYLATE SYNTHETASE I 447
It semble que la situation soit diff6rente dans le cas des cellules animales. Le poids inol6culaire de la poly(A) synth6tase des cellules d'ascite parait beaucoup plus faible que celui des RNA polym6rases. La pr6sence de s6quences de poly(A) li6 aux RNA messagers et aux RNA nucl6aires h4t6rog6nes peut faire penser que la poly(A) synth6tase aurait une existence propre ou, en tout cas, un r61e sp6cifique dans les cellules d 'Eukaryotes.
R~;SUME
I. L a p o l y ( A ) s y n t h 6 t a s e des ce l lu les d ' a s c i t e de sour i s , r e s p o n s a b l e de l ' i n c o r p o -
r a t i o n de [14CLAMP s u r u n e c h a i n e p o l y r i b o n u c l 6 o t i d i q u e p r 6 e x i s t a n t e , es t u n e pro-
t 6 i n e f a i b l e m e n t ac ide ( p o i n t i so61ec t r ique == 6 ± 0.05), de pois m o l d c u l a i r e v o i s i n
de 80 ooo. Mais son e x c l u s i o n d ' u n e c o l o n n e de S e p h a d e x G-2oo n o t a m m e n t , i n d i q u e -
r a i t qu ' e l l e p e u t e x i s t e r sous f o r m e p lus c o m p l e x e . L a v i t e s s e de s 6 d i m e n t a t i o n de la
p r o t 6 i n e e n z y m a t i q u e es t aba i s sde ~ force i o n i q u e 61ev6e. C e t t e d i s s o c i a t i o n p e u t
~ t re r6ve r s ib l e . Mais la c o m p o s i t i o n en s o u s - u n i t 6 s ne p e u t 8 t re a c t u e l l e m e n t a f f i rm6e .
2. L a p r o t ~ i n e e n z y m a t i q u e es t t h e r m o s e n s i b l e : elle p e r d 5 ° % de son a c t i v i t 6
a p r ~ s c h a u f f a g e p e n d a n t 4 r a i n ~l 45 °C. L ' A T P e t l ' a m o r c e u r p o l y r i b o n u c l ~ o t i d i q u e
o n t des r61es d i s t i n c t s d a n s la s t a b i l i s a t i o n de la s t r u c t u r e de la p ro t6 ine .
3. L a p r o t 6 i n e e n z y m a t i q u e a 6t6 pur i f ide e n v i r o n 15o fois m a i s la f r a c t i o n
pur i f ide c o n t i e n t e n c o r e des t r a c e s d ' a c i d e s n u c l 6 i q u e s e t d ' a c t i v i t 6 n u c l 6 a s i q u e . L a
p u r i f i c a t i o n a e n t r a i n 6 des m o d i f i c a t i o n s de l ' a c t i v i t 6 e n z y m a t i q u e e t l ' a p p a r i t i o n
de p r o p r i 6 t 6 s n o u v e l l e s d o n t l ' S t u d e fa i t l ' o b j e t de l ' a r t i c l e s u i v a n t .
B I B L I O G R A P H I E
i E. Horton, S. L. Liu, L. Dalgarno, E. M. Martin et T. S. Work, Nature, 204 (1964) 247. 2 M. Edmonds et R. Abrams, J. Biol. Chem., 235 (196o) 1142. 3 P. R. Venka ta raman et N. R. Mahler, ]. Biol. Chem., 238 (1963) lO58. 4 H. G. Klemperer, Biochim. Biophys. Acta, 72 (1963) 406. 5 M. E, Gottesman, Z. N. Canellakis et E. S. Canellakis, Biochim. Biophys. Acta, 61 (1962) 34. 6 J. T. August, P. J. Ortiz et J. Hurwitz, J. Biol. Chem., 237 (1962) 3786. 7 M. L. Giron et J. Huppert , C. R. Acad. Sci., 261 (1965) 1451. 8 A. Hadjivassiliou et G. Brawernaan, J. Mol. Biol., 20 (1966) i. 9 L. Lim, Z. N. Canel]akis et E. S. Canellakis, Biochem. Biophys. Res. Commun., 34 (1969) 536.
IO L. Lira et E. S. Canellakis, Nature, 227 (197 o) 71o. i i H. Burr et J. B. Lingrel, Nature, 233 (1971 ) 41 . 12 J. E. Darnell, R. Wall et R. J. Tushinski, Proc. NatlAcad. Sci. U.S., 68 (1971) 1321. 13 M. Edmonds, M. H. Vaughan, J. R. et H. Nakazato, Proc. Natl Acad. Sci. U.S., 68 (1971) 1336. 14 S. Ylee, J. Mendecki et G. Brawerman, Proc. Natl Acad. Sci. U.S., 68 (1971) 1331. 15 J. Kates, Cold Spring Harbor Syrup. Quant. Biol., 35 (197 o) 743. 16 L. Philipson, R. Wall, G. Glickman et J. E. Darnell, Proc. Natl Acad. Sci. U.S., 68 (1971) 2806. 17 M. C. Lai et P. H. Duesberg, Nature, 235 (1972 ) 383 • 18 D. Gillespi, S. Marshall et R. C. Gallo, Nature, 236 (1972) 227. 19 M. Terzi, A. Cascino et C. Urbani, Nature, 226 (197o) lO52. 20 C. Klein et E. Klein, Cancer Res., i i (1951) 466. 21 F. Lacour, J. Lacour, J. Harel et J. Huppert , J. Natl. Cancer Inst., 24 (196o) 3oi. 22 J. Harel, L. Harel, F. Lacour, A. B~er et J. Imbenotte , J. Mol. Biol., 7 (1963) 645. 23 O. Warburg et W. Christian, Biochem. Z., 31o (1941) 384. 24 O. H. Lowry, N. J. Rosebrough, A. L. Farr et R. J. Randall, J. Biol. Chem., 193 (1951) 265. 25 H. Svensson, Arch. Biochem. Biophys. Suppl., i (1962) 132. 26 B. J. Davis, Ann. N. Y. Acad. Sci., 121 (1964) 404 . 27 L. Pejaudier, R. Audran et M. Steinbuch, Biochim. Biophys. Acta, 229 (1971) 437. z8 R. G. Martin et B. N. Ames, J. Biol. Chem., 236 (1961) 1372. 29 R. J. Mans, Biochem. Biophys. Res. Commun., 45 (1971) 980.
Biochim. Biophys. Acta, 287 (1972) 438-447











![Traitement de la première poussée d'ascite - FMC-HGE · Singal AK et al. Hepatology 2012 [ePub] Survie TH pour cirrhose ... (dans les 12 heures) ... • Seul le traitement de la](https://img.pdfslide.fr/doc/110x75/5b9c454e09d3f2194e8bfc4f/traitement-de-la-premiere-poussee-dascite-fmc-singal-ak-et-al-hepatology.jpg)






