Embed Size (px)
Citation preview

Eur. J . Biochem. 57, 231-239 (1975)
The Regulation of Arginine Biosynthesis in Saccharomyces cerevisiae The Specificity of argR- Mutations and the General Control of Amino-Acid Biosynthesis
Jacqueline DELFORGE, Francine MESSENGUY, and Jean-Marie WIAME
Institut de Recherches du Centre d’Enseignement et de Recherches des Industries Alimentaires et Chimiques, and Laboratoire de Microbiologie de I’Universite Libre de Bruxelles
(Received April 5/May 24, 1975)
The regulation of arginine biosynthetic enzymes in yeast is subjected to a double control. One level of arginine enzyme synthesis is under the control of an apo-repressor, called ARGR. ARGR molecules control specifically the arginine pathway.
A second level of control of arginine biosynthesis has been disclosed. It also controls tryptophan, histidine, lysine, isoleucine-valine and leucine, and probably many more biosyntheses. The general mechanism is turned on in leaky mutants in any of the amino acid pathways mentioned above.
Very little is known yet about the regulation of amino acid biosynthesis in eucaryotic cells. The existence of only a few regulatory mutants (argR [l], cpaR [2], eth2 [3]) of the “apo-repressor” type has been reported. More recently Bollon [4] gave good evidence that in yeast also threonine deaminase, the first enzyme of isoleucine biosynthesis, is somehow involved in the control of its own synthesis.
Several laboratories have shown that in fungi Neurospora and yeast [5-81, a metabolic block in one particular biosynthesis such as arginine, lysine, histidine or tryptophan leads to a modification of the enzyme levels in several other pathways. Mutants affecting this effect have been obtained. Two classes of general biosynthetic mutants exist in yeast. The first class of mutants are unable to derepress arginine, tryptophan, histidine biosynthetic enzymes under derepressing conditions and are therefore sensitive to 3-amino-l,2,4-triazole [7,8]. In contrast, in the second class the levels of enzymes in these pathways are de-
Enzymes. L-Ornithine carbamoyltransferase (EC 2.1.3.3); fumarase or L-malate hydrolyase (EC 4.2.1.2); histidinol dehydro- genase or NAD oxidoreductase (EC 1.1.1.23); anthranilate synthase or chorismate pyruvate-lyase (amino acceptor) (EC 4.1.3.27); saccharopine dehydrogenase (NAD, lysine-forming) (EC 1.5.1.7); phosphoribosyl-ATP pyrophosphorylase (EC 2.4.2.12); NADP glutamic dehydrogenase (EC 1.4.1.4); NAD glutamic dehydro- genase (EC 1.4.1.2); argininosuccinase (EC 4.3.2.1); B amino- transferase (valine, isoleucine, leucine) (EC 2.6.1.42) ; imidazol- glycerolphosphate dehydratase (EC 4.2.1.19).
repressed even under non-derepressing conditions [8]. They have been selected as thermosensitive mutants but where they are really blocked is as yet unknown. However this situation looks quite different from that described in Salmonella typhirnurium with some of the histidine regulatory mutants : hisT, his W and hisU [9] mutants are presumably involved in tRNA maturation or tRNA modification enzymes, so that in these mutants not only histidine enzymes but several other biosynthetic enzymes are derepressed [lo], because unmodified tRNA’s cannot act properly as corepres- sors.
Several years ago, Bechet et al. [l] reported the existence of three complementation classes of regula- tory mutants in the arginine biosynthesis : argRI- , argRII- and argRIII- . These mutants have derepres- sed and constitutive levels for several enzymes in- volved in arginine biosynthesis. The constitutive levels of enzymes in argRI-, argRII- and argRIII- are the same in all three types of mutants, even when the three are combined in a single haploid strain, yet this constitutive and derepressed level is not the highest level of derepression obtainable for arginine biosyn- thetic enzymes : an arginine-leaky mutant can de- represse the level of L-ornithine carbamoyltransferase 5 - 6-fold compared to wild type, and 2- 3-fold compared with argR- mutants [ll].
Hoet and Wiame [12] gave some evidence as to the likelihood that argRI- , argRII- and argRIII- are
Eur. J. Biochem. .57 (1975)

232 The Specificity of argR- Mutations and the General Control of Amino-Acid Riosynthesis
mutations in an "apo-repressor" molecule rather than in the co-repressor (if this were arginyl-tRNA for instance). However, no investigations have been made so far about whether or not these mutations are acting specifically on arginine biosynthetic enzymes.
In this paper we first show that all three argR- mutations are indeed specific for the arginine pathway and we also show by the use of leaky mutations and the analysis of general amino acid pools that there is a second independent level of control, acting not only on arginine biosynthetic enzymes but also on histidine, lysine, tryptophan, isoleucine-valine and probably other biosynthetic pathways.
MATERIALS AND METHODS
Strains
Most of the strains used in this work are isogenic with our wild-type strain C1278b, mating type a.
~~ ~-
Strain Mating Genotype number type
wild type C1278b a 3962c a ] 85102 a BJ210 s(
BJ240 a BJ313 a 8020c a 5716d a
4426-td a 5 7 1 6 ~ -
DJ3 a
7305b c(
FM6916 a
7366b a
7368a a
MG409 a 7305d a MG583 a
BH14 a
7 3 3 0 ~ -
MG672 a
arg R l - -2 urgRll- -10 urgR//--lO nrgRII l - - l (isogenic with 1705d another urgRIII--l I wild-type strain) argRI- -1 2, arg RIII - - I argRI--12, argRll--40, argRIII- -1 aas2: 3-amino- 1,2,4-triazoIe-sensitive strain
isogenic with S288c. This mutant wdS kindly sent to us by G. R. Fink
original mutant-canR-argRII- -51, leaky-lysine auxotroph
segregant from a cross of DJ3 by wild-type (argRII--51, leaky-lysine auxotroph)
original mutant canR. Slightly leaky (isoleucine-valine) f I auxotroph
Segregant from the cross of mutant FM6916 by wild type (ile-van k I
Segregant from the cross of 7366b by 3501c (avgRI--2, mating type a). contains 2 mutations: argRI--2, (ile-Val) f I
argJ k : arginine-leaky strain, block unknown lys' : leaky-lysine auxotroph. Block unknown (ile-val) f 2 : leaky-isoieucine-vdline auxotroph
his' : leaky-histidine auxotroph revertant from revertant derived from yeast foam (a)
MG 389 ( h i s - ) : an external suppressor is responsible for the leakiness (M. Crabeel, Ph.D. Thesis)
urgRII- -51, arg' crvg(B,C/ nonsense mutations
(M. Minet, Ph.D. Thesis)
Mutants carrying the argR- mutations have been described elsewhere [l].
