Embed Size (px)
Citation preview

Eur. J . Biochem. 70, 447-456 (1976)
The Binding Site of Protein L1 on 23-S Ribosomal RNA of Escherichia coli 1. Isolation and Characterization
Paul SLOOF, Roger GARRETT, Alain KROL, and Christiane BRANLANT
Max-Planck-lnstilul fur Molekulare Genetik, Abteilung Wittmann, Berlin and lnstitut de Biologie Moltculaire et Cellulaire du Centre National de la Recherche Scientifique. Strasbourg
(Received May 25/Septetnber 3 , 1976)
Ribonucleoproteins were prepared by ribonuclease digestion of a reconstituted complex of ribosomal protein L 1 and 23-S RNA from Escherichia coli. Three main ribonucleoproteins were identified. The largest was only obtained in an impure state at low ribonuclease concentrations, whereas the two smaller ones, which were difficult to separate from one another electrophoretically, were stable over a range of enzyme concentrations.
The two smaller ribonucleoproteins yielded a total of 13 RNA subfragments that were judged to be homogeneous electrophoretically. The latter were characterized for molecular weight and the subfragment composition of each of these ribonucleoproteins was established. Furthermore, the subfragments were shown to be maintained together in each ribonucleoprotein by RNA-RNA interactions.
The primary and specific binding site of protein L1 was localized on one continuous RNA subfragment of about 110 nucleotides in length by two newly developed binding methods.
One approach to elucidating the ribosomal struc- ture is to isolate specific ribosomal fragments that are sufficiently small for detailed structural analyses using physical-chemical and physical methods. To this end, ribonucleoproteins have been isolated con- taining a single protein and its RNA binding region by ribonuclease digestion of protein . RNA com- plexes. This approach has been successfully applied, so far, to proteins S4, S8, S15 and S20 on 16-S RNA [l], proteins L18 and L25 on 5-S RNA [2] and to proteins L20, L23 and L24 on 2 3 4 RNA [3,4]. In the present work, we describe the isolation and character- isation of a ribonucleoprotein containing protein L1.
Protein L1 is one of the largest ribosomal proteins and binds directly to 23-S RNA of Escherichia coli [ 5 ] . It has been located in the subunit interface region of the 50-S subunit by antibody binding [6]. Moreover, it is essential for the association of ribosomal subunits (together with either L11 or L16), and for the formation of the 70-S ribosomal initiation com- plex [7].
Here, we demonstrate that three specific ribo- nucleoproteins, containing L1 and differing amounts ~~
Enzyme. T1 RNase (EC 3.1.4.8).
of RNA, can be produced by TI ribonuclease digestion of a complex of L1 and 23-S RNA. The structural homogeneity of the RNA component of these ribonucleoproteins was investigated for (a) dif- ferent degrees of ribonuclease digestion and (b) dif- ferent magnesium concentrations during digestion. The optimal conditions for preparing each ribo- nucleoprotein were established and the RNA sub- fragments contained in them were identified and characterised for molecular weight. Furthermore, two new methods were developed for determining the smallest RNA region required for specific protein L1 binding. Nucleotide sequence studies on these RNA components are given in two subsequent papers
91.
MATERIALS AND METHODS
Preparation of 23-S R N A , Protein L1 and L l .23-S-RNA Complex
32P-labelled and unlabelled 23-S RNA were ex- tracted from 50-S subunits of E. coli (strains A19 and MRE 600) by three successive phenol/dodecyl- sulphate extractions [3]. Protein L1 ( M , 24500)

448 Protein L1 Binding Site on 23-S RNA
was isolated by CM-cellulose chromatography and Sephadex gel filtration [lo] and identified electro- phoretically [ l l ] . The protein batches used were provided by Dr H. G. Wittmann.
Protein-L1 . 23-S-RNA complexes were prepared by incubating for 1 h at 42 "C, in Tris/Mg/K re- constitution buffer (0.03 M Tris-HC1, pH 7.4, 0.02 M MgClz, 0.35 M KC1 and 6 mM 2-mercaptoethanol), at a five-fold molar protein excess. Complex formation was checked by a standard gel method [12].
