Embed Size (px)
Citation preview

Em. J. Biochem. 14 (1970) 197-204
Regulation of Histidine Uptake by Specific Feedback Inhibition of Two Histidine Permeases in Saccharomyces cerewisiae
Marjolaine CRABEEL and Marcelle GRENSON
Laboratoire de Microbiologie, Facult6 des Sciences, UniversitA Libre de Bruxelles et Institut de Recherches du Centre d’Enseignement et de Recherches des Industries Alimentaires
et Chimiques, Bruxelles
(Received January 12/March 11, 1970)
Although a large fraction of the histidine accumulated from the medium remains free and intact in Xaccharomyces cerevisiae, it is not exchangeable with external histidine. Hence, the steady state concentration level of histidine is not determined by a balance between rates of inflow and outflow.
Preloading the cells with histidine results in a rapid inhibition of histidine uptake, whereas preloading with other amino acids has no effect on histidine uptake.
The inhibition by internal histidine affects the activity, and not the synthesis, of two specific histidine permeases. The histidine permease with high affinity for histidine is much more sensitive to this inhibition than the second histidine permease.
A mutant (his-p,) specifically affected in histidine uptake was isolated. Criteria for demonstrating that a mutation or a feedback control directly affect an uptake
It is concluded that the his-p, mutant is histidine-permease-less, and that the activity of the system are discussed.
histidine permeases are regulated by specific feedback inhibition.
The regulation of the intracellular concentration of metabolites like amino acids must involve a con- trol, not only of their biosynthesis, but also of the exchanges with the medium, i. e. of entry and exit of these metabolites.
The occurrence, in Saccharomyces cerevisiae, of a mechanism of regulation of histidine uptake by specific feedback inhibition has been suggested by the following observations. When protein synthesis is inhibited by cycloheximide, the rate of uptake of the amino acids tested (arginine, leucine, methionine and histidine) is almost completely inhibited within a short time, whereas when protein synthesis is prevented by histidine starvation in a histidine- requiring mutant, arginine uptake is inhibited, while the rate of histidine uptake remains constant [l]. The simplest interpretation of these facts seemed to be that the inhibitions were due to free amino acids or derivatives thereof, which accumulate as a result of the inhibition of protein synthesis.
The observations presented here support this hypothesis, and show that histidine uptake is regu- lated by specific feedback inhibition. Criteria for demonstrating that a feedback control is operating a t the level of an uptake system are discussed. A deduction of this is that some of the mutants considered as “permease-less” might be altered a t a level which is not the uptake system itself.
MATERIALS AND METHODS Organisms
The strains are all isogenic to the wild-type strain of Saccharomyces cerevisiae El278 b [2] and to its mating-tp: mutant 3962 c [3], except for the muta- tions mentioned. The strains MG389 (histidine-less), MG384 (phenylalanine-less) and MG776 (tyrosine- less) were obtained from the strain 21278b after treatment by X rays. Strain MG1216 was isolated from MG389 as growing slowly in the presence of low histidine concentrations. I ts genotype is (a) , his-, his-p,. Ethyl methanesulfonate was used as a mutagen [4]. MG1216 was crossed to 3962 c, and a prototrophic his-p, segregant (6037 a) was isolated from the re- sulting tetrads. The mutations affecting the uptake of arginine (arg-p,), of lysine (lys-p,), and of di- carboxylic amino acids (dic-p,) have been described previously [2 ,5 ,6] .
Media and Culture Methods They have been described previously [2]. Am-
medium is made of the mineral base 149, containing ammonium ions as sole source of nitrogen, and sup- plemented with vitamins and glucose.
Genetic Analysis The methods described by Hawthorne and Mor-
timer [5] were used.
198 Specific Feedback Inhibition of Histidine Uptake in Yeast Eur. J. Biochem.
Assay of Uptake Activity The method for measuring the initial velocity of
uptake of amino acids have been described previ- ously [2].
Extraction of the Soluble Pool Samples of exponentially growing cultures were
filtered onto Millipore membranes, washed rapidly with ice-cold water, and extracted for 1 O m i n with boiling water, ice-cold 5 O/,, trichloroacetic acid, or boiling 5 a/,, trichloroacetic acid.
