Embed Size (px)
Citation preview

Topology of Ionotropic GlutamateReceptors in Brains of Heterozygous and
Homozygous Weaver Mutant MiceTOMAS A. READER* AND JACQUES SENECAL
Centre de recherche en sciences neurologiques, Departement de physiologie, Faculte de medecine,Universite de Montreal, Montreal, Quebec, H3C 3J7 Canada
KEY WORDS NMDA; MK-801; AMPA; kainic acid; autoradiography
ABSTRACT In weaver mice, mutation of a G-protein inwardly rectifying K1 channelleads to a cerebellar developmental anomaly characterized by granule and Purkinje cellloss and, in addition, degeneration of dopaminergic neurons. To evaluate other deficits,ionotropic glutamate receptors sensitive to N-methyl-D-aspartate (NMDA), amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA), and kainic acid (KA) were exam-ined by autoradiography with [3H]MK-801, [3H]AMPA, and [3H]KA. These surveys werecarried out in selected areas of cerebral cortex, hippocampus and related limbic regions,basal ganglia, thalamus, hypothalamus, brainstem, and cerebellum from heterozygous(wv/1) and homozygous (wv/wv) weaver mutants, and compared to wild-type (1/1) mice.In wv/1 and wv/wv mutants, NMDA receptor levels were lower in cortical areas, septum,hippocampus, subiculum, neostriatum, nucleus accumbens, superior colliculus, and inthe cerebellar granular layer. Densities of KA receptors were lower in cortical areas,hippocampus, limbic system structures, neostriatum, nucleus accumbens, thalamus andhypothalamus, superior and inferior colliculi, and cerebellar cortex of wv/wv mutants.Levels of AMPA receptors in the weaver were higher than in 1/1 mice, particularly insomatosensory and piriform cortices and periaqueductal gray of wv/1, and in somato-sensory cortex, CA1 field of Ammon’s horn and cerebellar granular layer of wv/wv.Abnormal developmental signals, aberrant cellular responses, or a distorted balancebetween neurotransmitter interactions may underlie such widespread and reciprocalglutamate receptor alterations, while in the case of cerebellar cortex, NMDA receptorsare lacking due to a massive disappearance of cerebellar granule cells and some loss ofPurkinje neurons. Synapse 42:213–233, 2001. © 2001 Wiley-Liss, Inc.
INTRODUCTION
The weaver is a naturally occurring autosomal reces-sive mutation in mice (Sidman, 1968) and the homozy-gous (wv/wv) phenotype displays severe neurologicalimpairments, including ataxia, tremor, and poor limbcoordination (Lalonde, 1986, 1987; Lalonde and Botez,1986; Triarhou and Ghetti, 1987). As a consequence ofa postnatal loss of granule cells (Rakic and Sidman,1973a,b), defective neuron migration (Rezai and Yoon,1972; Rakic and Sidman, 1973c), and abnormal devel-opment of Purkinje cell dendrites (Sotelo, 1975) thecerebellum of wv/wv mutants is extremely atrophied.Failure of granule cell migration and neuronal deathseem to be linked; however, some granule cells die evenafter successful postnatal migration (Sotelo and Chan-geux, 1974). In addition, there is a progressive degen-eration of dopamine (DA)-containing neurons (Lane etal., 1977; Schmidt et al., 1982; Roffler-Tarlov and Gray-
biel, 1984); thus, the weaver behavioral phenotype (Tri-arhou and Ghetti, 1987; Lalonde et al., 1988) is theoutcome of an agranular cerebellum combined to a DAdeficit (Gupta et al., 1987; Triarhou et al., 1988). Inwv/wv mutants, about 50% of DA neurons of the sub-stantia nigra have degenerated by postnatal day 20–21(Verney et al., 1995; Verina et al., 1997), and it hasbeen linked to their time of origin (Bayer et al., 1995).As expected, tissue DA and densities of DA transport-ers are diminished (Lane et al., 1977; Schmidt et al.,1982; Roffler-Tarlov and Graybiel, 1984; Richter et al.,
Contract grant sponsor: Canadian Institutes of Health Research; Contractgrant number: MOP 38094; Contract grant sponsor: Reseau sante mentale duQuebec, Fonds de la recherche en sante du Quebec.