Isolution o j the L1 Ribonucleoprotein and Characterisation ojthe R N A Subfragments
100 pg L1 . 23-S-[j2P]RNA was digested in Tris/ Mg/K buffer with 20 pg T1 RNase (Sankyo, Japan; 3300 units/mg) for 30 min at 0 "C (standard1 condi- tions). The L1 ribonucleoprotein was fraciionated at 4 "C in an 87; polyacrylamide gel containing 10 mM Tris/acetate, pH 7.8, and 10 mM magnesium acetate (standard fractionation gel), and detected autoradiographically. The gel piece containing the ribonucleoprotein was then soaked in 8 M urea for 3 h at room temperature to deproteinise and dlenature the RNA before setting onto a 12- 15 % compound polyacrylamide gel containing 8 M urea, 0.1 % do- decylsulphate, 2.5 mM EDTA and 0.1 M Tris/borate buffer, pH 8.3 [3]. This is referred to as 'a standard denaturing gel'. The approximate molecular weights of the RNA subfragments were determined by co- electrophoresing denatured S4 ribonucleoprotein, of which the molecular weights of the subfragments were known from their nucleotide sequences [13]. In order to check that the RNA subfragments were homogeneous, the bands were excised and electro- phoresed in a 15 - 20 "/, compound polyacrylamide gel as described earlier [3].
Effect of TI RNase and Magnesium Concentration on the Yield and R N A Subjragment Content of the Ribonucleoproteins
L1 . 23-S-RNA complexes were preparled and digested as described above.
TI RNase. The enzyme concentration was in- creased over the enzyme : RNA (w/w) range 1 : 10- 1 : 1. The samples were digested for identical times in Tris/Mg/K buffer, at 0 "C, before co-electrophoresing.
Magnesium. The L1 . 23-S-RNA complex was dissolved in Tris/Mg/K buffer that had been adjusted to different magnesium concentrations in the range 0.01 - 20 mM magnesium acetate. The complex was digested with T1 RNase at an enzyme : RNA (w/w) ratio of 1 : 5 in the standard way. After digestion the ribonucleoproteins were fractionated electrophoretic-
ally and their RNA subfragment patterns were examined, as described above.
Protein Binding Experiments
Nitrocellulose Filter Method. Protein L1 was bound, separately, to 23-S RNA, the total RNA component of ribonucleoprotein 2 and to RNA subfragments 1, 2, 3 and 7. The gel slices containing ribonucleo- protein 2, and the RNA subfragments 1, 2, 3 and 7 were passed three times through a small syringe and the [32P]RNA was extracted by two successive treat- ments with phenol that was saturated in 0.1 M sodium acetate, pH 5.0. The aqueous layers were dialysed overnight against Tris/Mg/K buffer. Each of the [32P]RNA components listed above was preincubated at 40 "C for 5 min and cooled slowly, to facilitate renaturation of any denatured structure ; unlabelled 23-S RNA was treated similarly. lo4- lo5 counts/min [32P]RNA were then mixed with 44 pg unlabelled 23-S RNA in 0.3 ml Tris/Mg/K buffer. Protein L1 was added at increasing molar protein : RNA ratios from 1 : 1 - 10: 1. The complexes were incubated at 40 "C for 1 h, cooled on ice and passed slowly through nitrocellulose filters (Millipore) that had been equilibrated in Tris/Mg/K buffer at 0 "C. The filter was then washed with 3 ml Tris/Mg/K buffer at 0 "C and dried under an infrared lamp. The retained 32P was estimated by liquid scintillation counting.
Electrophoretic Method. Ribonucleoprotein 2 and RNA subfragment 3 were extracted from their re- spective polyacrylamide gels and dialysed into Tris/ Mg/K buffer as described above for the filter binding method. One part of each [32P]RNA sample was electrophoresed, directly, in a standard fractiona- tion gel; another part was reconstituted with protein L1, as for the filter method, and then co-electro- phoresed with the uncomplexed RNA. The positions of the RNA components were determined autoradio- graphically.
Deproteinisation of the Ribonucleoproteins in Dodecylsulphate Gels
The gel piece containing the ribonucleoprotein was excised from the standard fractionation gel and dialysed against 0.1 % sodium dodecylsulphate, 10 mM Tris/acetate, pH 7.8, and 1 mM magnesium acetate for 4 h at 20 "C. It was then polymerised onto an 8 % polyacrylamide gel made up in the same dodecyl- sulphate-containing buffer and run overnight at room temperature. Under these conditions, the dissociation of the protein was complete since no stained band was detected when the gel was stained with Coomassie brilliant blue. The subfragment contents of the RNA components were examined in standard denaturing gels (see above).