C .- E
.E
3 E
f
- aJ I S .P
g
Fig. 1. Mean generation. time of two histidine-less strains as a function of the inverse of L-histidine concentration in the medium. Both strains bear the same histidine-less mutation. MG1216 is his-p, in addition. MG389 (0 ) ; MG1216 (0 and x :
two independent experiments)
Identification and Assay of Accumulated Histidine Two different bioassays were used for identifying
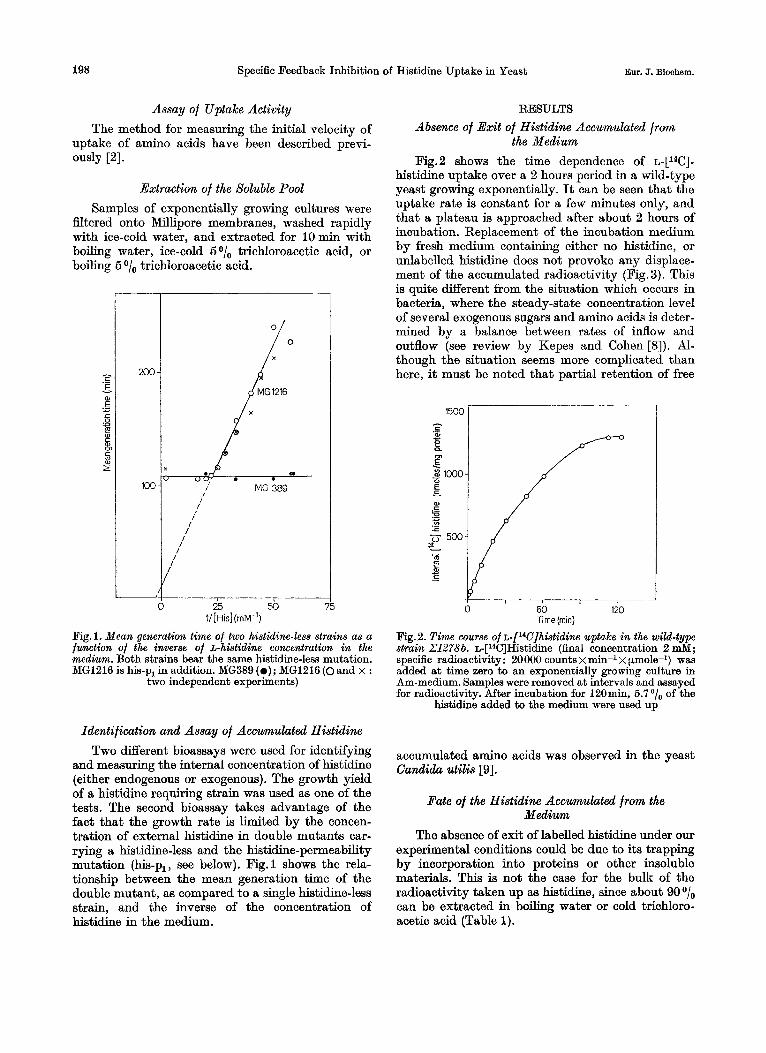
and measuring the internal concentration of histidine (either endogenous or exogenous). The growth yield of a histidine requiring strain was used as one of the tests. The second bioassay takes advantage of the fact that the growth rate is limited by the concen- tration of external histidine in double mutants car- rying a histidine-less and the histidine-permeability mutation (his-p,, see below). Fig.1 shows the rela- tionship between the mean generation time of the double mutant, a8 compared to a single histibe-less strain, and the inverse of the concentration of histidine in the medium.
RESULTS Absence of Exit of Histidine Accumulated from
the Medium Fig.2 shows the time dependence of L-[~~C]-
histidine uptake over a 2 hours period in a wild-type yeast growing exponentially. It can be seen that the uptake rate is constant for a few minutes only, and that a plateau is approached after about 2 hours of incubation. Replacement of the incubation medium by fresh medium containing either no histidine, or unlabelled histidine does not provoke any displace- ment of the accumulated radioactivity (Fig. 3). This is quite different from the situation which occurs in bacteria, where the steady-state concentration level of several exogenous sugars and amino acids is deter- mined by a balance between rates of inflow and outflow (see review by Kepes and Cohen [S]). Al- though the situation seems more complicated than here, it must be noted that partial retention of free
15001 1
0 60 120
Fig.2. Time wurse of ~-[~W]histidine uptake in the wild-type strain Z1278b. ~-[l~C]Histidine (final concentration 2 mM; specific radioactivity: 20000 counts x min-1 x &mole-1) was added at time zero to an exponentially growing culture in Am-medium. Samples were removed at intervals and assayed for radioactivity. After incubation for 120min, 5.7 of the
histidine added to the medium were used up
Time (min)
accumulated amino acids was observed in the yeast Candida utilis [9].
Fate of the Histidine Accumulated from the Medium
The absence of exit of labelled histidine under our experimental conditions could be due to its trapping by incorporation into proteins or other insoluble materials. This is not the case for the bulk of the radioactivity taken up as histidine, since about 90 O/,,
can be extracted ,in boiling water or cold trichloro- acetic acid (Table 1).
Vol. 14, No. 1,1970 M. CRABEEL and M. GRENSON 199
150 c
3 I 1
Time (rnin)
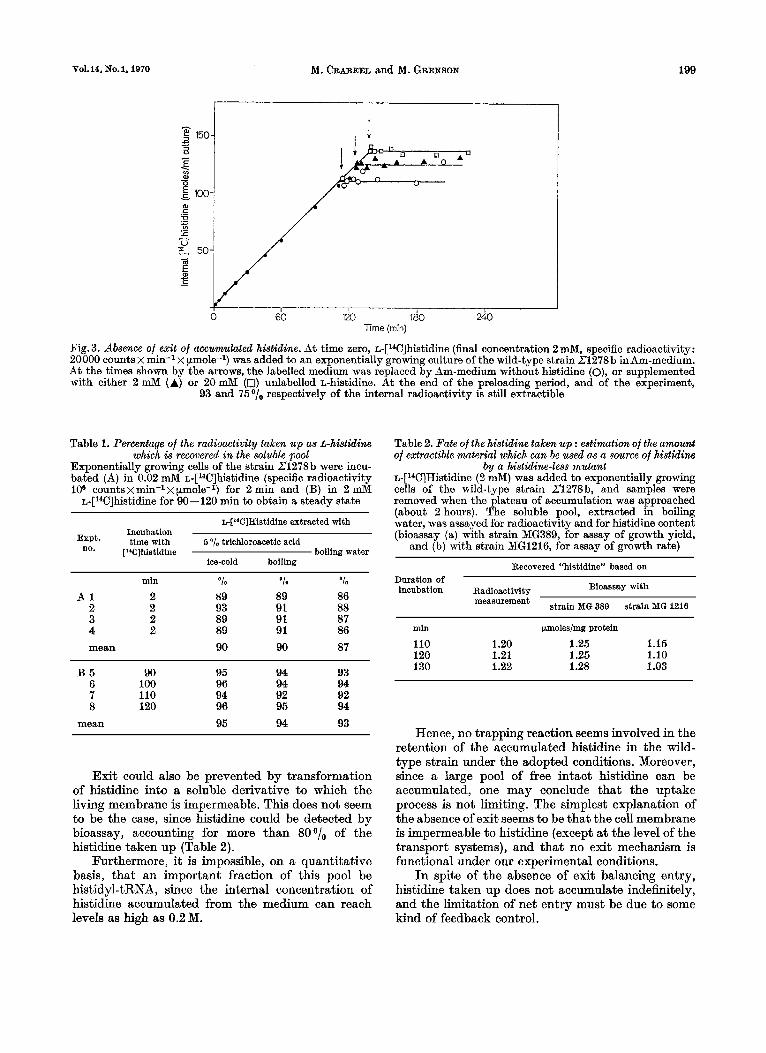
Fig. 3. Absence of exit of accumulated histidine. At time zero, ~-[l~C]histidine (final concentration 2 mM, specific radioactivity: 20000 counts X min-l X pmole-l) was added to an exponentially growing culture of the wild-type strain Z1278b in Am-medium. At the times shown by the arrows, the labelled medium was replaced by Am-medium without histidine (o), or supplemented with either 2 mM (A) or 20 mM (0) unlabelled L-histidine. At the end of the preloading period, and of the experiment,
93 and 75 o/o respectively of the internal radioactivity is still extractible