Cultures and Media
Cells were grown at 29 "C under conditions described earlier [30] in medium no. 150. This medium was buffered at pH 6.1 with citric acid-KOH, and supplemented with glucose and vitamins (final concen- tration 3%). This medium will be designated as M and nitrogen nutrients or other additions will be designated explicitly. If the additive is ammonia it is made 0.02 M in (NH,),SO,; if the additive is an amino acid it is usually at 1000 pg/ml, if not mentioned otherwise. Cells were harvested at a maximal ab- sorbance of 0.600 measured with a Beckman B spectro- photometer, which represents about 5 x lo6 cells per ml. The cells were washed and resuspended after centrifugation with 0.02 M Tris-HC1 buffer pH 8, or with bidistilled water when several different enzyme assays were performed with the same cell extract. Complete medium no. 868 has been described else- where [ 131.
Isolation of Mutants
Mutants were obtained after treatment of wild-type cells (for mutant DJ3) or strain MG672 (for mutant FM6916) with ethyl methanesulfonate for 1 h at 29 "C in 0.02 M potassium phosphate buffer pH 8. After that treatment 6% of sodium thiosulfate was added and the cells washed three times with bidistilled water. The cells were allowed to grow for 6 h in liquid complete medium no. 868 at 29°C. The cells were then washed and plated on NHZ + L-ornithine (200 pg/ml) and 3 pg/ml, 5 pg/ml or 8 pg/ml of L-canavanine, and 2- 5 x lo7 cells were put on each plate. Canavanine- resistant colonies were allowed to grow for several days at 29 "C. Under these conditions about 100 colo- nies grew per plate. All sizes of colonies were picked up. Most of them were permease-less mutants. These were discarded after the following tests: only general amino acid permease mutants (gap-) [14] grow on proline + 1 mg per ml of D-histidine, specific arginine permease mutants (argp-) are resistant to high cana- vanine concentrations (NH; + 50 to 100 pg of cana- vanine per ml). Regulatory mutants are not resistant to canavanine in the absence of ornithine. All other canavanine-resistant mutants were purified twice on complete medium and rechecked for canavanine resistance and no growth on medium M + ornithine as nitrogen source [15,1], so that the phenotype of an arginine regulatory mutant has to be: growth on M + NH,' + 8 pg/ml of L-canavanine + 200 pg/ml
Eur. J. Biochem. 57 (1975)

J . Delforge, F. Messenguy, and J.-M. Wiame 233
Table 1, Specific activities of ornithine carbamoyltransjerase und argininosuccinase in wild type, argR- mutant and in leaky arg' mutant Ornithine carbamoyltransferase was measured as citrulline produced and argininosuccinase as urea produced
Genotype and name of strain Nitrogen nutrients Enzyme activities
ornithine carbamoyltransferase argininosuccinase
pmol h-' mg protein-'
Wild type N H f 32 2.1 (C1278b) NHf + arginine 3 1.8 argRI- -2 NH: 56 (B J 1 02) NHZ + arginine 51 argRII--10 N H f 61 1.9 (BJ210) NHZ + arginine 48 argRIII- -1 NHf 64 (BJ313) NH: + arginine 58
(5716d) NHZ + arginine 59 1.7
-
-
-
-
-
argRI--12, argRIII- -1 NH: 66 2.0
argRI- -12, argRII--lO, argRIII- -1 N H f 59 (57 16c) NHf + arginine 55
-
-
arg' NH; 208 11.6 (MG409) NH: + arginine 4 2.3 arg', argRII--51 NH: 193 -
- (7 3 30c) NHf + arginine 54
of L-ornithine and no growth on medium M with ornithine as nitrogen source.
Chemicals
Arginine, ornithine, carbamoylphosphate, Tris buffer, argininosuccinic acid barium salt, phospho- ribosylpyrophosphate, L-histidinol, 2-oxoisovaleric acid, L-malic acid, chorismic acid were Sigma products.
Anthranilic acid was purchased from Calbiochem. L-Canavanine sulfate was a Schwarz Mann product. 3-Amino-l,2,4-triazole was an Aldrich Co-Inc. pro- duct. Purified arginase from rat liver (for arginino- succinase assay) was a gift from M. Penninckx [16].
Enzyme Assays
Enzymes were extracted by disruption of the cells with the French pressure cell (liquid was 0.02 M Tris- HCl buffer pH 8 or water). The pellet was discarded by centrifugation at 15000 rev./min at 0 "C in a Sorvall RC 2B centrifuge for 15 min. This crude extract was used right away for ornithine carbamoyltransferase and fumarase activity measurement, and was dialysed against appropriate buffers for: argininosuccinase, B aminotransferase, saccharopine dehydrogenase, an- thranilate synthase assays. Specific methods for en- zyme assays are referred to below: fumarase [17], L-ornithine carbamoyltransferase [12], histidinol de- hydrogenase [18], anthranilate synthase [19], sac- charopine dehydrogenase [20], phosphoribosyl-ATP
pyrophosphorylase [21], NADP glutamic dehydro- genase [22], NAD glutamic dehydrogenase [23], argininosuccinase [24], B aminotransferase [25]. Pro- tein was determined with the method described by Lowry et al. [26].
Measure of amino acid pools: 250 ml of exponen- tially growing cells, on M + NH,f medium (about 2 to 5 x lo6 cells per ml) were filtered on a millipore and washed four times with 15 ml of bidistilled water at 0 "C. The cells were then resuspended into 10 ml of 5 % cold trichloracetic acid. Extraction of the amino acids was continued for 20 min, in the cold room, with gentle stirring. The cells were filtered on a millipore. The liquid was collected into a vessel, and the filter rinsed three times with 4 ml of cold 5 % tri- chloracetic acid. The liquid volume was adjusted to 25 ml. An aliquot of this preparation (1 ml) was then passed through a Beckman amino acid analyzer (Model 120 c) following the method described by Moore and Stein [27].