P. Sloof, R. Garrett, A. Krol, and C. Branlant 449
A C
- L1-RNP
- TI-RNc~sQ
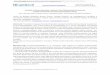
Fig. 1. Fructionution of’ rhr LI ribonucleoprotein. Complexes of L1 . 23-S-RNA were digested at T1-RNase: RNA ratio of 1 . 5 (w/w) for 30 inin at 0 “C and electrophoresed in S”/, poly- acrylamide gels containing 10 mM Trispcetate, pH 8, and 10 mM magnesium acetate for 3 h at 4 mA/tube. (A) Degraded 2 3 3 RNA stained for R N A with pyronin G. (B) Degraded L1 . 23-S-RNA complex stained with pyronin G. (C) Degraded L1 . 23-S-RNA complex stained for protein with Coomassie brilliant blue. L1-RNP = L1 ribonucleoprotein
RESULTS
Evidence for the Formation of Ribonucleoproteins
The protein-Ll . 23-S-RNA complex was digested with T1 RNase (enzyme : RNA 1 : 5, w/w) and electro- phoresed under standard conditions. One gel was stained for protein and another for RNA. The results are shown in Fig. 1. One main band was observed that stained for both protein and RNA, at amolarprotein: RNAcomponentratioofl (kO.25): 1, assuming a nucleotide length for the latter of about 140 nucleotides (see subsequent paper). It is shown below that this band actually contains two ribo- nuceloproteins that can only be separated by very long electrophoresis. At lower enzyme ratios (T1 RNase : RNA % 1 : 10 w/w), a slower-migrating RNA band was also observed (not shown) that contained protein (ribonucleoprotein 1 ). No faster-migrating ribonucleoprotein was observed, at any enzyme con- centration, which strongly suggests that no smaller ribonucleoprotein products are stable.
Conditions jbr Preparing the L1 Ribonucleoproteins
It has been shown for other ribosomal ribonucleo- proteins that their RNA content and stability is very sensitive during digestion, to both RNase and mag- nesium concentrations [3,13]. Therefore, we investi- gated their effects upon the L1 ribonucleoprotein using highly-labelled [32P]RNA complexes.
TI RNase. A series of enzyme concentrations were tested over the T1 RNase: RNA (w/w) range of 1 : 10 - 2 : 1. The results are illustrated in Fig. 2a. At the lowest enzyme ratio (1 : lo), two ribonucleo- proteins (1 and 2) were resolved in good yield. Under these conditions they were contaminated with large partially digested 23-S RNA fragments, and we were unable to purify them completely. The RF values of 1 and 2 were 0.16 and 0.20, respectively. At an enzyme ratio of 1 : 8, ribonucleoprotein 1 disappeared and only ribonucleoprotein 2 was observed that was uncontaminated with other 23-S RNA fragments. When the electrophoresis was continued for 24 h, a faster-migrating component was resolved, identical in mobility to that in sample IIIc, that did riot stain for protein. This was indistinguishable from ribo- nucleoprotein 2 in RNA subfragment composition (see below) and we concluded, therefore, that it was probably deproteinised ribonucleoprotein 2 (referred to subsequently as RNA fragment 2’); control ex- periments revealed the RNA component of ribo- nuceloprotein 2 does, indeed, migrate faster than ribonucleoprotein 2 (see below) and it was con- cluded, therefore, that some protein dissociation occurred during long electrophoresis. At higher en- zyme ratios (1 : 5) a broader band appeared in the posi- tion of ribonucleoprotein 2 (Fig. 2a). After a very long electrophoretic separation, this band resolved into two components that both stained for protein, namely ribonucleoproteins 2 and 3 (see sample IIIc in Fig. 2a). Finally, at high enzyme ratios ( 1 : 2), only the faster-migrating ribonucleoprotein 3 was present, and at even higher ratios (1 : 1) this disappeared and no smaller ones were detected.
Magnesium. The L1 . 23-S-RNA complex was digested at magnesium concentrations over the range 0.01 -20 mM at an enzyme ratio of 1 : 5. The ribo- nucleoprotein band was produced from 0.1 - 20 mM as shown in Fig. 3 a. However, at the lower magnesium concentrations it was extremely diffuse, indicating that it was structurally much more heterogeneous. The results in Fig. 3a demonstrate that in addition to the ribonucleoprotein being more homogeneous at high Mg2+ (20 mM), the yield and the resolution from other 23-S RNA fragments were also better. Above 20 mM, RNA aggregation effects were ob- served.