*Correspondence to: Dr. Tomas A. Reader, Departement de physiologie, Fac-ulte de medecine, Universite de Montreal, C.P. 6128 Succursale Centre-Ville,Montreal (Quebec) H3C 3J7 Canada. E-mail: [email protected]
Received 21 May 2001; Accepted 3 August 2001
Published online 00 Month 2001; DOI 10.1002/syn.10007
SYNAPSE 42:213–233 (2001)
© 2001 WILEY-LISS, INC.

1992; Strazielle et al., 1998), particularly in dorsalneostriatum (Rofler-Tarlov and Graybiel, 1987; Readeret al., 1999), a region with elevated serotonin (5-HT)content (Stotz et al., 1993) and hyperinnervated, asevidenced by 5-HT transporter topology (Dewar, 1999;Reader et al., 2001), probably a consequence of theextensive and very gradual postnatal loss of DA pro-jections (Reader and Dewar, 1999).
The weaver mutation was first mapped to chromo-some 16 in mouse (Reeves et al., 1989), and then shownto be a single amino acid substitution in a G-proteininwardly rectifying K1 channel, or GIRK2 (Mjaatvedtet al., 1995; Patil et al., 1995). The abnormal wvGIRK2protein assembles into homomultimeric channels thatlose the normal higher selectivity for K1 over Na1 andgive rise to basally active Na1 currents, leading to apoor resting membrane potential. The increased Na1
permeability and ensuing depolarizations could con-tribute to the pathology of cerebellar granule cells andnigral DA neurons (Kofuji et al., 1996; Tong et al.,1996). Also, there could be a loss of GIRK2-mediatedcurrents (Surmeier et al., 1996); the presence of het-eromultimers of GIRK2-weaver and GIRK1-wild typeappear to have reduced currents caused by nonselec-tive channel opening (Slesinger et al., 1996; Navarro etal., 1996). The aberrant behavior of mutated wvGIRK2channels has been considered as a molecular gain-of-function mutant (Lester and Karshin, 2000). Anothergain-of-function mutant is the Lurcher (Lc/1) mouse,the resultant of a mutated d2 gene (Lc d2) encoding aputative glutamate receptor subunit (Zuo et al., 1997)that represents a constitutively active cation channel,predominantly found in shafts and spines of Purkinjeneuron dendrites and in climbing fiber synapses in theearly postnatal period (Zhao et al., 1998). The Lc d2mutation produces constitutively active ionotropic re-ceptors sensitive to a-amino-3-hydroxy-5-methylisox-azole-4-propionic acid acid (AMPA) and to kainic acid(KA), and although those sensitive to N-methyl-D-as-partic acid (NMDA) did not exhibit constitutive activa-tion, they revealed delayed inactivation in response toagonist drugs (Lester and Karschin, 2000). In Lc/1mice, derived from the same wild-type strain as theweaver, there are profound alterations in the topologyof several glutamate receptors that were documentedin cerebellum, as expected, but also in other brainstructures including cerebral cortex, limbic system,and basal ganglia (Strazielle et al., 2000).
The gain-of-function of the mutated wvGIRK2 chan-nel leads to a greater susceptibility to drugs that arethought to block cation channels binding directlywithin the lumen, including the classical compoundverapamil, that acts on L-type Ca21 channels, but alsothe antagonist MK-801 that is known to block NMDAreceptor channels (Kofuji et al., 1996). In the presentstudy, the topologies of ionotropic glutamate receptorsof the NMDA, KA, and AMPA subtypes were examined
in discrete brain regions of heterozygous (wv/1) andhomozygous (wv/wv) mutants and the distributionscompared to that observed in wild-type (1/1) mice.These surveys were carried out by quantitative ligandbinding autoradiography using the antagonist dizo-cilpine, or [3H]MK-801, to label NMDA receptor channels,and the agonists [3H]kainic acid and [3H]a-amino-3-hydroxy-5-methylisoxazole-4-propionic acid for, respec-tively, KA and AMPA sites. A particular emphasis wasgiven here to the hippocampus, where glutamate re-ceptors are expressed in high levels, to the neostria-tum, since it is the main site of the DA deficiency inweaver mutants, and to the cerebellum, which displaysmajor granule cell losses, defective neuronal migration,and abnormal development as well as some loss ofPurkinje neurons.