RESULTS
Research of a Regulatory Mutant in Arginine Biosynthesis with Highly Derepressed Ornithine Carbamoyltransferase Levels
Table 1 shows the different levels of ornithine carbamoyltransferase and argininosuccinase activities in wild type, argR- mutants and in an arginine auxo-
Eur. J. Biochem. 57 (1975)
234 The Specificity of argR- Mutations and the General Control of Amino-Acid Biosynthesis
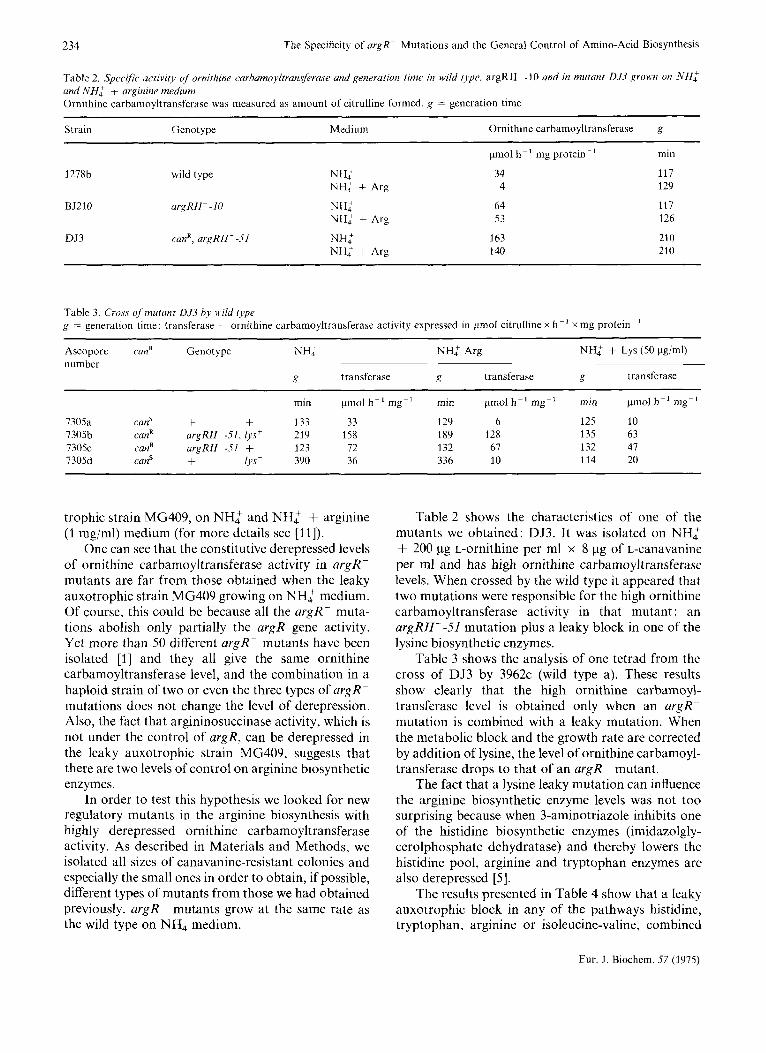
Table 2 Specific artivify of ornitlune carbamoyltransferase and generutlon t m e in wild ripe, argRII --I0 and in mutant DJ3 groun on NH: und NH: + arginine mediuni Ornithine carbamoyltransferase was measured as amount of citrulline formed g = generation time
Strain Genotype Medium Ornithine carbamoyltransferase g
1278b wild type NH:
BJ210 nrgRII- -10 NH:
DJ3 can', argRII- -51 NH:
NH: + Arg
NH: + Arg
NH: + Arg
pmol h-' mg protein-' min
34 117 4 129
64 117 53 126
3 63 210 140 210
Table 3. Cross ofmutant 053 by wild type g = generation time; transferase = ornithine carbamoyltransferase activity expressed in pmol citrulline x h-' x mg protein-'
Ascopore can' Genotype NH: NH: Arg NH: + Lys (50 pg/ml) number
a transferase g transferase g transferase
min p o l h - ' mg-' min pmol h- ' mg-' min pmol h- ' mg-'
7305a canS + + 133 33 129 6 125 10 7305b can' argRII--51, lys' 219 158 189 128 135 63 7305c canR argRII--Sl f 123 72 132 67 132 47 7305d canS + lys' 390 36 336 10 114 20
trophic strain MG409, on NHZ and NH: + arginine (1 mg/ml) medium (for more details see [Ill).
One can see that the constitutive derepressed levels of ornithine carbamoyltransferase activity in argR- mutants are far from those obtained when the leaky auxotrophic strain MG409 growing on NH: medium. Of course, this could be because all the argR- muta- tions abolish only partially the argR gene activity. Yet more than 50 different argR- mutants have been isolated [I] and they all give the same ornithine carbamoyltransferase level, and the combination in a haploid strain of two or even the three types of argR- mutations does not change the level of derepression. Also, the fact that argininosuccinase activity, which is not under the control of argR, can be derepressed in the leaky auxotrophic strain MG409, suggests that there are two levels of control on arginine biosynthetic enzymes.
In order to test this hypothesis we looked for new regulatory mutants in the arginine biosynthesis with highly derepressed ornithine carbamoyltransferase activity. As described in Materials and Methods, we isolated all sizes of canavanine-resistant colonies and especially the small ones in order to obtain, if possible, different types of mutants from those we had obtained previously. argR- mutants grow at the same rate as the wild type on NH, medium.
Table 2 shows the characteristics of one of the mutants we obtained: DJ3. It was isolated on NHZ + 200 pg L-ornithine per ml x 8 pg of L-canavanine per ml and has high ornithine carbamoyltransferase levels. When crossed by the wild type it appeared that two mutations were responsible for the high ornithine carbamoyltransferase activity in that mutant : an argRZZI--51 mutation plus a leaky block in one of the lysine biosynthetic enzymes.
Table 3 shows the analysis of one tetrad from the cross of DJ3 by 3962c (wild type a). These results show clearly that the high ornithine carbamoyl- transferase level is obtained only when an argR- mutation is combined with a leaky mutation. When the metabolic block and the growth rate are corrected by addition of lysine, the level of ornithine carbamoyl- transferase drops to that of an argR- mutant.
The fact that a lysine leaky mutation can influence the arginine biosynthetic enzyme levels was not too surprising because when 3-aminotriazole inhibits one of the histidine biosynthetic enzymes (imidazolgly- cerolphosphate dehydratase) and thereby lowers the histidine pool, arginine and tryptophan enzymes are also derepressed [ 5 ] .