R N A Sub fragment Con tents ojthe LI Ribonucleoproteins
Each ribonucleoprotein that is prepared by con- trolled ribonuclease digestion of a protein . R N A complex sustains a number of cuts and possibly, excisions of sequence, at accessible RNA regions. Consequently, a pattern of RNA subfragments are released upon dissociating in a denaturing gel, that

450 Protein L1 Binding Site on 2 3 3 RNA
I A B
11 A B
111 A B C
Tv A B
Fig.2. Fractionation of: ( a ) a T1 RNase digest qf ( A ) 23-S RNA and ( B ) an LI . 23-S-RNA complex; ( b ) the RNA subfragments of the ribonucleoproteins. (a) The digestions were performed in Tris/Mg/K buffer a t a T1-RNase : RNA ratio (w/w) of (I) 1 : 10, (11) 1 :8, (111) 1 : 5 and (IV) I : 2 for 30 min at 0 "C and were electrophoresed for 16 h at 4 "C in an 8 polyacrylamide gel containing 10 mM Trisjacetate, pH 8, and 10 mM magnesium acetate with circulating buffer. 'The three ribonucleoproteins are numbered. In sample IIIc, the electrophoresis was continued, under the same conditions for a further 8 h such that ribonucleoproteins 2 and 3 were partially resolved. (b) Gel slices containing the ribonucleoproteins shown in (a) and the corresponding gel regions in the control samples were polymerised onto, and electrophoresed through, a I-cm layer of a 12 "/, polyacrylamide gel containing 8 hil urea and 0.1 7" dodecylsulphate on a 12- 15 compound gel containing 8 M urea, 0.1 M Tris/borate, 2.5 mM EDTA, p H 8.3. The samples are numbered I to IV as in (a) and A represents the control gel region and B the ribo- nucleoprotein band. B1 and B2 in sample I denotes ribonucleoproteins 1 and 2, respectively
are normally characteristic for a given ribonucleo- RNase and magnesium concentrations of preparation protein. In addition to the use of these subfragments is helpful for elucidating which parts of the RNA in sequencing and topographical studies on the region are most protected by the protein and most ribonucleoproteins (see subsequent papers), it study of stably maintained within the ribonucleoprotein. Fur- the dependence of the subfragment patterns on T1 thermore, protein binding to the isolated subfragments

P. Sloof, R . Garrett, A. Krol, and C. Brdnlant 451
1 2 3
Fig.3. Fructionution of: [ m ) II TI R N u x digest q f ( A ) 23-S R N A and ( B ) un Ll . 23-S-RNA comp1e.r; [ h ) the R N A suhfrugmrnts of rhe riho- nuckoproteins. (a) The digestions were performed at a TI-RNase : RNA ratio of 1 : 8 (wlw) in 0.03 M Trisjacetate, pH 7.8. and magnesium acetate concentrations of (1) 20 mM, (2) 1 mM, (3) 0.1 mM, (4) 0.01 m M and for 30 min at 0 ”C. The digests were electrophoresed as in Fig. 2a. The ribonucleoproteins are indicated by arrows. The Iow-Mg” samplcs were divided into sections for subfragment analyses. (b) Gel slices containing the ribonucleoproteins excised from the gels shown in (a) were electrophoresed in a standard denaturing gel. The subfragment patterns are shown for the ribonucleoproteins in sample 1 (20 m M Mg”) and sample 4 (0.01 mM Mg”). a, b and c refer to the different sections of the ribonucleoprotein in sample 4 that were co-electrophoresed
yields information on the RNA section directly in- volved in protein binding.