MATERIALS AND METHODSAnimals and tissue preparation
All animal procedures were in strict accordance withthe Canadian Council on Animal Care Guide to theCare and Use of Experimental Animals (2nd edition),and the protocol was approved by the Comite de deon-tologie pour l’experimentation sur des animaux fromthe Universite de Montreal. The study was carried outwith six male mice from each of the three genotypes(1/1, wv/1, wv/wv), originated from the B6CBACa/astrain, purchased from the Jackson Laboratory (BarHarbor, ME). The animals were all killed by decapita-tion at about 3 months of age (85–115 days), theirbrains quickly removed, frozen in N-methylbutanecooled to 240°C with liquid nitrogen, and stored at280°C until used. Serial coronal sections 20-mm thickwere obtained with a cryostat, mounted on gelatin/chromium potassium sulfate-coated slides, and kept at280°C. On each slide there were 12–16 sections, cor-responding to consecutive coronal planes at about 500–600 mm intervals, so that an entire mouse brain couldbe observed and sampled in a single slide.
Quantitative ligand binding autoradiography
The slides for autoradiography, i.e., three slides peranimal and per assay, of which two were used for totalbinding and one for nonspecific labeling, were broughtto room temperature, preincubated in a Tris-HCl or aTris-acetate buffer for 15–30 min at 0–4°C (ice bath) toremove endogenous ligands, and then blown dry undera stream of air at room temperature. The incubationswith the different ligands were carried out followingprotocols described in detail previously (Dure andYoung, 1995; Greenamyre et al., 1985; Reader andSenecal, 2001; Strazielle et al., 2000). Briefly, theNMDA receptors were labeled with 10 nM of[3H]MK801 (21.7 Ci/mmol; NEN/DuPont, Boston, MA)for 120 min at room temperature in Tris acetate buffer(pH 7.4), containing in addition 100 mM of glutamic
214 T.A. READER AND J. SENECAL

acid and 100 mM of glycine; nonspecific labeling wasdetermined in the presence of 10 mM nonradioactiveMK801 (Sigma Chemical Co., St. Louis, MO). ForAMPA receptors, binding was carried out for 45 min at2–4°C with a 20-nM concentration of [3H]AMPA (a-amino-3-hydroxy-5-methylisoxazole-4-propionic acid,52.3 Ci/mmol; NEN/DuPont) in a 50 mM Tris-HClbuffer (pH 7.2) containing 2.5 mM CaCl2 and 30 mMKSCN. Nonspecific binding was determined in thepresence of 1 mM unlabeled L-glutamic acid (Sigma).KA receptors were labeled in a 50 mM Tris acetatebuffer (pH 7.4) with 37 nM of [3H]kainic acid (58.0Ci/mmol; NEN/DuPont) and the incubations were of 45min at 2–4°C; nonspecific binding was determined inthe presence of 100 mM unlabeled kainic acid (Sigma).After the incubations, the slides were washed twice inice-cold buffer, rinsed rapidly in a cold distilled water,and blown dry with cold air. The dried slides and tritiumstandards ([3H]Microscales™; Amersham, ArlingtonHeights, IL) were placed in autoradiographic cassetteswith tritium-sensitive [3H]Hyperfilm™ (Amersham).The cassettes were stored at 4°C and the films exposedfor 2–3 weeks, developed, fixed, and dried. The slideswith the tissue sections used in the incubations werefixed with 4% paraformaldehyde and stained withcresyl-violet (Nissl) to verify tissue integrity and theanatomical localization of the regions of interest.
