Embed Size (px)
Citation preview

Eur. J . Biochem. 74, 567-574 (1977)
Role of Divalent Ions in Folding of tRNA Jean-Louis LEROY, Maurice GUERON, Gilles THOMAS, and Alain FAVRE
Groupe de Biophysique du Laboratoire de Physique de la Matiere Condensee (Equipe de Recherches du Centre National de la Recherche Scientifique), Ecole Polytechnique, Palaiseau, and lnstitut de Recherche en Biologie Moleculaire, Universite de Paris VII
(Received November 26, 1976)
The native structure of tRNA is not achieved in low salt (4.5 mM Na', 25 "C), but can be restored by addition of divalent ions. We have explored the structure of the central region in Escherichiu coli tRNAfM"' by absorption and emission spectroscopy of Cthiouracil, and the structure of the anticodon loop in yeast tRNAPh' by fluorescence of the 'Y' base, versus the number of manganese ions bound to tRNA, which was derived from electron spin resonance. The fluo- rescence of the reduced 8- 13 photoproduct (in which 4-thiouracil at position 8 is crosslinked to cytosine at position 13) was also analysed.
In low salt (e .g . 4.5 mM Na'), the region of 4-thiouracil is affected strongly as the first eight Mn2+ bind to tRNA, whereas the fluorescence of the 'Y' base is affected only after four Mn2' are bound. Considering the structural similarities of the two tRNAs, this suggests that the re- organisation brought about by divalent ions starts in the central region, the anticodon loop being affected later. The binding of divalent ions to each region starts together with its restructuration.
Monovalent ions can substitute for divalent ions in this process, a 15 mM sodium concentration being equivalent to the binding of the first five Mn2+. If divalent ions are then added, even the first ones distribute themselves between both the central and the anticodon region. Alternatively, the renaturation may be achieved by monovalent ions only, implying that no sites exist whose occupancy by divalent ions is crucial for the native structure. These observations suggest that the role and means of divalent ion binding to tRNA are largely explainable in terms of a simple maganese-phosphate binding supplemented by electrostatic interaction with distant phosphates.
As polyions, the polynucleotides exhibit structures in solution which are strongly dependent on the con- centration and valence of cations. Typically, the com- pact native structures of biological polynucleotides are favoured by high salt concentrations and particu- larly by divalent ions, whereas in low salt, unfolded forms appear.
Transfer RNAs are among the best known and most studied nucleic acids but even in their case, and despite analysis of the:)(-ray structures [l , 21, the role and location of bound divalent cations is not clear.
Closely related is the question of their inter- changeability with monovalent cations. Similar struc-
A preliminary report was presented at the 10th meeting of the Federation of European Biochemical Societies, Paris, July 20- 25, 1975 (J. L. Leroy, G.Thomas, M. Gueron & A. Favre, Abstract 382).
Ahhreviution. ESR, electron spin resonance. Trivial Nunzes. 8- 13 photoadduct, the product formed by ir-
radiating E. coli tRNAfM" such that C-4 of residue 8 (thiourdcil) is linked to C-5 of residue 13 (cytosine); reduction of this photo- adduct adds an H to C-6 of cytosine (see Fig. l a ) .
tural alterations may be produced by varying concen- trations of monovalent and divalent ions, or the temperature.
Low-salt conditions can be found in which the native structure is not completely formed, as seen, for instance from optical emission properties of rare bases, whereas most of the stacking interactions are already present, as judged from hypochromicity at 260 nm. The addition of divalent ions restores the native structure. It is then important to try to follow the steps of this restoration, i.e. to characterize the various structural changes as a function of the number of bound ions, and to identify the binding sites.
The present work is a contribution to this problem. We chose manganese as the divalent ion because the number of bound ions may be derived conveniently by electron spin resonance (ESR) [3,4]. It is known that Mn2+ substitutes well for Mg2+ in the reactions of protein synthesis [ 5 ] .
We used the following well-known local probes of tRNA structure: in Escherichia coli tRNArM"', we
568 Divalent Ions in tRNA
.c - C
G-C-U- C, ! G
G \
20 7' u
I s 4 u
G - A , '"\ ,.' C* G -A- G . . . .