Table 1. Percentage of the radioactivity taken up as &-histidins which is recovered in the soluble pool
Exponentially growing cells of the strain 21278b were incu- bated (A) in 0.02 mM ~-[W]histidine (specific radioactivity 106 countsxmin-lx pmole-I) for 2 min and (B) in 2 mM
~-[~*C]histidme for 90-120 min to obtain a steady state
~,-['~C]Histidine extracted with Incubation time with 5 O l 0 trichloroacetic acid
ice-cold boiling ["Clhistidine boiling water
min '10 O10 '1. A 1 2 89 89 86
2 2 93 91 88 3 2 89 91 87 4 2 89 91 86 mean 90 90 87
B 5 90 95 94 93 6 100 96 94 94 7 110 94 92 92 8 120 96 95 94
mean 95 94 93
Exit could also be prevented by transformation of histidine into a soluble derivative t o which the living membrane is impermeable. This does not seem to be the case, since histidine could be detected by bioassay, accounting for more than BOO/, of the histidine taken up (Table 2).
Furthermore, it is impossible, on a quantitative basis, that an important fraction of this pool be histidyl-tRNA, since the internal concentration of histidine accumulated from the medium can reach levels as high as 0.2 M.
Table 2. Fate of the histidine taken up : estimation of the amount of extractible material which can be used as a source of histidine
by a histidine-less mutant ~-[l~C]Histidine (2 mM) was added to exponentially growing cells of the wild-type strain 2127813, and samples were removed when the plateau of accumulation was approached (about 2 hours). The soluble pool, extracted in boiling water, was assayed for radioactivity and for histidine content (bioassay (a) with strain MG389, for assay of growth yield,
and (b) with strain MG1216, for assay of growth rate)
Recovered "histidine" based on
Bioassay with Duration of incubation Radioactivity
measurement strain MG 589 strain MG 1216
min &moles/mg protein
110 1.20 1.25 1.16 120 1.21 1.25 1.10 130 1.22 1.28 1.03
Hence, no trapping reaction seems involved in the retention of the accumulated histidine in the wild- type strain under the adopted conditions. Moreover, since a large pool of free intact histidine can be accumulated, one may conclude that the uptake process is not limiting. The simplest explanation of the absence of exit seems t o be that the cell membrane is impermeable to histidine (except at the level of the transport systems), and that no exit mechanism is functional under our experimental conditions.
In spite of the absence of exit balancing entry, histidine taken up does not accumulate indefinitely, and the limitation of net entry must be due to some kind of feedback control.
200 Specific Feedback Inhibition of Histidine Uptake in Yeast Enr. J. Biochem.
6
5
4 A
3
2
1
6
5
4 A
3
2
1
0 30 60 90 120 150 Preloading time (min)
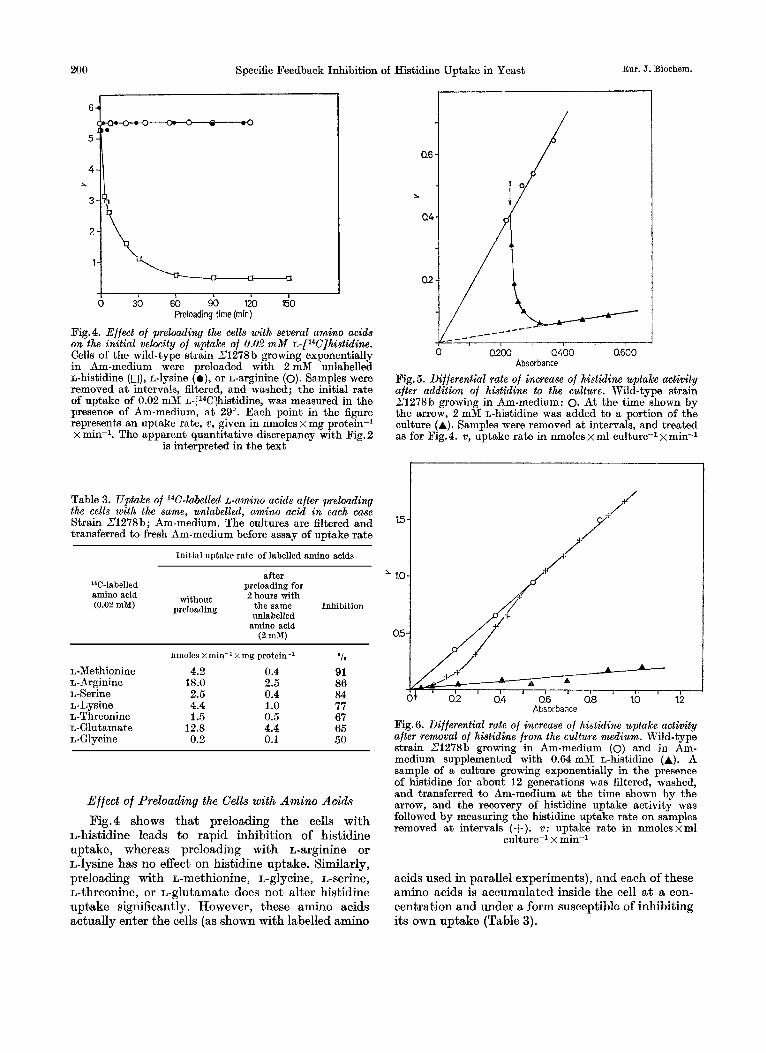
Fig.4. Effect of preloading the cells with several amino acids on the initial velocity of uptake of 0.02 m M ~-[ l~C]his t id ine . Cells of the wild-type strain 21278 b growing exponentially in Am-medium were preloaded with 2mM unlabelled L-histidine (n), L-lysine (o), or L-arginine (0). Samples were removed a t intervals, filtered, and washed; the initial rate of uptake of 0.02 mM ~-[l~C]histidine, was measured in the presence of Am-medium, at 29'. Each point in the figure represents an uptake rate, v, given in nmoles x mg protein-1 x min-l. The apparent quantitative discrepancy with Pig. 2
is interpreted in the text