Image analysis and statistics
Densitometry measurements of the films were car-ried out with an MCID™ image analysys system (Im-aging Research, St-Catharines, Ontario). Standardcurves, generated from the [3H]Microscales, were usedto convert the optical density (OD) values into receptordensities expressed in femtomoles per milligram of pro-tein (Reader and Strazielle, 1999; Unnerstall et al.,1982). The anatomical structures were defined follow-ing stereotaxic coordinates (Franklin and Paxinos,1997) and multiple OD measurements were taken fromeach region, depending on the extent of the structuresconsidered and their possible heterogeneity. Specificbinding was determined by digitally subtracting theimage of nonspecific labelling (channel 2) from the im-age of total binding (channel 1), and storing the imageof specific binding in channel 3. In the case of thecerebellum, to maximize the precision in the localiza-tion of the molecular and granule cell layers, the chan-nels containing the digitized image of specific bindingand the image of the cresyl-violet stained section (chan-nel 4) were linked; this allowed placement of the sam-pling tool on the image of the Nissl-stained section,while the actual OD readings were from the same areaon the specific binding image that was stored in chan-nel 3 (Hebert et al., 2001). The binding data is reportedas the mean values 6 SEM in femtomoles per milli-gram of protein (fmol/mg protein). Statistical compar-isons were carried out by one-way ANOVA, followed by
Fisher’s F-distribution test (Barlow, 1983; Frank andAlthoen, 1994).
RESULTSNMDA receptors
The distribution of [3H]MK-801 binding to NMDAreceptors in 1/1 mice was ubiquitous (Fig. 1) andheterogenous; indeed, a 20-fold difference was mea-sured (Table I) between the regions of lowest (cerebel-lar molecular layer) as opposed to the highest labeling(hippocampus CA1 field). In cortical areas of 1/1 mice,[3H]MK-801 binding levels were highest in orbital pre-frontal (MO), anterior cingulate (Cg1), primary motor(M1), primary somatosensory (S1), and visual (Vis),while slightly lower values were observed in the othercortices (Fig. 1A,D,G). In some areas, notably in M1,S1, and piriform (Pir), labeling was somewhat greaterin the outer layers (M1o, S1o, and Piro) than in theinner ones (M1i, S1i, Piri). In weaver mutants, corticalNMDA receptor levels were overall lower and the dif-ferences with 1/1 were greater in heterozygotes thanin homozygotes. In wv/1 mutants, [3H]MK-801 bindingwas lower in all 18 cortical areas examined (Fig.1B,E,H) and the levels compared to 1/1 miceamounted to 231% in MO, 226% in Cg1, 222% in M1,224% in M1o, 228% in M1i, 223% in secondary motor(M2), 220% in agranular insular (AI), 225% in S1,220% in S1o, 222% in S1i, 223% in secondary somato-sensory (S2), 223% in Pir, 225% in Piro, 224% in Piri,220% in retrosplenial granular (RSG), 215% in lateralentorhinal (LEnt), and 218% in both Vis and auditory(Au) areas. In wv/wv mutants, [3H]MK-801 bindingdensities were also lower throughout all cortical areas(Fig. 1C,F,I) but to a lesser degree than in wv/1; thesedifferences were significant compared to 1/1 mice inonly 13 of the areas, namely, MO (226%), Cg1 (222%),M1 (219%), M1o (223%), M1i (226%), M2 (222%), S1(218%), S1i (221%), Piro (221%), Piri (220%), RSG(220%), Vis (217%), and Au (215%).