F. 30-9
F .G
I p
A E
. . . . . ,G;U- C- G-G ,
C T
m G * A 50 >
I
yo 'A
E
- A
A I
- Y
I
A
c-u- c-&' A. D-G*
D' I . . . . G G-A-G - C. \ G .,G - A # m$G
{ -70 U
60 I t
A i c-u. ' G-A-C- A-C' m'k
6 . . . . . &c-u-G-u-G, C'
u- 50 m7G
G' A*
/ $ 4 20 C . 9
I I
B
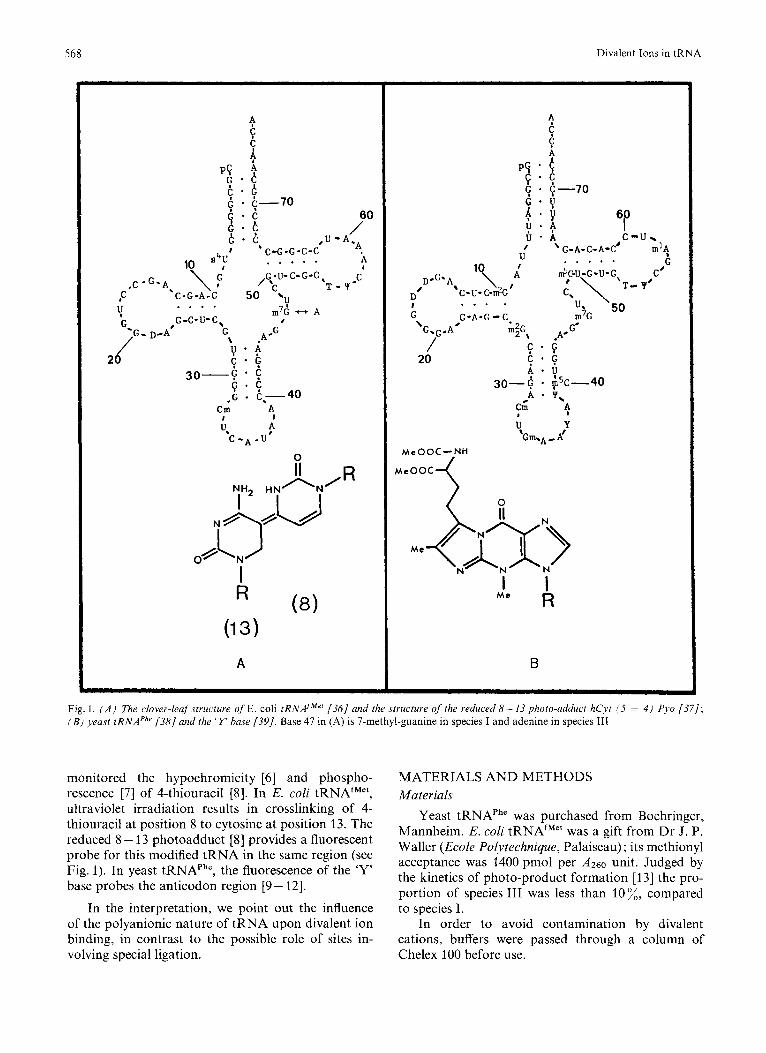
Fig. 1. ( A ) The clover-leaf structure of E. coli tRNAiMe' (361 and the structure of the reduced 8- 13 photo-aclduct hCji (5 = 4 ) Pyo (371; ( B ) yeast tRNAPhr (381 and the ' Y base (391. Base 47 in (A) is 7-methyl-guanine in species I and adenine in species 111
monitored the hypochromicity [6] and phospho- rescence [7] of 4-thiouracil [8]. In E. coli tRNAfMet, ultraviolet irradiation results in crosslinking of 4- thiouracil at position 8 to cytosine at position 13. The reduced 8 - 13 photoadduct [S] provides a fluorescent probe for this modified tRNA in the same region (see Fig. 1). In yeast tRNAPh', the fluorescence of the 'Y' base probes the anticodon region [9- 121.
In the interpretation, we point out the influence of the polyanionic nature of tRNA upon divalent ion binding, in contrast to the possible role of sites in- volving special ligation.
MATERIALS AND METHODS Materials
Yeast tRNAPhe was purchased from Boehringer, Mannheim. E. coli tRNAfMet was a gift from Dr J. P. Waller (Ecole Polytechnique, Palaiseau) ; its methionyl acceptance was 1400 pmol per unit. Judged by the kinetics of photo-product formation [13] the pro- portion of species I11 was less than lo"/,, compared to species I.
In order to avoid contamination by divalent cations, buffers were passed through a column of Chelex 100 before use.
J.-L. Leroy, M. Gueron, G . Thomas, and A. Favre 569
Preparation of Samples
To remove divalent cations, tRNA (50 ,4260 units in 2 ml) was heated for 5 min at 60"C, then dialysed successively at 6°C for at least 8 h, against 1 1 of the following buffers: first 1 M NaCl, 20 mM EDTA; then 1 M NaC1, 5 mM sodium cacodylate buffer with beads of Chelex 100; then (twice) 5 mM cacodylate buffer with Chelex beads. All solutions were at pH 7.0. The free sodium contributed by the cacodylate buffer amounts to 4.2 mM and is included in the sodium concentrations given. An analysis by spark spectro- scopy shows that the total concentration of extra- neous metals (mainly Ca) is less than 0.1 atom per tRNA molecule.
Mn2 Titrations
The X-band ESR spectrum of Mn2+ bound to tRNA is at least four times broader [3] than that of free Mn2+ (80 G vs 20 G peak-to-peak derivative). In the derivative presentation which is commonly used in ESR, this means that the peak height of the bound species is smaller than the free species by a factor of (4)2 = 16. For sufficiently low tRNA con- centrations, the 'bound' contribution may then be neglected, so that the ESR peak height measures the 'free' concentration. When necessary the small 'bound' contribution has been corrected for. The 'bound' fraction is obtained as 'total' minus 'free'.
Spectral Measurements
2°C on a Cameca TSN 258 spetrometer, using 200 p1 of a solution containing from 1.6 to 24 A260 units/ml of tRNA for the low-salt and high-salt (2 50 mM) samples respectively. The sensitivity was 0.3 pM free Mn2+. The tRNA concentrations were calculated assuming that 1 ,4260 unit/ml corresponds to 1.8 pM of tRNA in 5 mM cacodylate buffer, pH 7.
Fluorescence was observed with a Jobin-Yvon fluorimeter. For the 'Y' base the excitation and emis- sion wavelengths were A,,, = 320 nm, Aem = 440 nm, and for the reduced 8 - 13 photo-adduct A,,, = 390 nm, I,,, = 450 nm. The tRNA concentration was 1.6 ,4260
units/ml, so that the sample absorption in the 2-mm- thick cell was negligible at the wavelengths of interest. The fluorescence intensity of the 'Y' base was trans- formed into quantum yield by comparison to the value (7%) of Eisinger et al. [9] for tRNA in high Mg2+. The fluorescence quantum yield of the 8 - 13 photo-adduct was obtained by reference to the value of Favre and Yaniv [8].