The results presented in Table 4 show that a leaky auxotrophic block in any of the pathways histidine, tryptophan, arginine or isoleucine-valine, combined
Eur. J . Biochem. 57 (1975)
J. Delforge, F. Messenguy, and J.-M. Wiame 235
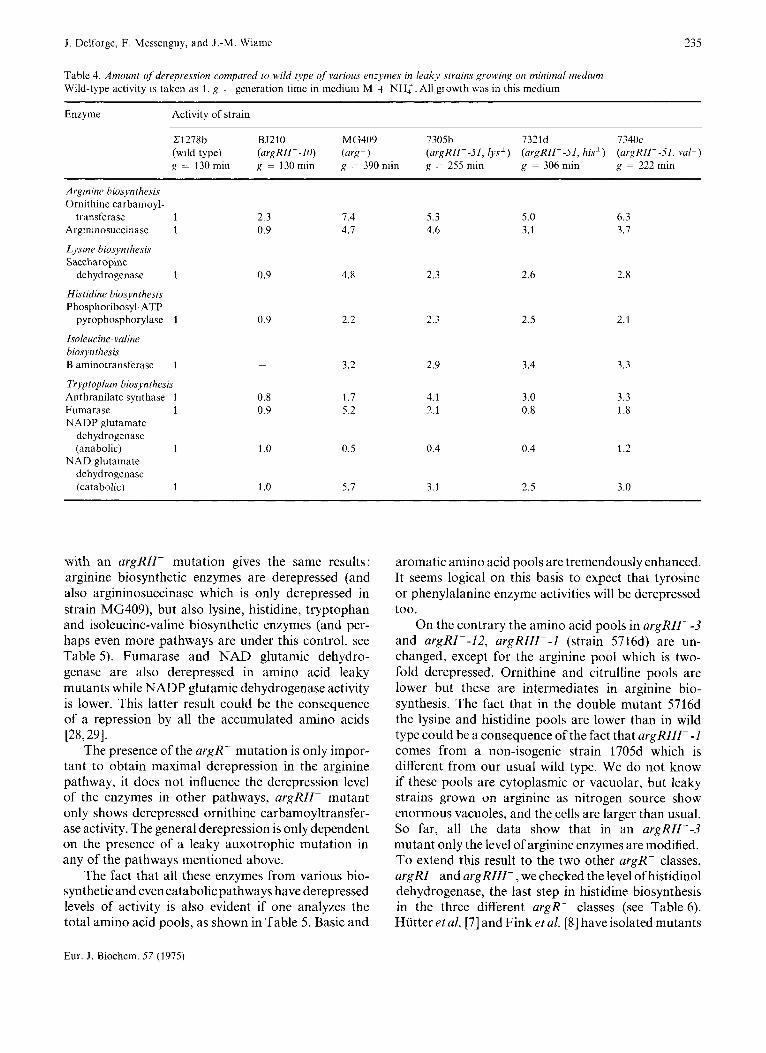
Table 4. Amount of derepression compared to wild type of various enzymes in leaky strains growing on minimal medium Wild-type activity is taken as 1, g = generation time in medium M + NH:. All growth was in this medium
~~
Enzyme Activity of strain
C1278b BJ210 MG409 7305b 7321d 7340c (wild type) (urgRII--lO) (arg') (argRII--51, lys') (argRII--51, his') (argRII--51, Val') g = 130 min g = 130 min g = 390 rnin g = 255 rnin g = 306 rnin g = 222 min
Arginine biosynthesis Ornithine carbamoyl-
transferase 1 2.3 7.4 5.3 5.0 6.3 Argininosuccinase 1 0.9 4.7 4.6 3.1 3.7
Lysine biosynthesi.y Sacchar opine
dehydrogenase 1 0.9 4.8 2.3 2.6 2.8
Histidine biosynthesis Phosphoribosyl-ATP
Isoleucine- valine biosynthesis B aminotransferase 1 - 3.2 2.9 3.4 3.3
Tryptophan biosynthesis
Fumarase 1 0.9 5.2 2.1 0.8 1.8 NADP glutamate
dehydrogenase (anabolic) 1 1 .o 0.5 0.4 0.4 1.2
NAD glutamate dehydrogenase (catabolic) 1 1 .o 5.7 3.1 2.5 3.0
pyrophosphorylase 1 0.9 2.2 2.3 2.5 2.1
Anthranilate synthase 1 0.8 1.7 4.1 3.0 3.3
with an argRII- mutation gives the same results: arginine biosynthetic enzymes are derepressed (and also argininosuccinase which is only derepressed in strain MG409), but also lysine, histidine, tryptophan and isoleucine-valine biosynthetic enzymes (and per- haps even more pathways are under this control, see Table 5). Fumarase and NAD glutamic dehydro- genase are also derepressed in amino acid leaky mutants while NADP glutamic dehydrogenase activity is lower. This latter result could be the consequence of a repression by all the accumulated amino acids [28,29].
The presence of the argR- mutation is only impor- tant to obtain maximal derepression in the arginine pathway, it does not influence the derepression level of the enzymes in other pathways, argRII- mutant only shows derepressed ornithine carbamoyltransfer- ase activity. The general derepression is only dependent on the presence of a leaky auxotrophic mutation in any of the pathways mentioned above.
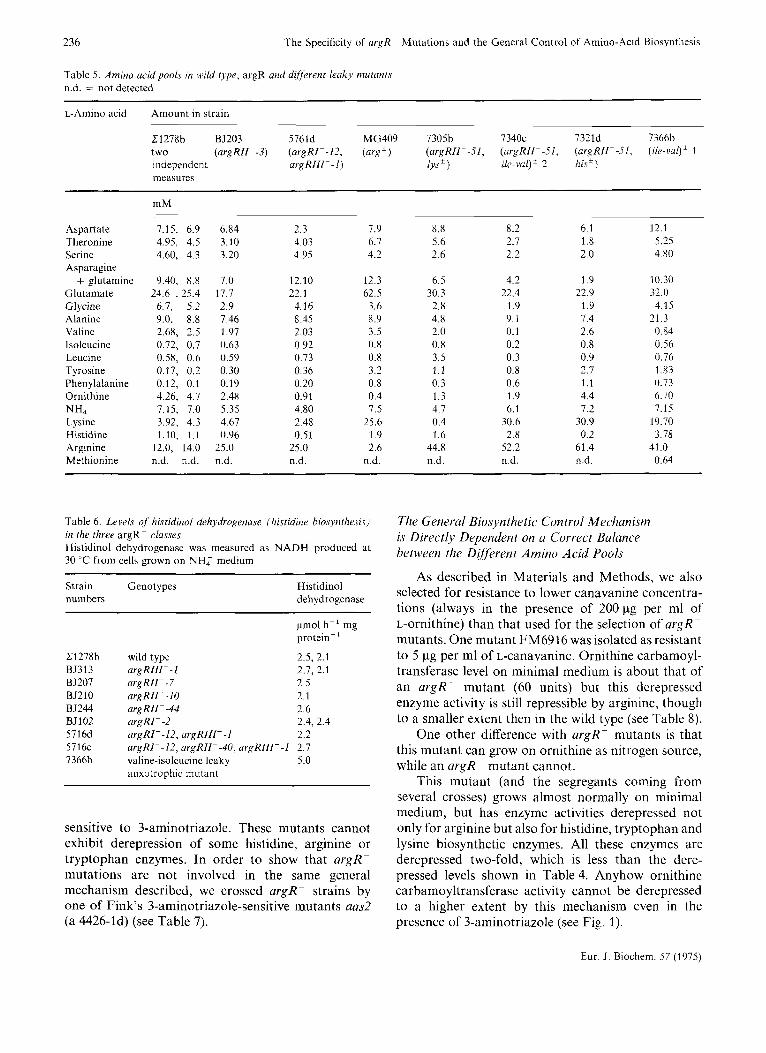
The fact that all these enzymes from various bio- synthetic and even catabolic pathways have derepressed levels of activity is also evident if one analyzes the total amino acid pools, as shown in Table 5. Basic and
aromatic amino acid pools are tremendously enhanced. It seems logical on this basis to expect that tyrosine or phenylalanine enzyme activities will be derepressed too.