The overall highest [3H]MK-801 labeling was in hip-pocampus (Fig. 1D,G) that exhibited a characteristicpattern. In 1/1 mice, receptor densities were greaterin dentate gyrus (DG) and Ammon’s horn field 1 (CA1)than in CA2 and CA3 fields (F3,20 5 5.846). Bindingin DG was 29% higher than in CA2 (F1,10 5 5.702;P , 0.05) and 44% greater than in CA3 (F1,10 5 10.477;P , 0.01), while in CA1 it was 31% higher than inCA2 (F1,10 5 6.541; P , 0.05), and 47% greaterthan in CA3 (F1,10 5 11.605; P , 0.01). In CA1 thehighest labeling was observed at the termination of theSchaffer-collateral/commissural pathway in stratumradiatum as well as in the granule cell layer. Thelabeling was also intense in stratum oriens (Or), butvery low in the subiculum (Sub), or isthmus of graymatter that connects the hippocampus with the RSGcortex. In wv/1 mutants (Fig. 1E,H), lower bindinglevels altered this pattern, particularly in the DG
IONOTROPIC GLUTAMATE RECEPTORS IN WEAVER MICE 215

Fig. 1
216 T.A. READER AND J. SENECAL

(225%; P , 0.001), CA1 (225%; P , 0.01) and CA3(220%; P , 0.05). The distribution was heterogenous(F3,20 5 7.794), with the highest densities in DG andCA1; indeed, in spite of lower labeling, binding in DGwas 35% higher compared to CA3 (F1,10 5 16.333;P , 0.01), and in CA1 it was 37% greater than in CA3(F1,10 5 5.459; P , 0.05). There was also a 14%decrease (P , 0.05) of [3H]MK-801 binding in Sub ofwv/1 mutants compared to 1/1 mice. In wv/wv mu-tants, labeling was lower than in 1/1 mice throughoutthe hippocampus (Fig. 1F,I); these differences weresignificant only in DG and CA1, amounting to, respec-tively, 216% (P , 0.01) and 219% (P , 0.05).Thus, the intrahippocampal NMDA receptor distribu-tion in wv/wv was more homogenous and labeling in DGwas 34% higher than in CA3 (F1,10 5 5.202; P , 0.05).Finally, in Sub of wv/wv mutants NMDA receptor den-sities were lower (216%; P , 0.05) compared to 1/1mice. In other structures related to the limbic system,such as olfactory tubercle (Tu), septum (SPT), andamygdala (Amyg), [3H]MK-801 binding amounted to400–470 fmol/mg protein and NMDA receptor levelswere lower only in SPT of wv/1 (238%; P , 0.001)and wv/wv (233%; P , 0.01) mutants compared to1/1 mice.
Three structures pertaining to the basal gangliawere studied (Table I), namely, the neostriatum, orcaudate-putamen (CPu), the lateral globus pallidus(LGP), and nucleus accumbens (Acb). In 1/1 mice,[3H]MK-801 binding was about 3-fold higher in CPuand Acb than in LGP. To obtain a more detailed topol-ogy, the neostriatum was divided, using the anteriorcommissure as a landmark, into four rostral quadrants(rNS) and two caudal (cNS) halves. Measurements inthese divisions revealed some heterogeneity (F5,30 5
7.708); binding in rNSvm was 14% higher than inrNSdm (F1,10 5 13.696; P , 0.01) and 22% greaterthan in rNSdl (F1,10 5 29.400; P , 0.001), whilelabeling in rNSvl was 15% higher than in rNSdl(F1,10 5 15.000; P , 0.01). In wv/1 mutants (Fig.1B), compared to 1/1 mice, labeling was lower in CPu(227%), rNSvm (221%), cNSd (225%), cNSv (223%),and Acb (223%). In wv/wv mutants, lower binding inCPu (225%) could be attributed to changes in three ofthe rostral quadrants (Fig. 1C), i.e., rNSdl (222%),rNvm (226%), and rNSvl (219%) and there was atrend in cNS. In Acb of the wv/wv mutants, NMDAreceptor levels were also lower (218%).
In thalamus, NMDA receptors were measured in theanterior nuclei (ThalA), while the main thalamic en-semble of nuclei was divided in latero-dorsal (ThalLD),ventro-lateral (ThalVL), medio-dorsal (ThalMD), andmedio-ventral (ThalMV) quadrants (Fig. 1D). Overall,labeling of thalamus and hypothalamus (Hyp) inweaver mutants was slightly lower than in 1/1 mice(Fig. 1E,F), but these differences were not significant.