Phosphorescence of 4-thiouracil in tRNA was measured at room temperature. It forms a band with a peak at 510 nm, and is superimposed on the
ESR measurements were carried out at 25
red flank of another peak, at 400nm. In E.co1i tRNAfMe', the lifetime of the 510-nm band is 3.5 ps (Favre et ul., unpublished). We therefore feel justified in describing this emission as phosphorescence [7,14, 151, but one should be aware that other authors have considered it to be a fluorescence with a lifetime in the nanosecond range [16,17]. The phosphorescence spectra were obtained on a Jobin-Yvon 'Bearn' fluori- meter, the excitation wavelength being 335 nm. The emission spectrum was decomposed into its 440-nm and 510-nm components by inspection.
Formation of Photo-Adduct
E. coli tRNAfMe' at a concentration of 40 A260
units/ml in 50 mM NaC1, 50 mM sodium cacodylate, 10 mM MgCL, pH 7, was irradiated with mono- chromatic light at 365 nm as already described [18]. The reduction of the photo-adduct by borohydride [8] was facilitated by omitting Mg" from the reaction mixture.
RESULTS
Existence of Structure in Low Sall
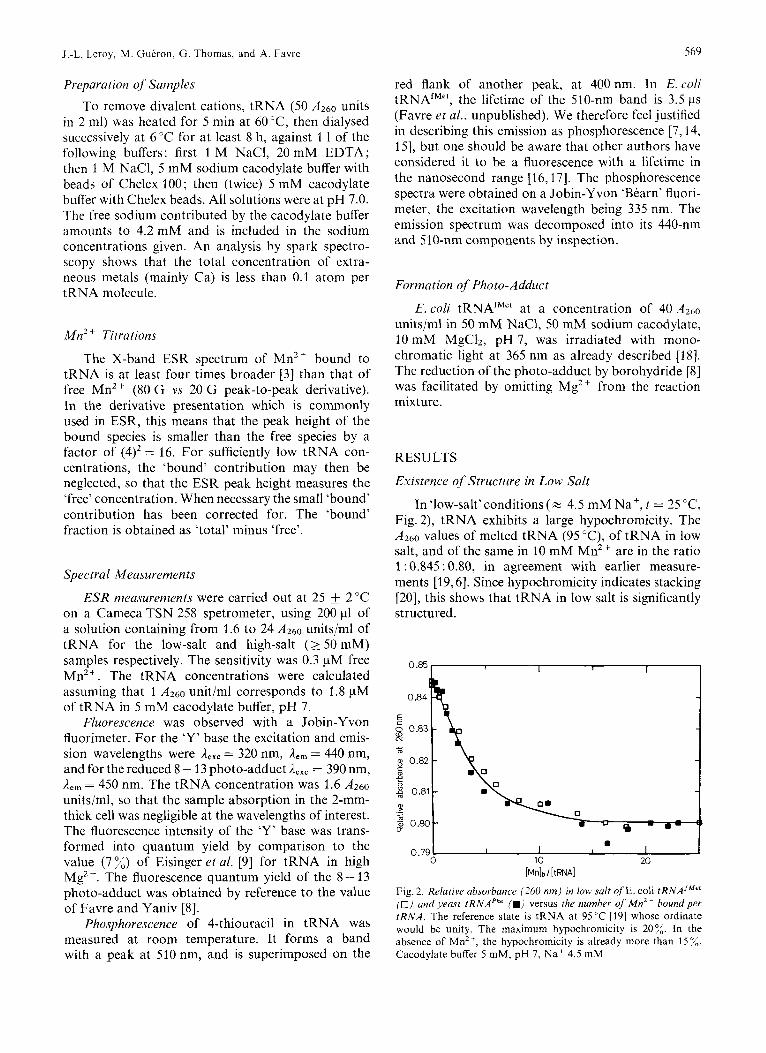
In 'low-salt' conditions (% 4.5 mM Na', t = 25 "C, Fig. 2), tRNA exhibits a large hypochromicity. The ,4260 values of melted tRNA (95 "C), of tRNA in low salt, and of the same in 10 mM Mn2' are in the ratio 1 : 0.845 : 0.80, in agreement with earlier measure- ments [19,6]. Since hypochromicity indicates stacking [20], this shows that tRNA in low salt is significantly structured.