On the contrary the amino acid pools in argRII--3 and argRI--12, argRIIIp-I (strain 5716d) are un- changed, except for the arginine pool which is two- fold derepressed. Ornithine and citrulline pools are lower but these are intermediates in arginine bio- synthesis. The fact that in the double mutant 5716d the lysine and histidine pools are lower than in wild type could be a consequence of the fact that argRIII--I comes from a non-isogenic strain 1705d which is different from our usual wild type. We do not know if these pools are cytoplasmic or vacuolar, but leaky strains grown on arginine as nitrogen source show enormous vacuoles, and the cells are larger than usual. So far, all the data show that in an argRII--3 mutant only the level of arginine enzymes are modified. To extend this result to the two other argR- classes, argRI- and argRIII- , we checked the level of histidinol dehydrogenase, the last step in histidine biosynthesis in the three different argR- classes (see Table 6). Hutter et al. [7] and Fink et al. [S] have isolated mutants
Eur. J . Biochem. 57 (1975)
236 The Specificity of argR- Mutations and the General Control of Amino-Acid Biosynthesis
Table 5. Amino arid pools in wild type, argR and different leaky mutants n.d. = not detected
L-Amino acid Amount in strain
C1278h BS203 5761d MG409 7305h 7340c 7321d 7366b two (argRII -3) (urgRIFI2 , (arg') (argRIIF51, (argRIl--51, (argRII--51, ( i l e - v a ~ ' 1 independent at-gRff l - - I) lys ' ile-Val)' 2 his') measures
m M
Aspartate 7.15, 6.9 6.84 2.3 7.9 8.8 8.2 6.1 12.1 Theronine 4.95, 4.5 3.10 4.03 6.7 5.6 2.7 1 .8 5.25 Serine 4.60, 4.3 3.20 4.95 4.2 2.6 2.2 2.0 4.80 Asparagine
+ glutamine 9.40, 8.8 7.0 12.10 12.3 6.5 4.2 1 .9 10.30 Glutamate 24.6 , 25.4 17.7 22.1 62.5 30.3 22.4 22.9 32.0 Glycine 6.7, 5.2 2.9 4.16 3.6 2.8 1.9 1.9 4.15 Alanine 9.0, 8.8 7.46 8.45 8.9 4.8 9.1 1.4 21.3 Valine 2.68, 2.5 1.97 2.03 3.5 2.0 0.1 2.6 0.84 Isoleucine 0.72, 0.7 0.63 0.92 0.8 0.8 0.2 0.8 0.56 Leucine 0.58, 0.6 0.59 0.73 0.8 3.5 0.3 0.9 0.76 Tyrosine 0.17, 0.2 0.30 0.36 3.2 1.1 0.8 2.7 1.83
Ornithine 4.26, 4.7 2.48 0.91 0.4 1.3 1.9 4.4 6.70 NH4 7.15, 7.0 5.35 4.80 1.5 4.7 6.1 7.2 7.15 Lysine 3.92, 4.3 4.61 2.48 25.6 0.4 30.6 30.9 19.70 Histidine 1.10, 1.1 0.96 0.51 I .9 1.6 2.8 0.2 3.78 Arginine 12.0, 14.0 25.0 25.0 2.6 44.8 52.2 61.4 41 .0 Methionine n.d. n.d. n.d. n.d. n.d. n.d. n.d. n.d. 0.64
Phenylalanine 0.12, 0.1 0.19 0.20 0.8 0.3 0.6 1.1 0.73
Table 6. Levels of histidinol dehydrogenase [hislidinr biosynthesis) in the three argR classes Histidinol dehydrogenase was measured as NADH produced at 30 "C from cells grown on NHZ medium
Strain Genotypes Histidinol numbers dehydrogenase
pmol h-' mg protein- '
C1278h wild type 2.5, 2.1 BS313 argRIII- -1 2.7, 2.1 BS201 argRII--7 2.5
BJ244 argRII--44 2.6 BJ102 argRI- -2 2.4, 2.4
5716c argRI--12, argRII--40, urgRl l l - - l 2.1 1366b valine-isoleucine leaky 5.0
BJ210 argRII--10 2.1
5716d argRI- -12, argRIII- -1 2.2
auxotrophic mutant
sensitive to 3-aminotriazole. These mutants cannot exhibit derepression of some histidine, arginine or tryptophan enzymes. In order to show that argR- mutations are not involved in the same general mechanism described, we crossed argR- strains by one of Fink's 3-aminotriazole-sensitive mutants aas2 (a 4426-1d) (see Table 7).
The General Biosynthetic Control Mechanism is Directly Dependent on a Correct Balance between the D ~ > r e n t Amino Acid Pools
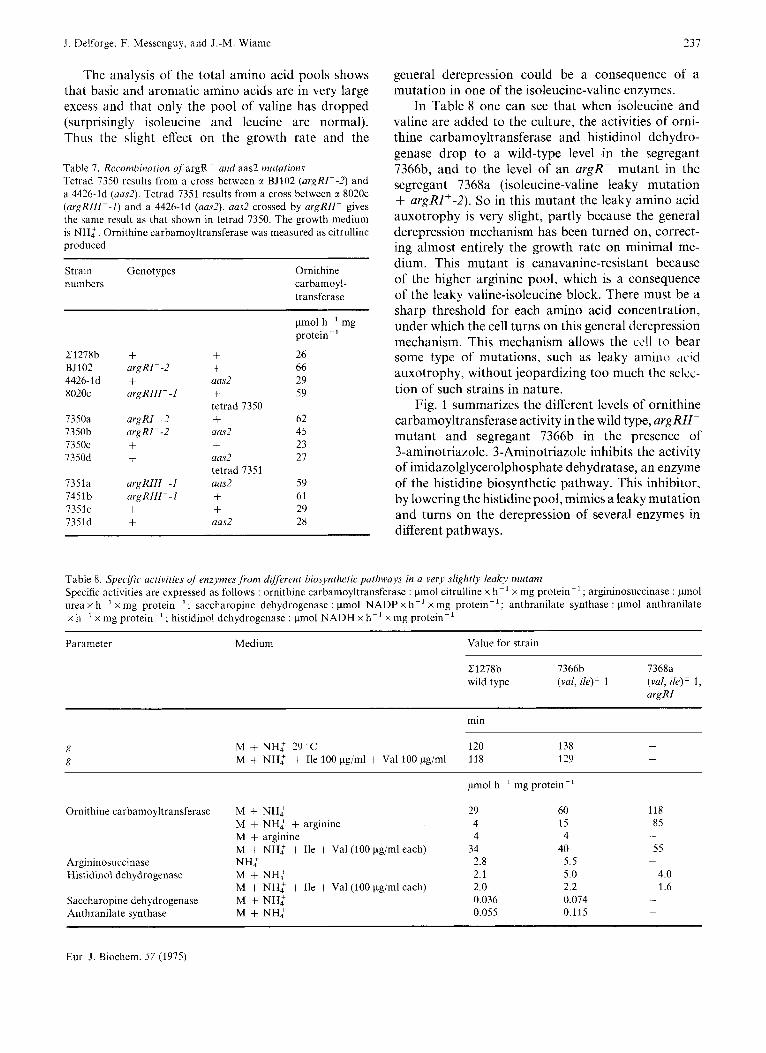
As described in Materials and Methods, we also selected for resistance to lower canavanine concentra- tions (always in the presence of 200pg per ml of L-ornithine) than that used for the selection of argR- mutants. One mutant FM6916 was isolated as resistant to 5 pg per ml of L-canavanine. Ornithine carbamoyl- transferase level on minimal medium is about that of an argR- mutant (60 units) but this derepressed enzyme activity is still repressible by arginine, though to a smaller extent then in the wild type (see Table 8).