In brainstem, NMDA receptors are sparse and fewstructures were surveyed; only in superior colliculus(CS) of wv/wv mutants [3H]MK-801 binding densitieswere lower (228%) than in 1/1 mice. In cerebellumthere was a well-defined topology of labeling (Figs. 1J,4A), with high densities in the granular layer (CBgran).An ANOVA comparison of receptor levels in molecular(CBmol), CBgran, and deep cerebellar nuclei (CBnuc)of 1/1 mice confirmed this heterogeneity (F2,15 546.685), revealing 5-fold higher [3H]MK-801 bindinglevels in CBgran than in CBmol (P , 0.001), and3-fold greater than in CBnuc (P , 0.001). In wv/1mutants, labeling was slightly lower (Figs. 1K, 4B);however, these differences were not significant and the
Fig. 1. Autoradiograms of [3H]MK-801 binding in the forebrain andcerebellum of a wild-type mouse (1/1; A,D,G,J), and from heterozy-gous (wv/1; B,E,H,K) and homozygous (wv/wv; C,F,I,L) weaver mu-tants. A: Section obtained from the rostral forebrain of a 1/1 mouse,showing labeling of NMDA receptors in the cingulate (Cg1), primarymotor (M1), secondary motor (M2), primary somatosensory (S1), sec-ondary somatosensory (S2), agranular insular (AI), and piriform (Pir)cortices. Labeling is also quite distinct in the neostriatum (CPu;caudate-putamen) and in the lateral septum (SPT). B: Section at anequivalent plane from a wv/1 mutant, showing lower NMDA receptordensities in Cg1, M1o and M1i, M2, S1o and S1i, S2, Piro and Piri, andAI cortices as well as in the ventro-medial quadrant of rostral neo-striatum (rNSvm) and SPT. C: In a wv/wv mutant, [3H]MK-801 bind-ing levels were lower in Cg1, M1o and M1i, M2, S1i, Piro and Piricortices as well as in SPT and in three quadrants of rostral neostria-tum: rNSdl, rNSvm and rNSvl. D: Section from a 1/1 mouse showingthe dorsal hippocampus with intense labeling in the dentate gyrus(DG) and in the fields of Ammon’s horn (CA1–CA3), particularly instratum oriens (Or). In the CA1 field labelling is higher than inCA2–CA3. Also note the labeling in amygdala (Amyg) and in the mainmass of the thalamus (Thal), that at this level was divided in quad-rants (see Table I), as well as the hypothalamus (Hyp); the lattershowed a very sparse and uniform labeling. E: In an equivalentsection from a wv/1 mutant, [3H]MK-801 binding levels were lower inDG, CA1, and CA3 as well as in the lateral entorhinal area (LEnt),S1o, S1i, Pir and retrosplenial granular (RSG) cortices. F: In thehippocampus of a wv/wv mutant, densities of NMDA receptors arealso lower than in 1/1 mice, particularly in RSG, S1i, Pir, DG, and
CA1. G: This transverse section through the caudalmost portion ofthalamus and ventral hippocampus illustrates the intense labeling inOr, as well as binding in RSG, visual (Vis), and auditory (Au) cortices.H: In a section at this level from a wv/1 mutant, note lower bindingin RSG, Vis, and Au cortices as well as in the DG, CA1, and subiculum(Sub). I: Similar lower NMDA receptor levels are seen in RSG, Vis,Au, DG, CA1, and Sub from a wv/wv mutant. J: This section travers-ing the brainstem and cerebellum of a 1/1 mouse comprises thecerebellum and medulla oblongata. Note the defined, albeit discrete,[3H]MK-801 binding in the granular layer (CBgran) and the veryscant labeling in the molecular layer (CBmol); both layers were alsoconsidered as total cerebellar cortex (CBctx) by measuring severalentire lobules at each level of section. K: In the brainstem and cere-bellum of a wv/1 mutant, binding is still identifiable in both CBgranand CBmol layers. L: The caudalmost section of the brainstem andcerebellum from a wv/wv mutant exhibits the considerable and char-acteristic atrophy of the latter structure. In spite of the reductions inareas, the cerebellar cortex with its CBmol and CBgran layers canstill be identified; binding levels in Cbgran are lower than in 1/1 miceand wv/1 mutants. In this and in the following figures, the nontrans-formed autoradiographic images digitized at a resolution of about 20mm/pixel were exported as TIFF files and for the illustrations wereresampled at a resolution of 300 3 300 dpi. The interaural (IA) planesand the anatomical nomenclature, sometimes simplified, are from themouse stereotaxic atlas of Franklin and Paxinos (1997). The calibra-tion standards for [3H]MK-801 binding are in femtomoles per milli-gram of protein (20–2,898 fmol/mg P) and the horizontal bar corre-sponds to 1 mm.