0 .a5 I I
0.79 I 1 I I 0 10 20
[Mn]b/[tRNA]
Fig. 2. Relative absorbance (260 nm) in low, salr of' E. coli tRNAJM" (0) and yeast tRNAPhe (H) versus the number of Mn2+ bound per tRNA. The reference state is tRNA at 95°C [19] whose ordinate would be unity. The maximum hypochromicity is 20%. In the absence of MnZ+, the hypochromicity is already more than 15 7". Cacodylate buffer 5 mM, pH 7, Na' 4.5 mM
I
570 Divalent Ions in tRNA
I 1 I I
0.8
0 10 20 [Mn]b/[tRNA]
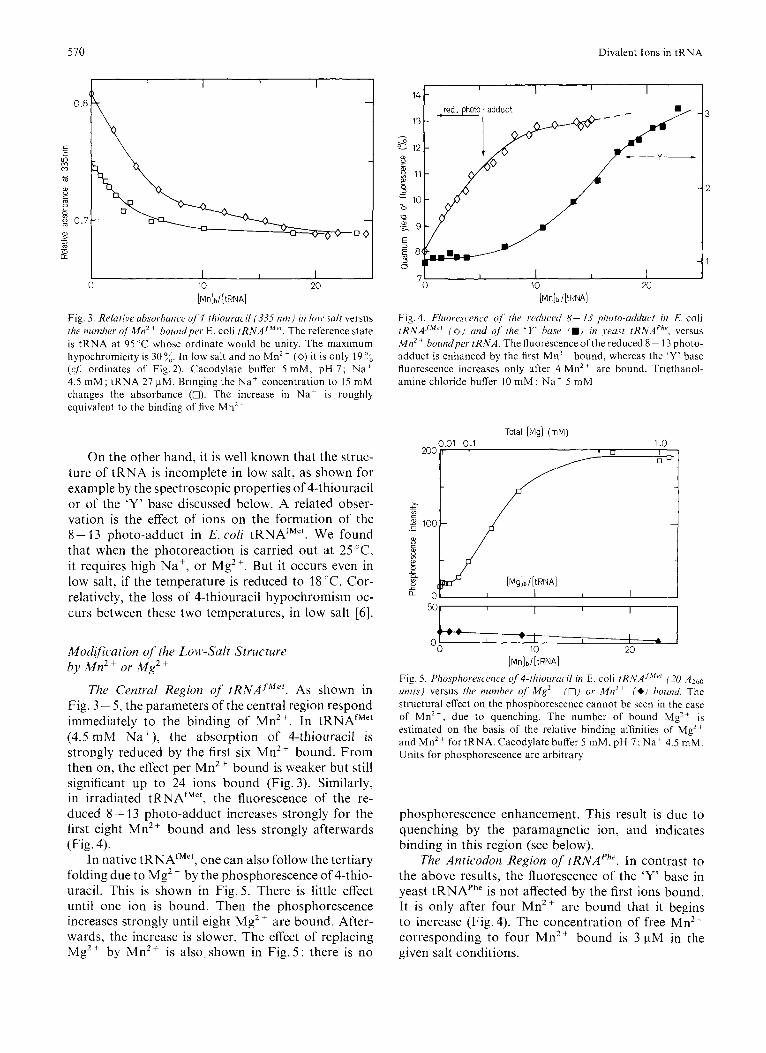
Fig. 3. Relutive ubsorbunce of 4-thiouruci~ (335 n m ) in Ion. s d r versus the number q f M n z + boundyer E. coli tRNAJ"". The reference state is tRNA at 95 ' C whose ordinate would be unity. The maximum hypochromicity is 30%. In low salt and no Mn2+ (0) it is only 19°C (q f : ordinates of Fig.2). Cacodylate buffer 5 mM, pH 7 ; N a + 4.5 m M ; tRNA 27 pM. Bringing the Na' concentration to 15 mM changes the absorbance (0). The increase in Na' is roughly equivalent to the binding of five M n Z +
On the other hand, it is well known that the struc- ture of tRNA is incomplete in low salt, as shown for example by the spectroscopic properties of 4-thiouracil or of the 'Y' base discussed below. A related obser- vation is the effect of ions on the formation of the 8 - 13 photo-adduct in E. coli tRNAfM". We found that when the photoreaction is carried out at 25"C, it requires high Na', or Mg2+. But it occurs even in low salt, if the temperature is reduced to 28°C. Cor- relatively, the loss of 4-thiouracil hypochromism oc- curs between these two temperatures, in low salt [6].
Modification oj' the Low-Salt Structure by Mn2+ or M 2 +
The Central Region of rRNAf""'. As shown in Fig. 3 - 5, the parameters of the central region respond immediately to the binding of Mn2+. In tRNAfM" (4.5 mM Na+), the absorption of 4-thiouracil is strongly reduced by the first six Mn2+ bound. From then on, the effect per Mn2+ bound is weaker but still significant up to 24 ions bound (Fig.3). Similarly, in irradiated tRNAfMe', the fluorescence of the re- duced 8 - 13 photo-adduct increases strongly for the first eight Mn2+ bound and less strongly afterwards (Fig. 4).
In native tRNAtM", one can also follow the tertiary folding due to Mg2+ by the phosphorescence of 4-thio- uracil. This is shown in Fig.5. There is little effect until one ion is bound. Then the phosphorescence increases strongly until eight Mg2 + are bound. After- wards, the increase is slower. The effect of replacing Mg2+ by Mn2+ is also shown in Fig. 5 : there is no
I I 14 - I I
red. photo -adduct
a, c
13 -
2 1 1 -
'0 10 20 [Mn]b /[tRNA]
Fig. 4. E'luore,scmcc 41' tiir retluccd 8- I3 piio~o-utkh~ct in E. coli rRNAJ~L'" 1 0 ) und of [lie ' Y base (W) in jeu.y/ /RNAPh' , versus Mn" boundper t R N A . The fluorescence of the reduced 8 ~ 13 photo- adduct is enhanced by the first M n Z + bound, whereas the 'Y' base fluorescence increases only after 4 MnZ+ are bound. Triethanol- amine chloride buffer 10 mM ; Na+ 5 mM
Total [Mg] (rnM) 1 .o o w
2ooo.01 0.1
1 1 U 1
-
I
OO 2 10 20
[Mn]b/[tRNA]
Fig. 5. Phosphorescmce qf'4-rIiiourucil in E. coli tRNA'~"" [ZO AZbo units) versus the number of M g 2 + (0) or Mn2+ ( *J bound. The structural effect on the phosphorescence cannot be seen in the case of M n Z + , due to quenching. The number of bound Mg2+ is estimated on the basis of the relative binding affinities of Mg" and Mn2' for tRNA. Cacodylate buffer 5 mM, pH 7; N a + 4.5 mM. Units for phosphorescence are arbitrary
phosphorescence enhancement. This result is due to quenching by the paramagnetic ion, and indicates binding in this region (see below).