One other difference with argR- mutants is that this mutant can grow on ornithine as nitrogen source, while an a r g K mutant cannot.
This mutant (and the segregants coming from several crosses) grows almost normally on minimal medium, but has enzyme activities derepressed not only for arginine but also for histidine, tryptophan and lysine biosynthetic enzymes. All these enzymes are derepressed two-fold, which is less than the dere- pressed levels shown in Table 4. Anyhow ornithine carbamoyltransferase activity cannot be derepressed to a higher extent by this mechanism even in the presence of 3-aminotriazole (see Fig. 1).
Eur. J. Biochem. 57 (1975)
J . Delforge. F. Messenguy, and J.-M. Wiame 237
The analysis of the total amino acid pools shows that basic and aromatic amino acids are in very large excess and that only the pool of valine has dropped (surprisingly isoleucine and leucine are normal). Thus the slight effect on the growth rate and the
Table 7. Recombination of argR- and aas2 mutations Tetrad 7350 results from a cross between c( BJ102 ( u r g I W - 2 ) and a 4426-Id ( a d ) . Tetrad 7351 results from a cross between 01 8020~ ( a r g R l l l ~ - l ) and a 4426-1d (aas2). am2 crossed by argRll- gives the same result as that shown in tetrad 7350. The growth medium is NH: . Ornithine carbamoyltransferase was measured as citrulline produced
Strain Genotypes numbers
Ornithine carbamoyl- transferase
C1278b BJ102 4426-Id 8020c
7350a 7350b 7350c 7350d
7351a 7451b 7351c 7351d
+ argR1- -2 + argR111- - 1
argR1- -2 argR1- -2 + + argRll l - - l urg R l l I - - 1 + +
+ + aas2 + tetrad 7350 + aas2 + aas2 tetrad 7351 a m 2 + + a m 2
pmol h-' mg protein-'
26 66 29 59
62 45 23 27
59 61 29 28
general derepression could be a consequence of a mutation in one of the isoleucine-valine enzymes.
In Table 8 one can see that when isoleucine and valine are added to the culture, the activities of orni- thine carbamoyltransferase and histidinol dehydro- genase drop to a wild-type level in the segregant 7366b, and to the level of an avgR- mutant in the segregant 7368a (isoleucine-valine leaky mutation + argRI+-2). So in this mutant the leaky amino acid auxotrophy is very slight, partly because the general derepression mechanism has been turned on, correct- ing almost entirely the growth rate on minimal me- dium. This mutant is canavanine-resistant because of the higher arginine pool, which is a consequence of the leaky valine-isoleucine block. There must be a sharp threshold for each amino acid concentration, under which the cell turns on this general derepression mechanism. This mechanism allows the ccll to bear some type of mutations, such as leaky amino 'ic.id auxotrophy, without jeopardizing too much the selcc- tion of such strains in nature.
Fig. 1 summarizes the different levels of ornithine carbamoyltransferase activity in the wild type, argRII- mutant and segregant 7366b in the presence of 3-aminotriazole. 3-Aminotriazole inhibits the activity of imidazolglycerolphosphate dehydratase, an enzyme of the histidine biosynthetic pathway. This inhibitor, by lowering the histidine pool, mimics a leaky mutation and turns on the derepression of several enzymes in different pathways.