Table 3. Uptake of 14C-labelled L-amino acids after preloading the cells with the same, unlabelled, amino acid in each m e Strain 21278b; Am-medium. The cultures are filtered and transferred to fresh Am-medium before assay of uptake rate
Initial uptake rate of labelled amino acids
after 14C-labelled ureloadins for
2 hours &h the same Inhibition unlabelled
without preloading
amino acid (0.02 mM)
amino acid (2 mM)
nmoles x min-' x mg protein-'
L-Methionine 4.2 0.4 L- Arginine 18.0 2.5 L-Serine 2.5 0.4 L-Lysine 4.4 1.0 L-Threonine 1.5 0.5 L-Glutamate 12.8 4.4 L-Glycine 0.2 0.1
l o 91 86 84 77 67 65 50
Effect of Preloading the Cells with Amino Acids Fig.4 shows that preloading the cells with
L-histidine leads to rapid inhibition of histidine uptake, whereas preloading with L-arginine or rJ-lysine has no effect on histidine uptake. Similarly, preloading with L-methionine, L-glycine, L-serine, L-threonine, or L-glutamate does not alter histidine uptake significantly. However, these amino acids actually enter the cells (as shown with labelled amino
Absorbance Fig. 5. Differential rate of increase of histidine uptake activity after addition of histidine to the culture. Wild-type strain Z1278b growing in Am-medium: 0. At the time shown by the arrow, 2 mM L-histidine was added to a portion of the culture (A). Samples were removed a t intervals, and treated as for Fig. 4. v, uptake rate in nmoles x ml culture-1 x min-1
1.5 i 1.0-
A A
. A A 1 1 1 1 1 1 1 1 1 , ,
1.0 1.2 Absorbance
Fig. 6. Differential rate of increase of histidine uptake activity after removal of histidine from the culture medium. Wild-type strain Z1278b growing in Am-medium (0) and in Am- medium supplemented with 0.64 mM L-histidine (A). A sample of a culture growing exponentially in the presence of histidine for about 12 generations waa filtered, washed, and transferred to Am-medium at the time shown by the arrow, and the recovery of histidine uptake activity was followed by measuring the histidine uptake rate on samples removed at intervals (f). v: uptake rate in nmolesxml
culture-l x min-1
acids used in parallel experiments), and each of these amino acids is accumulated inside the cell a t a con- centration and under a form susceptible of inhibiting its own uptake (Table 3).
Vol. 14, No. 1, 1970 &I. CRABEEL and M. GRENSON 201
Figs. 5 and 6, using the graphic method ofMonod, Pappenheimer and Cohen-Bazire [lo], show that the inhibition observed after preloading the cells with histidine affects the activity, and not the synthesis, of the uptake system. This method of distinguishing a modification of activity from a modification of differential rate of synthesis in uptake systems was used in the case of dicarboxylic amino acids uptake in yeast, where a repression phenomenon was ob- served in one strain and an inhibition of activity in another strain [6], and in the case of several uptake systems of aliphatic amino acids which appear as inducible [22]. The fact that in the present case the recovery of activity after elimination of histidine from the medium is rather slow (Fig.6) can be ex- plained by the dependence of the removal of the accumulated inhibitor on protein synthesis. This is consistent with the fact that residual growth after histidine starvation of a histidine-less mutant is about two doublings.
These data clearly show that there is a specific feedback inhibition on the process of histidine uptake, and indicate that the uptake of other amino acids is probably regulated in the same way.