The Anticodon Region of tRNAPh'. In contrast to the above results, the fluorescence of the 'Y' base in yeast tRNAPhe is not affected by the first ions bound. It is only after four Mn2+ are bound that it begins to increase (Fig. 4). The concentration of free Mn2+ corresponding to four Mn2+ bound is 3 pM in the given salt conditions.
J.-L. Leroy, M. Gueron, G. Thomas, and A. Favre 571
I I I I I 55 "Y+
I I 1 0 10 20
[MnlbhRNAI
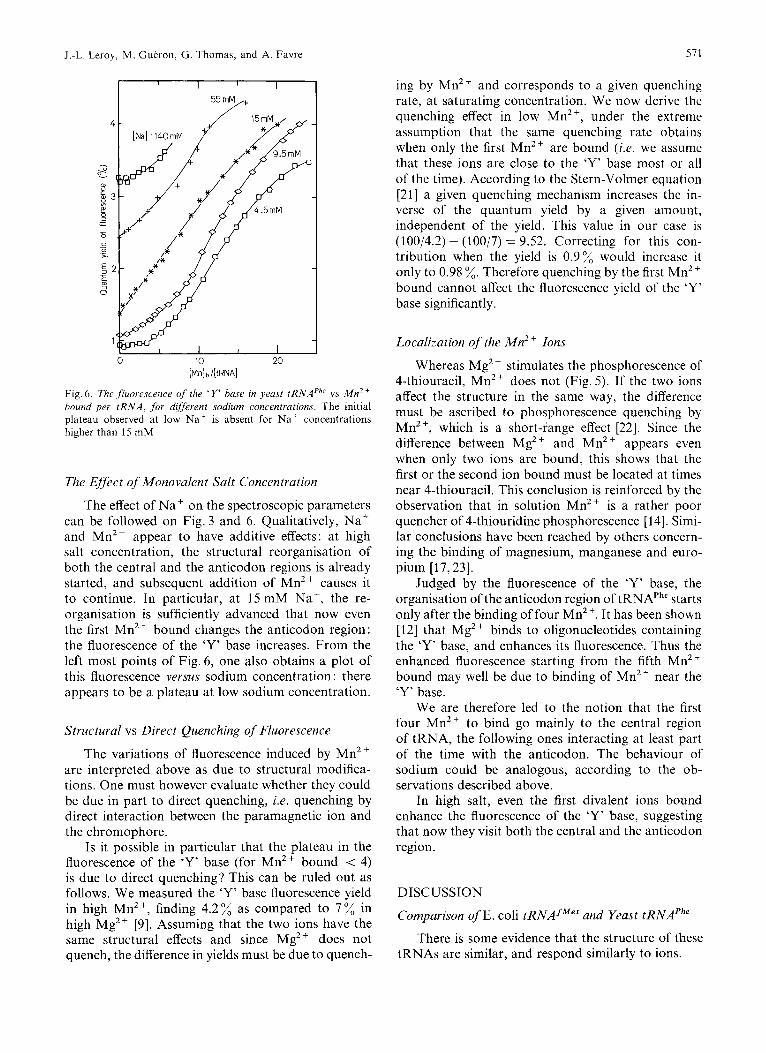
Fig. 6 . The fluorescence of the ' Y' hase in yeast tRNAPh' vs M n 2 + hound per t R N A , for different sodium concentrations. The initial plateau observed at low Naf is absent for N a + concentrations higher than 15 mM
The Effect of Monovalent Salt Concentration
The effect of Na+ on the spectroscopic parameters can be followed on Fig.3 and 6. Qualitatively, Na+ and Mn2+ appear to have additive effects: at high salt concentration, the structural reorganisation of both the central and the anticodon regions is already started, and subsequent addition of Mn2+ causes it to continue. In particular, at 15 mM Na', the re- organisation is sufficiently advanced that now even the first Mn2+ bound changes the anticodon region: the fluorescence of the 'Y' base increases. From the left most points of Fig. 6, one also obtains a plot of this fluorescence versus sodium concentration : there appears to be a plateau at low sodium concentration.
Structural vs Direct Quenching of Fluorescence
The variations of fluorescence induced by Mn2+ are interpreted above as due to structural modifica- tions. One must however evaluate whether they could be due in part to direct quenching, i.e. quenching by direct interaction between the paramagnetic ion and the chromophore.
Is it possible in particular that the plateau in the fluorescence of the 'Y' base (for Mn2+ bound < 4) is due to direct quenching? This can be ruled out as follows. We measured the 'Y' base fluorescence yield in high Mn2+, finding 4.2% as compared to 7% in high Mg2+ [9]. Assuming that the two ions have the same structural effects and since Mg2+ does not quench, the difference in yields must be due to quench-
ing by Mn2' and corresponds to a given quenching rate, at saturating concentration. We now derive the quenching effect in low Mn2+, under the extreme assumption that the same quenching rate obtains when only the first Mn2+ are bound (i.e. we assume that these ions are close to the 'Y' base most or all of the time). According to the Stern-Volmer equation [21] a given quenching mechanism increases the in- verse of the quantum yield by a given amount, independent of the yield. This value in our case is (100/4.2) - (100/7) = 9.52. Correcting for this con- tribution when the yield is 0.9% would increase it only to 0.98 %. Therefore quenching by the first Mn2' bound cannot affect the fluorescence yield of the 'Y' base significantly.
Localization of the Mn2+ Ions
Whereas Mg2 + stimulates the phosphorescence of 4-thiouracil, Mn2+ does not (Fig. 5). If the two ions affect the structure in the same way, the difference must be ascribed to phosphorescence quenching by Mn2+, which is a short-I'ange effect [22]. Since the difference between MgZ + and Mn2+ appears even when only two ions are bound, this shows that the first or the second ion bound must be located at times near 4-thiouracil. This conclusion is reinforced by the observation that in solution Mn2+ is a rather poor quencher of 4-thiouridine phosphorescence [14]. Simi- lar conclusions have been reached by others concern- ing the binding of magnesium, manganese and euro- pium [17,23].