TI RNuse. The RNA subfragment patterns of the ribonucleoproteins produced with increasing en- zyme concentrations (Fig. 2a) and the corresponding
RNA fragments obtained from digested 23-S RNA are shown in Fig.2b. The following changes in the subfragment patterns occurred at increasing enzyme concentrations. (a) There was a decrease in size of the largest subfragment from about 135 nucleotides

452
S4- RN P
Protein L1 Binding Site on 23-S RNA
L1-RNP
Fig.4. Molecular weight determination of the RNA subfragments from the ribonucleoproteins. On the left are the subfragments from an S4 ribonucleoprotein (SCRNP) that are of known sequence [l3]. On the right are the subfragments obtained from the ribonucleoproteins (see Fig.2a) that were co-electrophoresed with the S4 rib'onucleoprotein in a denaturing gel. The migration distance of the S4 RNA sub- fragments are plotted against the log of nucleotide length, n. The nucleotide lengths of the RNA subfragments from L1 ribonucleoprotein (LI-RNP) were calculated from this line and are given on the right
in ribonucleoprotein 1 to about 110 nucleotides (see below) in ribonucleoprotein 3 (samples I and IV in Fig. 2 b). (b) The complexity decreased between ribo- nucleoproteins 2 (sample 11) and 3 (sample I'V). For the latter only three subfragments, namely 3 , 7 and 12 were generally observed, although occassionally small amounts of 11 were also detected. Less complex ribo- nucleoproteins were never found. In order to establish whether any of the bands contained subfragment mixtures they were excised from the gel. Subfragments 1 - 9 were electrophoresed in 15 - 20 % conipound polyacrylamide gels containing 8 M urea (see Ma- terials and Methods) and subfragments 10- 3 3 were electrophoresed in a 20% gel with 8 M urea. After overnight electrophoresis single sharp bands were detected and there was no evidence of further resolu- tion of any of the bands, indicating that they were all very likely to be homogeneous. 23-S RNA prepared from both E. coli strain A19 and strain MIRE 600 was used in these experiments. Although subfragments with the same electrophoretic mobilities were resolved for ribonucleoproteins, differences in the relative in- tensities were observed. Subfragment bands 1 and 4 from strain A19 were much stronger than for strain MRE 600 whereas subfragments 8 and 9 were weaker. For ribonucleoprotein 3, no differences were observed between the two strains. In all of the figures results from strain A19 are presented.
Magnesium. The subfragment patterns of the ribo- nucleoproteins prepared over a range of MgZt con- centrations (Fig, 3 a) were examined. The results are shown in Fig. 3 b for the two extreme concentrations of 20 mM and 0.1 mM Mg". At 20 mM Mg2 ' the subfragment pattern corresponded to ribonucleo- protein 2 which contains subfragment 3 in low yield (Fig. 3 b). At 0.1 mM Mg2+ the ribonucleoprotein formed a broad band (Fig. 3 a) and was, therefore, split into three sections. The upper section (a) contained almost pure subfragment 1 which indicates that all the smaller subfragments have dissociated. Section (b) contained a mixture of subfragments 1 and 2 and section (c) contained mainly subfragment 2. These large subfragments were absent in the control gel pieces (data not shown). Although faster-migrating bands were present in (b) and (c), most of them do not correspond in mobility to L1 ribonucleoprotein sub- fragments and they derive from degraded 23-S RNA fragments that are visible in the control in Fig. 3a (gels 3A and 4A, respectively). At increasing Mg2+ concentrations (from 1 - 20 mM), an increasing num- ber of smaller RNA subfragment bands from the L1 ribonucleoprotein were identified in the denaturing gels (data not shown). This evidence suggests that under the low Mg2 conditions, RNA subfragments tend to dissociate from the ribonucleoproteins producing more heterogeneous ribonucleoprotein moieties, but

P. Sloof, R. Garrett, A. Krol, and C. Branlant 453
ultimately leaving mainly the largest subfragments 1 and 2 in the ribonucleoproteins. One interpretation of this is that RNA subfragments 1 and 2 are parent subfragments for the others. This point is considered again in the Discussion.
Molecular Weights of the RNA Subfragments. The molecular weights of the RNA subfragments of the L1 ribonucleoproteins were determined by comparing their electrophoretic mobilities with the subfragments of the S4 ribonucleoprotein that are of known nucleo- tide sequence. The S4 ribonucleoprotein was provided by E. Ungewickell. The results are presented in Fig. 4 and show that subfragments 1 - 13 are between 17 and 130 nucleotides long. The size of the largest sub- fragment 1 ' from ribonucleoprotein 1 was determined in a separate experiment, and found to be 135 nucleotides long.
Localising the Primary Protein L1 Binding Site within the Protected RNA Region
The purpose of the binding experiments was two- fold. First to establish that the RNA regions of the ribonucleoproteins contained the primary specific binding site of protein L1, by showing that similar binding occurred to the RNA site as to the whole 23-S RNA, thereby establishing that the protein had not migrated to another site during the complex digestion. Second, to determine the smallest RNA subfragment to which the protein could still bind strongly and specifically, thereby localising the minimum length of RNA containing the primary protein binding site. Two methods were developed for these purposes, namely a nitrocellulose filter method and an electro- phoretic method.