A Mutant Specifically Affected in Histidine Uptake
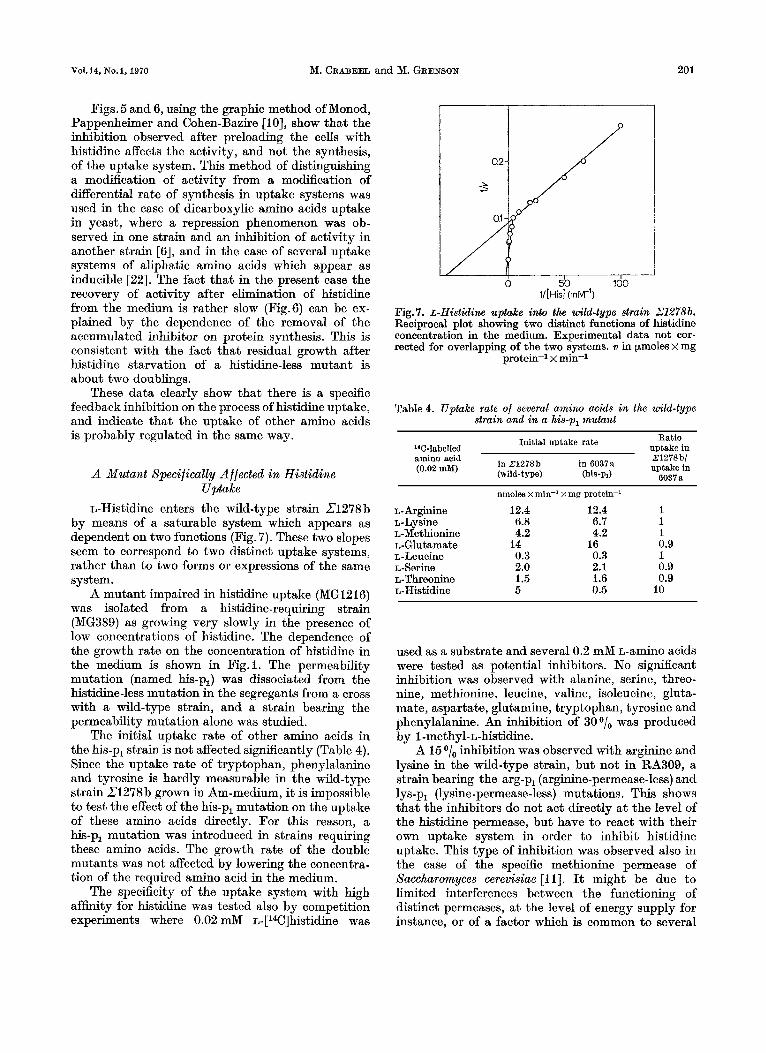
L-Histidine enters the wild-type strain Z1278 b by means of a saturable system which appears as dependent on two functions (Fig. 7). These two slopes seem to correspond to two distinct uptake systems, rather than to two forms or expressions of the same system.
A mutant impaired in histidine uptake (MGl216) was isolated from a histidine-requiring strain (MG389) as growing very slowly in the presence of low concentrations of histidine. The dependence of the growth rate on the concentration of histidine in the medium is shown in Fig.l. The permeability mutation (named his-p,) was dissociated from the histidine-less mutation in the segregants from a cross with a wild-type strain, and a strain bearing the permeability mutation alone was studied.
The initial uptake rate of other amino acids in the his-p, strain is not affected significantly (Table 4). Since the uptake rate of tryptophan, phenylalanine and tyrosine is hardly measurable in the wild-type strain 21278b grown in Am-medium, it is impossible to test the effect of the his-p, mutation on the uptake of these amino acids directly. For this reason, a his-p, mutation was introduced in strains requiring these amino acids. The growth rate of the double mutants was not affected by lowering the concentra- tion of the required amino acid in the medium.
The specificity of the uptake system with high affinity for histidine was tested also by competition experiments where 0.02 mM ~-[l~C]histidine was
Fig.7. .&-Histidine uptake into the wild-type strain Z1278b. Reciprocal plot showing two distinct functions of histidine concentration in the medium. Experimental data not cor- rected for overlapping of the two systems. v in pmoles X mg
protein-1 x min-1
Table 4. Uptake rate of several amino acids in the wild-type strain and in a his-pl mutant
Ratio uptake in Z1278b/ uptake in
Initial uptake rate
in H1278b in 6037 a (wild-type)
14C-labelIed amino acid (0.02 mM)
(his-pd 6037a
L- Arginine L-Lysine L-Methionine L-Glutamate L-Leucine L-Serine L-Threonine L-Histidine
nmoles x min-' x mg protein-'
12.4 6.8 4.2
0.3 2.0 1.5 5
14
12.4 1 6.7 1 4.2 1
16 0.9 0.3 1 2.1 0.9 1.6 0.9 0.5 I0
used as a substrate and several 0.2 mM L-amino acids were tested as potential inhibitors. No significant inhibition was observed with alanine, serine, threo- nine, methionine, leucine, valine, isoleucine, gluta- mate, aspartate, glutamine, tryptophan, tyrosine and phenylalanine. An inhibition of 300/, was produced by 1 -methyl-L-histidine.
A 15 O/,, inhibition was observed with arginine and lysine in the wild-type strain, but not in RA309, a strain bearing the arg-p, (arginine-permease-less) and lys-p, (lysine-permease-less) mutations. This shows that the inhibitors do not act directly a t the level of the histidine permease, but have to react with their own uptake system in order to inhibit histidine uptake. This type of inhibition was observed also in the case of the specific methionine permease of Saccharomyces cerevisiae [ l l ] . It might be due to limited interferences between the functioning of distinct permeases, a t the level of energy supply for instance, or of a factor which is common to several

Eur. J. Biochem. 202 Specific Feedback Inhibition of Histidine Uptake in Yeast
transport systems and limited in amount, as sug- gested by Winkler and Wilson [12].
The characteristics of the his-p, mutant and competition experiments using a low histidine con- centration as substrate show that the uptake system with high affinity for histidine is very specific.