Judged by the fluorescence of the 'Y' base, the organisation of the anticodon region of tRNAPhe starts only after the binding of four Mn2+. It has been shown [12] that Mg2+ binds to oligonucleotides containing the 'Y' base, and enhances its fluorescence. Thus the enhanced fluorescence starting from the fifth Mn2' bound may well be due to binding of Mn2+ near the 'Y' base.
We are therefore led to the notion that the first four Mn2+ to bind go mainly to the central region of tRNA, the following ones interacting at least part of the time with the anticodon. The behaviour of sodium could be analogous, according to the ob- servations described above.
In high salt, even the first divalent ions bound enhance the fluorescence of the 'Y' base, suggesting that now they visit both the central and the anticodon region.
DISCUSSION
Comparison of E. coli tRNAfMe' and Yeast tRNAPhe
tRNAs are similar, and respond similarly to ions. There is some evidence that the structure of these
572 Divalent Ions in tRNA
O . l E j [ , \ , 1 1
0 0 10 20 0 10 20
[Mn]b/ [tRNA] [Mnlb/ [ t m A l
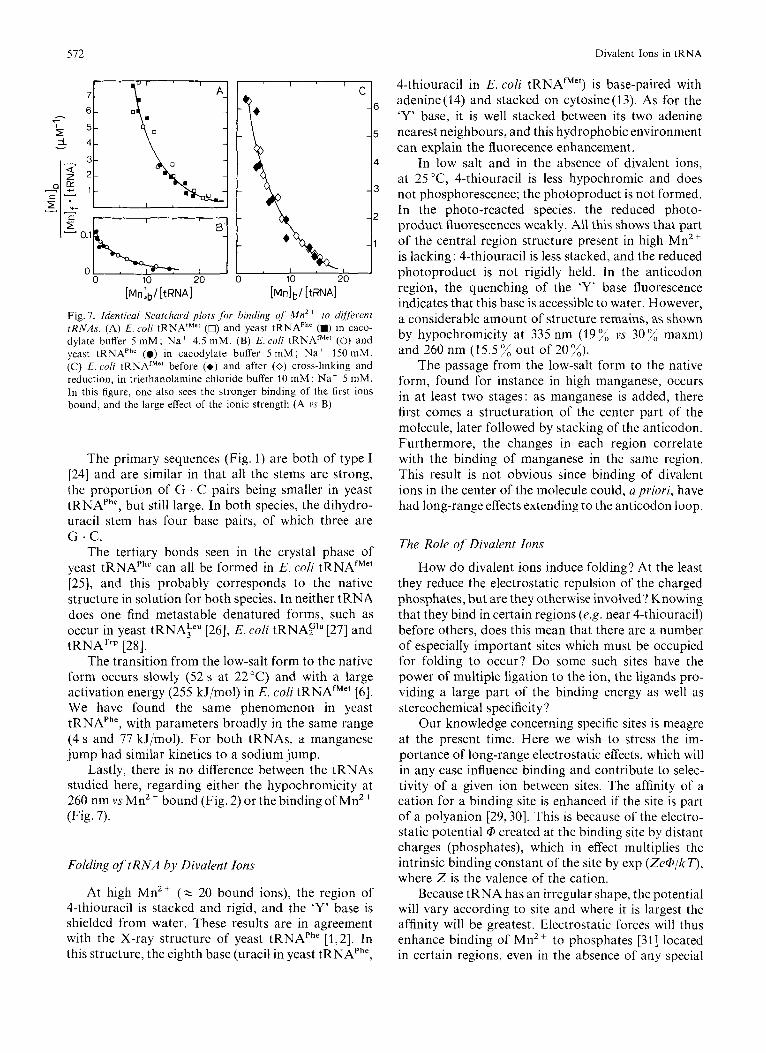
Fig. 7. Identical Scatchasd plots f;>r binding of' M n 2 + to different rRNAs. (A) E. coli tRNAfMe' (0) and yeast tRNAPh' (m) in caco- dylate buffer 5 mM; Na+ 4.5 mM. (B) E. coli tRNAfMr' (0) and yeast tRNAPhe (0) in cacodylate buffer 5 m M ; Na' 150 mM. (C) E. coli tRNAfMe' before (+) and after (0) cross-linking and reduction, in triethanolamine chloride buffer 10 m M ; Na' 5 mM. In this figure, one also sees the stronger binding of the first ions bound, and the large effect of the ionic strength (A vs B)
The primary sequences (Fig. 1) are both of type I [24] and are similar in that all the stems are strong, the proportion of G . C pairs being smaller in yeast tRNAPhe, but still large. In both species, the dihydro- uracil stem has four base pairs, of which three are
The tertiary bonds seen in the crystal phase of yeast tRNAPhe can all be formed in E. coli tRNAfMel [25], and this probably corresponds to the native structure in solution for both species. In neither tRNA does one find metastable denatured forms, such as occur in yeast tRNA?" [26], E. coli tRNAF'" [27] and tRNATrp [28].
The transition from the low-salt form to the native form occurs slowly (52 s at 22°C) and with a large activation energy (255 kJ/mol) in E. coli tRNAfM" [6]. We have found the same phenomenon in yeast tRNAPhe, with parameters broadly in the same range (4 s and 77 kJ/mol). For both tRNAs, a manganese jump had similar kinetics to a sodium jump.
Lastly, there is no difference between the tRNAs studied here, regarding either the hypochromicity at 260 nm vs Mn2 + bound (Fig. 2) or the binding of Mn2+ (Fig. 7).