Table 8. Specific uctivities of enzymesfronz different hiosynthetic putkwuys in a very slightly leaky mutant Specific activities are expressed as follows : ornithine carbamoyltransferase : pmol citrulline x h-' x mg protein-' ; argininosuccinase : pmol urea x h- ' x mg protein-'; saccharopine dehydrogenase : pmol NADP x h-' x mg protein-'; anthranilate synthase : pmol anthranilate x h-' x mg protein-'; histidinol dehydrogenase : pmol NADH x h-' x mg protein-'
Parameter Medium Value for strain
C1278b 7366b 7368a wild type (val, ile)' 1 (Val, ik)' I,
argRl-
min
- M + NH,+ 29 "C 120 138 M + NH: + Ile 100 pg/ml + Val 100 pg/ml 118 129 -
pmol h-' mg protein-'
Ornithine carbamoyltransferase M + NH: M + NH: + arginine M + arginine M + NH: + Ile + Val (100 pg/ml each)
Argininosuccinase NH: Histidinol dehydrogenase M + NH:
Saccharopine dehydrogenase M + NH: Anthranilate synthase M + NH:
M + NH: + Ile + Val (100 pg/ml each)
29 4 4
34 2.8 2.1 2.0 0.036 0.055
60 15 4
40 5.5 5.0 2.2 0.074 0.115
118 85
55 -
-
4.0 1.6
Eur. J. Biochem. 57 (1975)
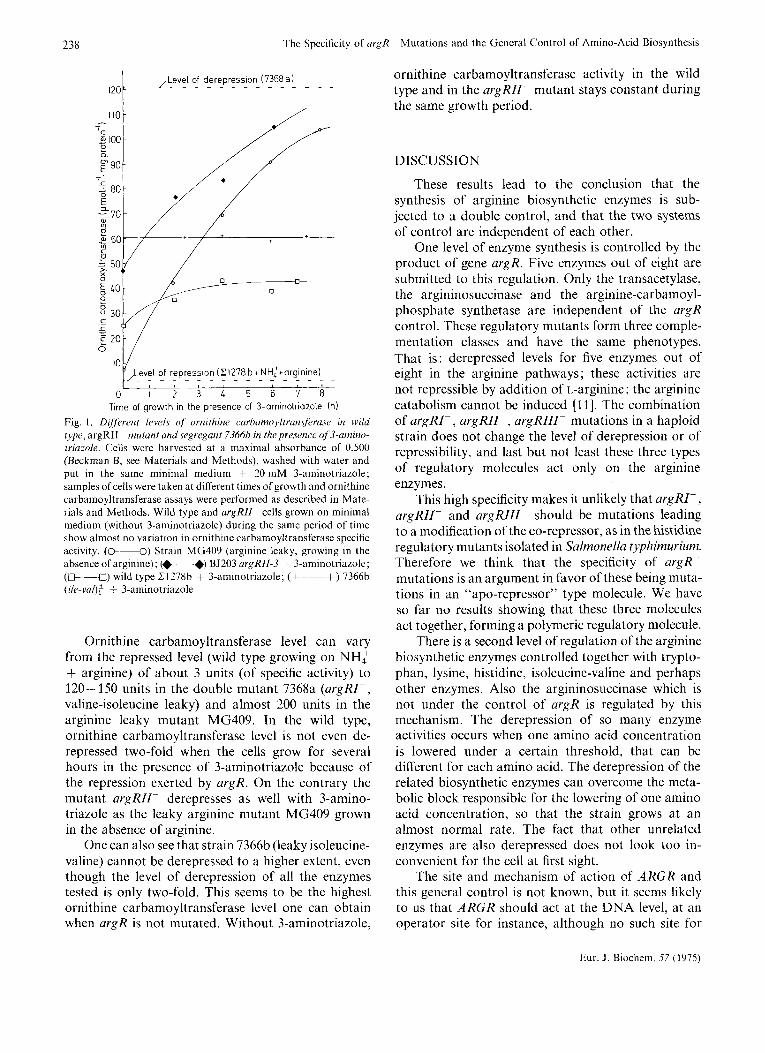
238 The Specificity of argR- Mutations and the General Control of Amino-Acid Biosynthesis
__o-
0
10 Level of repression (El278 b +NHt+arginine) _ _ - _ _ _ _ _ - - - - - - - - -
0 1 2 3 L 5 6 7 8 Time of growth in the presence of 3-aminotriazole (h )
Fig. 1. Dijferetzt levels qf ornithine carbumoyltransjeruse in wild type, argRIl- mutant andsegregant 7366b in the presence oj3-amino- lriazole. Cells were harvested at a maximal absorbance of 0.500 (Beckman B, see Materials and Methods), washed with water and put in the same minimal medium + 20mM 3-aminotriazole; samples ofcells were taken at different times of growth and oruithine carbamoyltransferase assays were performed as described in Mate- rials and Methods. Wild type and argRII- cells grown on minimal medium (without 3-aminotriazole) during the same period of time show almost no variation in ornithine carbamoyltransferase specific activity. ( 0 - 0 ) Strain MG409 (arginine leaky, growing in the absence of arginine): (+ +) BJ203 argRII-3 + 3-aminotriazole; (--a) wild type 21278b + 3-aminotriazole; (+--+) 7366b (ile-var): + 3-aminotriazole
Ornithine carbamoyltransferase level can vary from the repressed level (wild type growing on NH: + arginine) of about 3 units (of specific activity) to 120-150 units in the double mutant 7368a (argRI- , valine-isoleucine leaky) and almost 200 units in the arginine leaky mutant MG409. In the wild type, ornithine carbamoyltransferase level is not even de- repressed two-fold when the cells grow for several hours in the presence of 3-aminotriazole because of the repression exerted by argR. On the contrary the mutant argRII- derepresses as well with 3-amino- triazole as the leaky arginine mutant MG409 grown in the absence of arginine.
One can also see that strain 7366b (leaky isoleucine- valine) cannot be derepressed to a higher extent, even though the level of derepression of all the enzymes tested is only two-fold. This seems to be the highest ornithine carbamoyltransferase level one can obtain when argR is not mutated. Without 3-aminotriazole,
ornithine carbamoyltransferase activity in the wild type and in the argRII- mutant stays constant during the same growth period.
DISCUSSION
These results lead to the conclusion that the synthesis of arginine biosynthetic enzymes is sub- jected to a double control, and that the two systems of control are independent of each other.
One level of enzyme synthesis is controlled by the product of gene argR. Five enzymes out of eight are submitted to this regulation. Only the transacetylase, the argininosuccinase and the arginine-carbamoyl- phosphate synthetase are independent of the argR control. These regulatory mutants form three comple- mentation classes and have the same phenotypes. That is: derepressed levels for five enzymes out of eight in the arginine pathways ; these activities are not repressible by addition of L-arginine ; the arginine catabolism cannot be induced [ll]. The combination of argRI- , argRII-, argRIII- mutations in a haploid strain does not change the level of derepression or of repressibility, and last but not least these three types of regulatory molecules act only on the arginine enzymes.
This high specificity makes it unlikely that argRI- , argRII- and argRIII- should be mutations leading to a modification of the co-repressor, as in the histidine regulatory mutants isolated in Salmonella typhimurium. Therefore we think that the specificity of argR- mutations is an argument in favor of these being muta- tions in an “apo-repressor” type molecule. We have so far no results showing that these three molecules act together, forming a polymeric regulatory molecule.
There is a second level of regulation of the arginine biosynthetic enzymes controlled together with trypto- phan, lysine, histidine, isoleucine-valine and perhaps other enzymes. Also the argininosuccinase which is not under the control of argR is regulated by this mechanism. The derepression of so many enzyme activities occurs when one amino acid concentration is lowered under a certain threshold, that can be different for each amino acid. The derepression of the related biosynthetic enzymes can overcome the meta- bolic block responsible for the lowering of one amino acid concentration, so that the strain grows at an almost normal rate. The fact that other unrelated enzymes are also derepressed does not look too in- convenient for the cell at first sight.
The site and mechanism of action of ARGR and this general control is not known, but it seems likely to us that ARGR should act at the DNA level, at an operator site for instance, although no such site for
Eur. J . Biochem. 57 (1975)

J. Delforge, F. Messenguy, and J.-M. Wiame 239
ARGR action has been determined on the DNA yet. On the other hand the general control seems more appropriate to act at the translational level. For ex- ample, the amount of messenger RNA produced for most of the enzymes in yeast, under growth in minimal medium, might be in excess so that only part of it is translated into the necessary proteins, producing enough amino acids to get into protein.