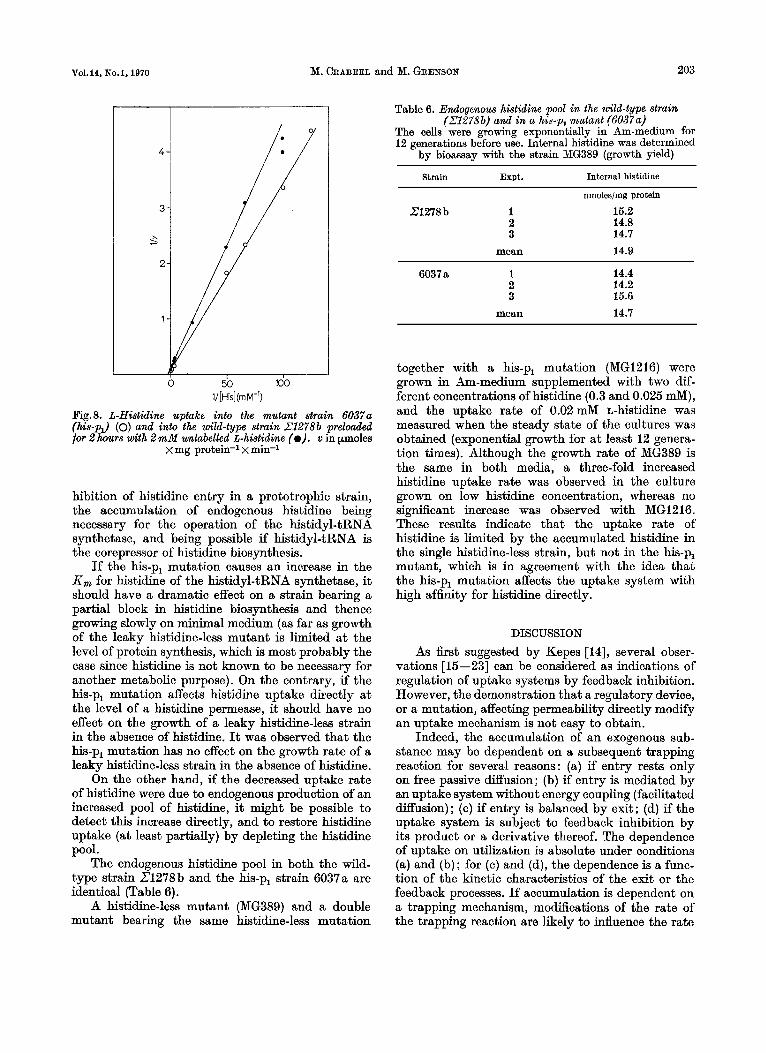
The kinetic characteristics of histidine uptake in the his-p, mutant are modified as shown in Fig.8. The uptake function with high affinity for histidine is no longer detectable.
The residual histidine uptake activity in the his-p, mutant seems specific too, since it is not inhibited significantly by a number of L-amino acids (0.5 mM; the same amino acids as those used for the preceding competition test ; ~-[l~C]histidine concen- tration was 0.05 mM), including phenylalanine and tryptophan. Significant inhibition was observed only with tyrosine (3Oo/J and with glutamate (60°/,). Since the inhibition by tyrosine appears as com- petitive, it is not excluded that the uptake system with low affinity for histidine might be able to trans- port tyrosine too. Since no inhibition was observed with phenylalanine or tryptophan, the specificity of this permease seems different from that of the aroma- tic amino acid permease of Salmonella typhimurium [13]. The inhibition by glutamate of the uptake ac- tivity with low affinity for histidine is by no means an indication of a lack of specificity of this uptake system, since the inhibition by glutamate is not com- petitive and only partial, and that it is absent in a mutant impaired (directly or indirectly?) in the dicarboxylic amino acids uptake activity. This indirect type of inhibition may help to distinguish separate systems with similar specificity: the in- hibition of one of the two histidine uptake functions by arginine and lysine, and of the other function by glutamate suggests that these functions are linked to two distinct permeases.
Sensitivity of the Histidine Permeases to Feedback Inhi bition
As shown in Table 5, the permease with high af- finity for histidine seems much more sensitive to feedback inhibition than the uptake system with low affinity for histidine. This is also apparent when Fig.2 is compared to Fig.4. Fig.2 shows progressive inhibition of the uptake of 2 mM histidine which establishes much more slowly than inhibition of the uptake of 0.02mM [14C]histidine (Fig. 4) by preloading with 2 mM unlabelled histidine. Indeed, considering the kinetic characteristics of the two histidine uptake systems, it is clear that histidine enters the cell essentially by the high affinity system a t a 0.02 mM histidine concentration, whereas in the presence of a 2 mM external histidine concentration, the low af- finity system is predominant. After preloading the cells with 2 mM L-histidine for 120 min, the kinetic properties are modified (Fig. 8) : the function with high affinity for histidine is no longer detectable, whereas the other function has kept more than 70 O l 0 of its activity.
I s his-p, a True Permease-less Mutation? The observations above are compatible with two
different interpretations of the mutation : (a) it might prevent the formation of a functional histidine permease, or (b) the mutation might act indirectly on histidine uptake by creating conditions of feedback inhibition of the histidine permease.