G . C.
Folding q f t R N A by Divalent Ions
At high Mn2+ (z 20 bound ions), the region of 4-thiouracil is stacked and rigid, and the 'Y' base is shielded from water. These results are in agreement with the X-ray structure of yeast tRNAPhe [1,2]. In this structure, the eighth base (uracil in yeast tRNAPhe,
4-thiouracil in E. coli tRNAfMet) is base-paired with adenine (14) and stacked on cytosine (1 3). As for the 'Y' base, it is well stacked between its two adenine nearest neighbours, and this hydrophobic environment can explain the fluorecence enhancement.
In low salt and in the absence of divalent ions, at 25 "C, 4-thiouracil is less hypochromic and does not phosphorescence; the photoproduct is not formed. In the photo-reacted species, the reduced photo- product fluorescences weakly. All this shows that part of the central region structure present in high Mn2+ is lacking: 4-thiouracil is less stacked, and the reduced photoproduct is not rigidly held. In the anticodon region, the quenching of the 'Y' base fluorescence indicates that this base is accessible to water. However, a considerable amount of structure remains, as shown by hypochromicity at 335 nm (19% vs 30% maxm) and 260 nm (15.5 % out of 20%).
The passage from the low-salt form to the native form, found for instance in high manganese, occurs in at least two stages: as manganese is added, there first comes a structuration of the center part of the molecule, later followed by stacking of the anticodon. Furthermore, the changes in each region correlate with the binding of manganese in the same region. This result is not obvious since binding of divalent ions in the center of the molecule could, a priori, have had long-range effects extending to the anticodon loop.
The Role of Divalent Ions
How do divalent ions induce folding? At the least they reduce the electrostatic repulsion of the charged phosphates, but are they otherwise involved? Knowing that they bind in certain regions (e.g. near 4-thiouracil) before others, does this mean that there are a number of especially important sites which must be occupied for folding to occur'? Do some such sites have the power of multiple ligation to the ion, the ligands pro- viding a large part of the binding energy as well as stereochemical specificity?
Our knowledge concerning specific sites is meagre at the present time. Here we wish to stress the im- portance of long-range electrostatic effects, which will in any case influence binding and contribute to selec- tivity of a given ion between sites. The affinity of a cation for a binding site is enhanced if the site is part of a polyanion [29,30]. This is because of the electro- static potential @ created at the binding site by distant charges (phosphates), which in effect multiplies the intrinsic binding constant of the site by exp (Ze@/k T), where Z is the valence of the cation.
Because tRNA has an irregular shape, the potential will vary according to site and where it is largest the affinity will be greatest. Electrostatic forces will thus enhance binding of Mn2+ to phosphates [31] located in certain regions, even in the absence of any special

J .-L. Leroy, M. Gueron, G. Thomas, and A. Favre
local interactions. The corner of the 'L' is expected to have a large potential, because it is near the center of the molecule and has a large concentration of phos- phates. It would therefore be expected that Mn2+ would bind to this region first, and this should con- tribute towards explaining the affinity of Mn2+ for the vicinity of 4-thiouracil, demonstrated by the quenching of phosphorescence.
The electrostatic interpretation developed above applies to the low-salt situation where the effect of monovalent ions is not too large. In high salt, mono- valent ions will bind to the phosphates, thus reducing the electrostatic potential and, together with it, the difference in affinity between the various phosphate sites. When divalent ions are added, they will then be statistically distributed in many regions of the mole- cule. In particular, even the first Mn2+ ions bound are expected to visit both the central and anticodon regions, as observed in this work.
The electrostatic interpretation of monovalent and divalent ion binding suggests that monovalent ions may substitute for divalent ions in their struc- tural role. As they bind to phosphates, they will neutralize the repulsion and allow folding to proceed. This is indeed observed in the experiments reported here, and is also documented in the literature.
For instance the active forms of E. coli tRNATrp [32] or E.coli tRNAG'"[27] can be obtained at high sodium concentration, in the absence of divalent ions. Similarly, Reid and Robillard [33] observed by proton magnetic resonance that the tertiary structure of E. coli tRNA:"' is formed even without magnesium, in 110 mM Na'.
We thus feel that electrostatic effects are an im- portant factor in the binding of divalent ions to tRNA. Conversely, the structural role of divalent ions may be in large part the screening of the electrostatic repulsion between phosphates, thus explaining that they may be replaced by monovalent ions in their structural effects.
The possibility that a small number of sites have a special ligation is not excluded by the above con- siderations. An indication in this direction comes from binding studies at 4°C in high salt ( 2 0.1 M Na') where one would expect electrostatic effects to be weak : one binding site is found, which has a specially large affinity [34,35]. However, occupancy of this site by a divalent ion may be structurally unimportant since the present experiments show that on the one hand at least four to eight ions are needed to form the struc- ture in low salt, whereas on the other hand, the native structure is formed in high salt in the absence of divalent ions.
We thank R. G. Shulman and S. Blanquet for critical discus- sions of the manuscript. This work was supported by DClPgation Generule u la Recherche Scientqique et Technique, grant 74.7.0372 to M.G.
513
REFERENCES
1.
2.
3.
4.
5.
6.
7.
8. 9.
10.
11.
12.
13.
14. 15.
16.
17.
18.
19.
20.
21.
22.
23.
24. 25.
26
27
28
29. 30
31
32 33
34
Quigley, G. J., Wang, A. H . J., Seeman, N. C., Suddath, F. L., Rich, A,, Sussman, J. L. & Kim, S. H. (1975) Proc. Notl Acad. Sci. U.S.A. 72, 4866-4870.