Nitrocellulose Filter Method. The binding curves for protein L1 with 23-S RNA and the RNA component of ribonucleoprotein 2 are shown in Fig. 5 a, b. They show saturation of complex retention between 40 and 45 at a molar excess of input protein. We concluded that the binding was specific (see below) and the near identity of the binding curves suggested, moreover, that the binding constants of the protein to the two RNA's were similar. The binding curves for the sub- fragments 1, 2, 3 and 7 are shown in Fig. 5c, d, e, f. The first three show saturation of retention, and closely resemble those with 23-S RNA and the RNA com- ponent of ribonucleoprotein 2, whereas subfragment 7 does not. There are two main conclusions. First, sub- fragments 1, 2 and 3 all contain the main specific binding site of protein L1; the result for subfragment 3 shows that the protein binding site is contained within about 110 nucleotides (Fig. 4). The binding to sub- fragment 7 (71 nucleotides) is weak and shows no saturation. Although for this subfragment we cannot distinguish between specific and unspecific binding, the most likely explanation is that the binding is specific
60 la
60 lc 20
0 0 2 4 6 8 10 0 2 4 6 8 10
0 0 2 4 6 8 10 0 2 4 6 8 10
Protein/RNA (mollmol) - Fig. 5. Binding saturation curves. These curves were prepared for the binding of protein L1 with (a) 23-S RNA, (b) the RNA com- ponent of ribonucleoprotein 2 and ( c - f ) the R N A subfragments 1, 2, 3 and 7, respectively. Protein-RNA binding was estimated from the degree of retention of [3ZP]-RNA on nitrocellulose filters as described in Materials an Methods. The percentage retention expresses the 32P retained on the filter as a percentage of the input 32P (in counts/min)
and weak due to a partial loss of part of the primary binding site. Second, two trends are discernible in the curves in Fig. 5 that may be significant. (a) There is a shift in the onset of saturation of retention to higher molar protein inputs for smaller RNA regions sug- gesting that a slight decrease in binding constant may occur in the transition from 2 3 4 RNA to sub- fragment 3. (b) The percentage retention at the plateau increases gradually from 23-S RNA (40 z) through to subfragment 3 (52 %) which suggests that when less RNA is present, the protein is more accessible for binding to the filter surface. This method was de- veloped with a view to distinguish specific from un- specific binding. To this end, experiments were per- formed in which ribosomal proteins (e,g. S8) were incubated with ribosomal RNA's in Tris/Mg/K buffer where they bind specifically, and in low salt (Tris/ Mg buffer) where they bind unspecifically (e.g. [14]). For the former, a plateau was always reached at inter- mediate levels of retention (30 - 60 %), whereas for the latter, the retention always increased linearly to almost 100 %, presumably because the molar protein : RNA ratio of 1 : l is surpassed, leading to more

454 Protein L1 Binding Site on 23-S RNA
A + L l -L l
0-
B + L1 - L.1
Fig. 6. Electrophoretic defrmminofion ofprotein binding. (A) The RNA component of ribonucleoprotein 2 was electrophoresed in a standard. fractionation gel, with and without specifically bound protein L1. (B) RNA subfragment 3 electrophoresed, as in (A), with and without bound protein L1
effective filter binding. Nevertheless, two control ex- periments were always necessary. First, to show that the RNA component alone is not retained on the filter, under the conditions of the experiment and, second, to show that the filter is not saturated with protein, by establishing that the same percentage retention is obtained when double quantities of pro- tein and RNA are passed through the filter. The latter controls were performed in each of the above experi- ments.