For instance, all the properties of the mutant could be explained if the mutation affects the histidyl-tRNA synthetase in such a way that its apparent affinity constant (K,) for histidine is in- creased. The consequences ofthis would be; (a) a slow growth of an histidine-less mutant on low external concentrations of histidine, and (b) the feedback in-
Table 5. Sensitivity of the histidine permeases to feedback inhibition The cells were incubated for 2 hours in Am-medium supplemented with the indicated concentrations of unlabelled L-histidine before the assays. The internal concentration of histidine was estimated by radioactivity measurements (see Table 2), except
for the pool at time zero which was determined by bioassay with the strain MG389
High afflnity uptake system, strain 127Sb Low affinity uptake system, strain 6037a Histidine conc. for Initial uptake rate Initial uptake rate
preloading of ["Clhistidine Inhibition Pool size of ["Clhistidine Inhibition Pool size (0.02 mH) (0.1 mM)
mM nmoles x min-I x mg protein-' nmoleslmg protein nmoles x min-' xmg protein-' nmoleslmg protein
0 5.20 - 15 2.88 - 15 2 0.53 90 1392 2.03 30 1349 5 0.35 93 1768 1.92 33 1667
10 0.44 91 - 1.76 39 1772 15 0.45 91 2041 1.73 40 1914 20 0.45 91 2095 1.68 42 1940
Vol. 14, No. 1,1970 M. CRABEEL and M. GRENSON 203
4-
I 0 50 100
’ 7
i
l/[HL](mM-’)
Fig.8. L-Histidine uptake into the mutant strain 6037a (his-pJ (0) and into the wild-type strain 21278b preloaded for 2 hours with 2 m M unlabelled L-histidine (0). v in pmoles
x mg protein-l x min-l
hibition of histidine entry in a prototrophic strain, the accumulation of endogenous histidine being necessary for the operation of the histidyl-tRNA synthetase, and being possible if histidyl-tRNA is the corepressor of histidine biosynthesis.
If the his-p, mutation causes an increase in the K , for histidine of the histidyl-tRNA synthetase, it should have a dramatic effect on a strain bearing a partial block in histidine biosynthesis and thence growing slowly on minimal medium (as far as growth of the leaky histidine-less mutant is limited a t the level of protein synthesis, which is most probably the case since histidine is not known to be necessary for another metabolic purpose). On the contrary, if the his-p, mutation affects histidine uptake directly at the level of a histidine permease, it should have no effect on the growth of a leaky histidine-less strain in the absence of histidine. It was observed that the his-p, mutation has no effect on the growth rate of a leaky histidine-less strain in the absence of histidine.
On the other hand, if the decreased uptake rate of histidine were due to endogenous production of an increased pool of histidine, it might be possible to detect this increase directly, and to restore histidine uptake (at least partially) by depleting the histidine
The endogenous histidine pool in both the wild- type strain Z1278b and the his-p, strain 6037a are identical (Table 6).
A histidine-less mutant (MG389) and a double mutant bearing the same histidine-less mutation
pool.
9’
/
I 0 50 100
Table 6. Endogenous histidine pool in the wild-type strain (ZlZ78b) and in a his-p, mutant ( 6 0 3 7 ~ )
The cells were growing exponentially in Am-medium for 12 generations before use. Internal histidine was determined
by bioassay with the strain MG389 (growth yield)
Expt. Internal histidine Strain
nmoleslmg protein
Z1278b 1 15.2 2 14.8 3 14.7
mean 14.9
6037 a 1 2 3
14.4 14.2 15.6
mean 14.7
together with a his-p, mutation (MG1216) were grown in Am-medium supplemented with two dif- ferent concentrations of histidine (0.3 and 0.025 mM), and the uptake rate of 0.02mM L-histidine was measured when the steady state of the cultures was obtained (exponential growth for a t least 12 genera- tion times). Although the growth rate of MG389 is the same in both media, a three-fold increased histidine uptake rate was observed in the culture grown on low histidine concentration, whereas no significant increase was observed with MG1216. These results indicate that the uptake rate of histidine is limited by the accumulated histidine in the single histidine-less strain, but not in the his-p, mutant, which is in agreement with the idea that the his-p, mutation affects the uptake system with high affinity for histidine directly.
DISCUSSION As -first suggested by Kepes [la], several obser-
vations [15-231 can be considered as indications of regulation of uptake systems by feedback inhibition. However, the demonstration that a regulatory device, or a mutation, affecting permeability directly modify an uptake mechanism is not easy to obtain.
Indeed, the accumulation of an exogenous sub- stance may be dependent on a subsequent trapping reaction for several reasons: (a) if entry rests only on free passive diffusion; (b) if entry is mediated by an uptake system without energy coupling (facilitated diffusion); (c) if entry is balanced by exit; (d) if the uptake system is subject to feedback inhibition by its product or a derivative thereof. The dependence of uptake on utilization is absolute under conditions (a) and (b) ; for (c) and (d), the dependence is a func- tion of the kinetic characteristics of the exit or the feedback processes. If accumulation is dependent on a trapping mechanism, modifications of the rate of the trapping reaction are likely to influence the rate

204 M. CRABEEL and M. GRENSON: Specific Feedback Inhibition of Histidine Uptake in Yeast Eur. J. Biochem.
of uptake, for instance if the trapping reaction itself is inhibited by feedback regulation, or if it is not operating as a result of a mutation. Such a situation is illustrated in the case of the uptake of uracil and uridine in yeast [24], where two types of mutations were shown to alter uracil and uridine uptake, one type a t the uptake level and the other a t the level of the first utilizing enzyme. The effect on perme- ability of this second class of mutation is due to feedback inhibition on the corresponding permeases, since uptake is restored by pyrimidine starvation in this type of mutants, whereas it is not in uracil or uridine-permease-less mutants.
Inhibition of uptake after preloading the cells with the unlabelled substrate seems a better indication of regulation of net uptake by the accumulated sub- stance or a derivative. However, this inhibition might be due to other effects, such as repression of the synthesis of the uptake system, induction of an exit mechanism, induced destruction of the uptake system, etc.
We think that the criteria used in the present work show, with a rather high degree of confidence, that there is a control on histidine uptake by feedback inhibition a t the level of the histidine permeases in yeast. First, histidine is transported by an accumu- lating and specific uptake system. No exit of the accumulated substrate or a derivative thereof occurs under our experimental conditions. An inhibition of histidine uptake is observed after preloading the cells with histidine only. Hence, the specificity of the preloading effect is the same as that of the permease, which is a condition for this inhibition to act as a regulatory mechanism. Finally, the observed inhibi- tion from inside the cell affects the activity of the histidine uptake system and not its synthesis.