Therefore, there should exist general regulatory molecules which under normal conditions lower the effect of the message, but if a sharp drop in one amino acid pool appears, this control is turned off, and all the message is read, producing more enzymes, restab- lishing that amino acid pool to a concentration as close as possible to normal. This would then be a safety mechanism which allows the cell to adapt more easily to some stressing conditions.
Studies of operator-like mutants could help in differenciating those two levels of control occurring in the arginine biosynthetic pathway. If several operator mutants have the same, or lower, constitutive levels of enzymes than urgR- mutants, this would suggest that the general mechanism is not likely to act at the same site. Promotor mutants could also be helpful in dissociating these two regulatory mecha- nisms controlling one biosynthetic pathway.
These data also point out one important fact in the isolation of a regulatory mutant by resistance to an analog: we isolated several leaky mutants (even if the growth rate is not much affected) as canavanine- resistant. Ornithine carbamoyltransferase activity was derepressed and less repressible by addition of arginine, and, even on minimal medium, the arginine pool is 3-4-fold higher than the pool in wild type, and yet the mutation which is responsible for these character- istics is unrelated to arginine metabolism. Some of the regulatory mutants reported in the literature have similar phenotypes and might be slightly leaky mu- tants. Thus the specificity of action of a regulatory mutation is a necessary condition for it to be a candi- date as a mutation in an apo-repressor molecule.
We thank Rosette Loriau for her skilful technical assistance. Jacqueline Delforge is a Boursidre de l’lnstitut pour I’Encourage- ment de lu Recherche Scientifique duns I’lndustrie et I’Agriculture. This work was supported by a grant from the Fonds de la Recherche Fondumentule Collective.
REFERENCES
1. Btchet, J., Grenson, M. & Wiame, .1. M. (1970) Eur. J . Biochem.
2. Thuriaux, P., Ramos, F., Pierard, A., Grenson, M. & Wiame,
3. Cherest, H., Eichler, F. & Robichon-Szulmdjster, H. de (1969)
4. Bollon, A. (1974) Nature (Lond.) 250, 630-634. 5 . Carsiotis, M. & Lacy, A. M. (1965) J . Bucteriol. 89, 1472-
6. Carsiotis, M., Jones, R. F., Lacy, A. M., Cleary, T. J. & Fank-
7. Schiirch, A,, Miozzdri, J. & Hiitter, R. (1974) J . Bacteriol. 117,
8. Wolfner, M:, Yep, D., Messenguy, F. & Fink, G. R. (1975)
9. Lewis, J. A. & Ames, B. N. (1972) J . Mol. Biol. 65, 131 - 142. 10. Rizzino, A. A,, Bresdlier, R. S. & Freundlich, M. (1974) J .
Bucteriol. 17, 449 - 455. 11. Wiame, J. M. (1970) in Recent Advances in Microbiology
(Perez-Mizaveta & D. Pilaez, eds) Report to the Xth Con- gress of Microbiology, Mexico, August 1970.
12. Hoet, P. P. & Wiame, J. M. (1974) Eur. J . Biochem. 43, 87-92. 13. Dubois, E., Grenson, M. & Wiame, J. M. (1974) Eur. J . Bio-
14. Grenson, M., Hou, C. & Crabeel, M. (1970) J . Bucteriol. 103,
15. Thuriaux, P., Ramos, F., Wiame, J. M., Grenson, M. & Bechet,
16. Penninckx, M., Simon, J. P. & Wiame, J. M. (1974) Eur. J .
17a. Racker, E. (1950) Biochim. Biophys. Acta, 4, 211-214. 17b. Pollakis, E. S. & Bartley, W. (1965) Biochem. J. 97, 284-297. 18. Fink, G. R. & Styles, C. A. (1974) Genetics 77, 231 -244. 19. Tamir, H. & Srinivasan, P. R. (1970) Methods Enzymol. 17A,
20. Broquist, H. P. (1970) Methods Enzymol. I7B, 124-129. 21. Kovach, J. S., Berberich, M. A,, Venetianer, P. & Goldberger,
R. F. (1969) J . Bucteriol. 97, 1283-1290. 22. Boehringer, C. F. (1968) Biochimica Catalogue, Boehringer
Mannheim GmbH, Mannheim, Germany. 23. Boehringer, C. F. (1968) Biochemical Catalogue, Boehringer
Mannheim GmbH, Mannheim, Germany. 24. Ratner, S. (1955) Methods Enzymol. 2, 356-367. 25. Jenkins, W. T. & Taylor, R. T. (1970) Methods Enzymol. 17A,
26. Lowry, 0. H., Rosenbrough, N. J., Farr, L. & Randall, R. J.
27. Moore, S.&Stein, W. H.(1963)MethodsEnzymol. 6,819-831. 28. Thomulka, K. W. & Moat, A. G. (1972) J . Bucteriol. 109,
29. Drillien, R., Aigle, M. & Lacroute, F. (1973) Biochem. Biophys.
30. Ramos, F., Thuriaux, P., Wiame, J. M. & Bkhet, J. (1970)
I2,31- 39.
J. M. (1972) J . Mol. Biol. 67, 277-287.
J . Bucteriol. 97, 328 - 336.
1477.
hauser, D. B. (1970) J . Bacteriol. 104, 98-106.
1131-1140.
J . Mol. Bid . Submitted for publication.
chem. 48,603-616.
770- 771.
J. (1968) Arch. Int. Physiol. Biochem. 76, 955-956.
Biochem.9.429 - 442.
401-406.
802- 807.
(1951) J . Biol. Chem. 193, 265-275.
25-33.
Res. Commun. 53,367- 372.
Eur. J . Biochem. 12,40-47.
J. Delforge, F. Messenguy, and J.-M. Wiame, Institut de Recherches du C.E.R.I.A., Avenue Emile-Gryzon 1, B-1070 Bruxelles, Belgium
Eur. J. Biochem. 57 (1975)


![Mutations in the Pectin Methyltransferase QUASIMODO2 ...Mutations in the Pectin Methyltransferase QUASIMODO2 Influence Cellulose Biosynthesis and Wall Integrity in Arabidopsis[OPEN]](https://img.pdfslide.fr/doc/110x75/609d35a4bb8ab77aa43faa7a/mutations-in-the-pectin-methyltransferase-quasimodo2-mutations-in-the-pectin.jpg)









![Extension of 2D FEniCS implementation of Cosserat non ... · The objective of the internship is the extension of the existing 2D FEniCS implementation of Cosserat elasticity [9] to](https://img.pdfslide.fr/doc/110x75/604d6997ec52f606395b1501/extension-of-2d-fenics-implementation-of-cosserat-non-the-objective-of-the-internship.jpg)
