Electrophoretic Method. This binding me1 hod is based on the principle that a small RNA region will migrate differently in the presence and absence of a bound protein. Protein L1 is especially suitable for this approach because it is both basic and large. The RNA component of ribonucleoprotein 2, and subfragment 3 were extracted from their respective gels. Each RNA moiety was dissolved in Tris/Mg/K reconstitution buffer and divided into two parts. One part was in- cubated with protein L1 and the other was no(. Both parts were then co-electrophoresed in a standard fractionation gel. The results, which are given in Fig.6, show that there was a large migration dif- ference between the complex and its RNA compo- nent. We believe this difference is attributable to (a) the higher molecular weight of the complex and (b) the lower net negative charge in the complex. Reasons for believing that no major conformational
A
-0
-1 -2 -3 - A -6 -7
- 11 - 12
- 13
Fig. I. The R N A components oJ rihonucleoprotrin (2+.?). (A) The gel piece containing the ribonucleoprotein was dialysed against 0.1 dodecylsulphate, 1 mM magnesium acetate and 10 mM Tris/acetdte, pH 7.8, for 4 h at 20 "C to dissociate the protein and then electrophoresed at room temperature in an 8 % polyacryl- amide gel containing the same buffer. (B) The subfragments of the RNA components 2 and 3 were separated in a standard denaturing gel
change occurs in the RNA structure on protein bind- ing are considered in the subsequent papers.
RNA-RNA Interactions within the Ribonucleoproteins
The RNA subfragments within a ribonucleoprotein are maintained together either by RNA-RNA, or by protein-RNA interactions, or both. The importance of RNA-RNA interactions in the L1 ribonucleo- protein was investigated by deproteinising the mix- ture of ribonucleoprotein (2 + 3) prepared at an enzyme ratio of 1 :5 (Fig.2a) by dodecylsulphate treatment, in the presence of 1 mM magnesium, and electrophoresing the RNA components. The results in Fig. 7A demonstrate two well-resolved RNA bands. They were not stained with Coomassie brilliant blue indicating their complete deproteinisation. They were excised from the gel and co-electrophoresed in a standard denaturing gel. The results in Fig. 7B show

P. Sloof, R. Garrett, A. Krol, and C. Branlant 455
that they have different subfragment compositions. The subfragments were readily identified by com- parison with the results shown in Fig.2b. Between them, the two RNA components account for all of the main subfragments; RNA band 2 contains sub- fragments 1, 2, 4, 6, 11, and 13 and RNA band 3 contains 3, 7, and 12. In some experiments subfrag- ments 3 and 7 also occurred in RNA band 2, indicating that additional cuts were produced in one of the larger subfragments. These cuts are considered in detail in the subsequent paper. These two RNA components have the same subfragment patterns as ribonucleo- protein 2 and 3 (see Fig. 2).
Since these two sets of subfragments co-migrate in their respective RNA complexes (Fig. 7A), in the absence of protein, they must be held together exclu- sively by RNA-RNA interactions, and, therefore, the protein is not essential for maintaining them together. We cannot completely eliminate, however, that some very small subfragments that are not recovered in the denaturing gel dissociate with the protein.
DISCUSSION
Digestion of reconstituted protein-Ll . 23-S-RNA complexes with T1 RNase yielded L1 ribonucleopro- tein that could be fractionated on 8 % polyacrylamide gels, in the presence of 20 mM Mg2+ that was essen- tial for the maximum stability of the ribonucleo- protein. Three ribonucleoproteins were identified at different enzyme concentrations. The number of RNA subfragments within each of them and their nucleotide lengths were determined electrophoretic- ally.
The specificity of the ribonucleoprotein was check- ed by reconstitution experiments in which a specific and similar binding was demonstrated between protein L1 and 23-S RNA, the whole RNA region of ribo- nucleoprotein 2 and subfragments 1, 2 and 3. This suggests that subfragments 1, 2 and 3 are parent sub- fragments and that subfragment 3 (1 10- 120 nucleo- tides) is the primary and specific binding site of pro- tein L1. Two new binding methods were developed for these experiments that should prove especially useful for studying interactions between proteins and relatively small RNA fragments.
From the electrophoretic migration of the ribo- nucleoprotein, and from the subfragment patterns, one cannot make accurate estimates of the molecular weight of the whole RNA region. Indeed, this is only possible from the nucleotide sequence data that is presented in the subsequent papers. However, there are some suggestions from the protein binding ex- periments, and from the RNA subfragment com- position of ribonucleoproteins prepared at low mag- nesium concentrations, that subfragments 1, 2 and 3
may be the parent subfragments for most of the smaller ones. This inference is reinforced by experi- ments in which the ribonucleoproteins were treated consecutively with 2 M and 4 M urea (data not given), as was performed earlier for the protein S4 ribonucleoprotein [lS]. This resulted in the stepwise removal of some of the smaller RNA subfragments without altering the electrophoretic mobility of either the L1 ribonucleoprotein or of its RNA component. In other words, as demonstrated earlier for other ribo- nucleoproteins [3,1S], although each ribonucleopro- tein is approximately sequence homogeneous, it is heterogeneous with respect to the number of enzyme cuts in a few positions.