About the mechanism of this inhibition, practi- cally nothing is known. Since inhibition shows the same type of specificity as the permease, it is reason- able to think that it acts a t the level of the specific element of the uptake system, i. e. the permease, as far as it is the only specific step of the transport mechanism. At this level, two possible mechanisms must be considered. The receptor of the inhibitory signal might be a specialized site comparable to those of allosteric enzymes. But, on the other hand, inhibi- tion might be the result of the combination of one of the products of the reaction (internal histidine 2 ) with the site which liberates it inside the cell, once the translocation and energy coupled reaction have occurred.
I n order to stop net inflow of exogenous meta- bolites, and hence prevent excessive accumulation, living cells seem to use two different mechanisms: exit balancing entry, and feedback inhibition of entry. From the point of view of cell economy, feed- back inhibition appears as the best system. The utilization of one or the other of these mechanisms
2.
3.
4.
5. 6.
7.
8.
9.
10.
11.
12.
13. 14.
15. 16.
17.
18.
19. 20.
21.
22.
23.
is most probably not exclusive, and the difference might be a question of predominance of one of the systems on the other. Indeed, feedback inhibition appears as best suited to control the accumulation of a substance from the outside, whereas exit is able to prevent the formation of excessive concentrations of endogenous products. The colIaboration of these two mechanisms in the regulation of the intracellular pool of uracil and uridine has been demonstrated in yeast [24].
The authors are grateful to Mrs. F. Muyldermans- Verhaegen for her skilful technical assistance. This work was supported by a grant from the Fonds de la Recherche Fondamentale Collective. M. C. holds a prcdoctoral fellow- ship from the I. R. S. I. A.
REFERENCES 1. Grenson, M., Crabeel, M., Wiame, J. M., and BBchet, J.,
Biochem. Bioahws. Res. Commun. 30 (1968) 414. Grenson, M., MOu&et, M., Wiame, J. M.', andBbchet, J.,
Biochim. Biophys. Acta, 127 (1966) 325. BBchet, J., Grenson, M., and Wiame, J. M., Eur. J .
Biochem. 12 (1970) 31. Lindegren, G., Hwang, Y. L., Oshima, Y., and Linde-
gren, C. C., Can. J . Genet. Cytol. 7 (1965) 491. Grenson, M., Biochim. Biophys. Acta, 127 (1966) 339. Joiris, C. R., and Grenson, M., Arch. Intern. Physiol.
Biochem. 77 (1969) 154. Hawthorne, D. C., and Mortimer, R. K., Genetics, 45
(1960) 1085. Kepes, A., and Cohen, G. N., In The Bacteria (edited by
Gunsalus and Stanier), Academic Press, New Pork 1962, Vol. IV, p. 179.
Cowie, D. B., and McClure, F. T., Biochim. Biophys. Acta, 31 (1959) 236.
Monod, J., Pappenheimer, A. M., and Cohen-Bazire, G., Biochim. Biopl~ys. A d a , 9 (1952) 648.
Gits, J. J., and Grenson, M., Biochim. Biophys. Acta, 135 (1967) 507.
Winkler, H. H., and Wilson, T. H., Biochim. Biophys. Acta, 135 (1967) 1030.
Ferroluzzi-Ames, G., Arch. Biochem. 104 (1964) 1. Kepes, A., In The Cellular Functions of Membrane
Transport (edited by Hoffman), Prentice-Hall, Engle- wood Cliffs 1964.
Scott, J. M., and Spencer, B., Biochem. J . 96 (1965) 78P. Dreyfuss, J., and Pardee, A. B., J . Bacteriol. 91 (1966)
2275. Yamamoto, L. A., and Segel, I. H., Arch. Biochem.
Biophys. 114 (1966) 523. VallBe, M., and Jeanjean, R., Bioc?kim. Biophys. A d a ,
150 (1968) 599. Ring, K., and Heinz, E., Biochem. 2. 344 (1966) 446. Ring, K., Gross, W., and Heinz, E., Berichte der Bunsen-
gesellschaft fur physikalische Chemie, 71 (1967) 893. Bedko, P. V., Wood, T. C., and Segel, I. H., Arch. Bio-
chem. Biophys. 122 (1967) 783. Wilev. W. R., and Matchett, W. H., J . Bacteriol. 95
(1968) 959. Gits. J. J., and Grenson, M., Arch. Intern. Physiol. Bio-
chem. 77 (1969) 153. 24. Grenson, M., Eur. J. Biochem. 11 (1969) 249.
M. Crabeel et M. Grenson Institut de Recherche5 du C. E. R. I. A. Avenue Emile Gryzon 1, B-1070 Bruxelles, Belgium










![Control of Vascular Sap pH by the Vessel-Associated Cells ... · of Poitiers University. Perfusion Technique This technique (initially used by Van Bel [1974] to study the uptake of](https://img.pdfslide.fr/doc/110x75/5eda89c0febf237c0c3b770d/control-of-vascular-sap-ph-by-the-vessel-associated-cells-of-poitiers-university.jpg)









![Antisenserepression chloroplast inProc. Natl. Acad. Sci. USA90(1993) 6161 centrifugation. The transport of [14C]malate was used to normalize [32P]P, uptake activities of different](https://img.pdfslide.fr/doc/110x75/5ffa530e20ae570318129513/antisenserepression-chloroplast-in-proc-natl-acad-sci-usa901993-6161-centrifugation.jpg)