Ladner, J. E., Jack, A,, Robertus, J. D., Brown, R. S., Rhodes, D., Clark, B. F . C. & Klug, A. (1975) Proc. Narl Acad. Sci.
Danchin, A. & Gueron, M. (1970) Eur. J . Biochem. 16, 532-
Cohn, M . & Townsend, J. (1954) Nature (Lond.) 173, 1090-
Yarus, M . & Rashbaum, S. (1972) Biochemistr.y, 11, 2043-
Cole, P. E., Yang, S. K. & Crothers, D. M. (1972) Biochcm-
Shalitin, N. & Feitelson, J . (1976) Biochemistrj. 15, 2092-
Favre, A. & Yaniv, M. (1971) FEBS Lett. 17, 236-240. Eisinger, J . E., Feuer, B. & Yamane, T. (1970) Proc. Notl
Beardsley, K. & Cantor, C. R. (1970) Proc. Notl Acad. Sci.
Robinson, B. & Zimmerman, T. P. (1971) J . Biol. Chem. 240.
Maelicke, A,, Von der Haar, F. & Cramer, F. (1973) 5iopolI.-
Delaney, P., Bierbaum, J . & Ofengand, J. (1974) Arcl7. Biochc~m.
Favre, A. (1974) Photochem. Photohiol. 19, 15-19. Shalitin, N. & Feitelson, J. (1973) J . Chem. P1ij.s. 59. 1045-
Pochon, F. & Cohen, S. S. (1972) Biochem. Biophj.s. Rcs.
Kayne, M. S. & Cohn, M. (1974) Biochemistrj. 13, 4159-
Favre, A., Michelson, A. M. & Yaniv, M. (1971) J . Mol. Biol.
Seno, T., Kobayashi, M. & Nishimura, S. (1969) Biodiini. Biophys. Acta, 174, 71 - 85.
Bush, C. A. (1974) in Basic Principles in Nuclei(, Acid Chern- istry, vol. 2 (T'so, P. 0. P., ed.) p. 109, Academic Press. New York.
Turro, N. J. (1967) Molecular Photocheniistrj,, pp. 92 -96. Benjamin, New York.
Eisinger, J . E. & Lamola, A. A. (1971) in Eucited States of' Proteins and Nucleic Acids (Steiner, R. F. & Weinryb, I . , eds) p. 181. Plenum, New York.
Wolfson, J . M. & Kearns, D . R. (1975) Biochemistr); 14.
Levitt, M . (1969) Nature (Lond.) 224, 759-763. Kim, S. H., Sussman, J. L., Suddath, F. L., Quigley. G. J. ,
Mc Pherson, A., Wang, A. H. J., Seeman, N. C. & Rich. A. (1974) Pmc. Nut1 Acad. Sci. U.S.A. 71, 4970-4974.
Lindahl, T., Adams, A. & Fresco, J. R. (1966) Proc. Nut1
Eisinger, J. E. & Gross, N. (1975) Bioc/~emi.sIrj', 14, 4031 -
Gartland, W. J. & Sueoka, N. (1966) Proc. Notl Acad. Sci.
Scatchard, G. (1949) Ann. N . Y . Acud. Sci. 51, 660-672. Edsall, J.T. & Wyman, J. (1958) Bioph~.sica/ Chemistry, Chap. 9.
Shulman, R. G. , Sternlicht, H. & Wyluda, B. J . (1965) J . Chem.
Ishida, T. & Sueoka, N. (1968) J . Mol. B id . 37, 313- 316. Reid, B. R. & Robillard, G. T. (1976) Nature (Lond.) 257.
Stein, A. & Crothers, D. M. (1976) Biochemistry, 15, 157- 160.
U.S.A. 72, 4414-4418.
536.
1091.
2049.
istry, 11, 4358-4368.
2097.
Acad. Sci. U.S.A. 65, 638-644.
U.S.A. 65, 39-46.
110- 117.
men, 12, 27-43.
Biophys. 61, 260- 267.
1051.
Commun. 47, 720-726.
4165.
58, 367 - 379.
1436- 1444.
Acud. Sci. U.S .A . 55, 941 -948.
4041.
U.S.A. 55, 948-956.
Academic Press, New York.
P h j . ~ . 43, 3116-3122.
287 - 291.

574 J.-L. Leroy, M. Gueron, G. Thomas, and A. Favre: Divalent Ions in tRNA
35. Bina-Stein, M. &Stein, A. (1976) Biochemisfry, 15,3912-3917. 38. RajBhandary, U. L., Chang, S. H., Stuart, A,, Faulkner, D. 36. Dube, S. K. , Marcker, K . A., Clark, B. F. C. & Corry, S. (1968) R., Hoskinson, R. M. & Khorana, H. G. (1967) Proc. Narl
Acad. Sci. U.S.A. 57, 751-758. 37. Favre, A,, Roques, B. & Fourrey, J . L. (1972) FEBS Lett. 24, 39. Nakanishi, K. , Furatachi, N., Funamizu, M., Grunberger, D.
& Weinstein, I. B. (1970) J . Am. Chem. SOC. 92, 7617-7619.
Nature (Lond.) 218,232-233.
209-214.
J.-L. Leroy and M. Gueron*, Groupe de Biophysique du Laboratoire de Physique de la Matiere Condensee, Ecole Polytechnique, F-91128 Palaiseau-Cedex, France
G. Thomas and A. Favre, Institut de Recherche en Biologie Moleculaire, Universite de Paris VII, Tour 43, 2 Place Jussieu, F-75221 Paris-Cedex-05, France
* To whom correspondence should be addressed





