Some inferences can be made about the organisa- tion of the RNA and protein L1 in the ribonucleo- protein. First, the data presented show that all of the subfragments can be fairly strongly maintained in the ribonucleoproteins by RNA-RNA interactions alone. Second, protein L1 is very accessible in the SO-S sub- unit [16], unlike some other proteins that produce ribonucleoproteins (e.g. S20 and L24). This acces- sibility is compatible with the relatively small RNA binding site of protein L1 of about 110 nucleotides.
In conclusion, nucleotide analyses of the subfrag- ments, and of the whole RNA region, are necessary in order to confirm or extend the above results and conclusions. Moreover, such analyses will facilitate the localisation of the RNA region within the 23-S RNA molecule and give the sequence relationship between the subfragments. These objectives are at- tained in the next paper.
We thank C. Schulte for help in characterising protein L1. P. S. received a long-term EMBO fellowship. A.K. receivcd a fellowship from lu Ligue Franpzis Confre le Cancer. The work was supported by grants from EMBO and the DeutscIit, Forschungs- gerneinschajt (Lo R. G.) and by the Cmtre National de la Recherchr Scientifique. Drs J. P. Ebel and H. G. Wittmann are thanked for their encouragement and support.
REFERENCES
1 . Zimmermann, R . A. (1974) in Rihosomes (M. Nomura, A. TissiPrea & P. Lengyel, eds) pp. 225-269, Cold Spring Harbor Press, New York.
2. Gray, P. N., Bellemare, G . , Monier, R., Garrett, R. A. & Stoffler, G . (1973) J . Mol. Biol. 77, 133- 152.
3. Branlant, C., Krol, A,, Sriwidada, J . , Fellner, P. & Crichton, R.
4. Branlant, C., Krol, A , , Sriwidada, J., Ebel, .I. P., Sloof, P.
5. Garrett, R. A,, Muller, S., Spierer, P. & Zimmermann, R. A.
6. Morrison, C. A,, Garrett, R. A,, Zeichhardt, H. & Stoffler, G .
7. Kazemie, M. (1975) Eur. 1. Biochem. 58, 501 -510. 8. Branlant, C., Krol, A , , Sriwidada, J. , Ebel, J. P., Sloof, P.
& Garrett, R. A. (1976) Eur. J . Biochem. 70,457-469. 9. Branlant, C., Korobko, V. & Ebel, J . P. (1976) Eur. J . Bioclzcrn.
70, 471 -482.
(1973) FEBS Lett. 35, 265 - 272.
& Garrett, R. A. (1975) FEBS Lef t . 52, 195-201.
(1974) J . Mol. Biol. 88, 553- 557.
(1973) Mol. G m . Genet. 127, 359-368.

456 P. Sloof, R. Garrett, A. Krol, and C. Branlant: Protein L1 Binding Site on 2 3 3 RNA
10. Hindennach, I., Kaltschmidt, E. & Wittmann, H. G. (1971)
11. Kaltschmidt, E. & Wittmann, H. G. (1970) Anal. Biochem. 36,
12. Schulte, C., Morrison, C. A. & Garrett, R. A. (1973) Bio-
13. Ungewickell, E., Garrett, R. A,, Ehresmann, C., Stiegler, P.
14. Schulte, C. & Garrett, R. A. (1972) Mol. Gen. Genet. 119,
15. Ungewickell, E., Ehresmann, C., Stiegler, P. & Garrett, R. A.
16. Morrison, C. A. (1973) Doctorate Thesis. Free University of
Eur. J . Biochem. 23, 12- 16. 345- 355.
(1975) Nucleic AcidRes. 2, 1867- 1888. 401 -412.
chemistry, 13, 1032- 1037. Berlin.
& Fellner, P. (1975) Eur. J . Biochem. 51, 165- 180.
P. Sloof and R. A. Garrett, Max-Planck-Institut fur M'olekulare Genetik, IhnestraOe 63/73, D-1000 Berlin (West) 33-Dahlem
A. Krol and C. Branlant, Institut de Biologie Moleculaire et Cellulaire du C.N.R.S., 15 Rue Rene-Descartes, Esplanade, F-67000 Strasbourg, France