IONOTROPIC GLUTAMATE RECEPTORS IN WEAVER MICE 217

receptor gradient was as for 1/1 mice (F2,15 521.709). Indeed, in CBgran layer of wv/1, [3H]MK-801binding was more than 3-fold higher than in CBmoland CBnuc (P , 0.001). In wv/wv mutants there wasan important decrease of NMDA receptor densities inthe greatly atrophied CBgran layer (Figs. 1L, 4C); thisrelative reduction was of 244% and 232% compared tothe levels measured in 1/1 mice and wv/1 mutants,respectively. The overall NMDA receptor topology wasconserved (F2,15 5 11.066) in wv/wv mutants and, inspite of lower [3H]MK-801 binding, relative receptor
levels in CBgran layer were more than 3 times higherthan in CBmol layer (P , 0.01) and 75% higher thanin the CBnuc (P , 0.05).
AMPA receptors
The distribution of [3H]AMPA binding (Fig. 2) wasalso ubiquitous and heterogenous and in 1/1 micethere was a 14-fold difference in receptor levels (TableII) between the regions of lowest (cerebellar molecularlayer) as opposed to the highest binding densities (hip-pocampus CA1 field). In cortical areas of 1/1 mice, the
TABLE I. Distribution of [3H]MK-801 binding sites in brain regions of wild-type (1/1) and in heterozygote (wv/1)and homozygote (wv/wv) weaver mutant mice1
Region 1/1 wv/1 wv/wv F values
Cortical areasOrbital prefrontal MO 606 6 49 417 6 25*** 446 6 20** F2,14 5 9.570Anterior cingulate Cg1 541 6 28 398 6 37** 422 6 27* F2,15 5 6.109Primary motor M1 515 6 16 401 6 28** 419 6 21** F2,15 5 7.481Primary motor, outer M1o 577 6 36 438 6 51* 444 6 29* F2,15 5 3.953Primary motor, inner M1i 445 6 26 319 6 18** 328 6 28** F2,15 5 8.300Secondary motor M2 515 6 26 398 6 30** 401 6 13** F2,15 5 7.678Agranular insular AI 517 6 30 413 6 26* 453 6 22 F2,15 5 4.041Primary somatosensory S1 505 6 24 380 6 33** 415 6 14* F2,15 5 6.512Primary somatosensory, outer S1o 538 6 36 433 6 35* 440 6 30 F2,15 5 3.021Primary somatosensory, inner S1i 418 6 26 328 6 19* 331 6 22* F2,15 5 5.082Secondary somatosensory S2 499 6 36 384 6 36* 418 6 16 F2,15 5 3.742Piriform Pir 481 6 23 369 6 34* 451 6 27 F2,15 5 4.077Piriform, outer Piro 470 6 21 354 6 38** 371 6 15* F2,15 5 5.439Piriform, inner Piri 421 6 22 321 6 26** 336 6 12** F2,15 5 6.872Retrosplenial granular RSG 442 6 15 356 6 16* 356 6 31* F2,15 5 4.955Lateral entorhinal LEnt 494 6 19 420 6 26* 465 6 25 F2,13 5 2.636Visual Vis 543 6 23 447 6 17** 450 6 28* F2,15 5 5.667Auditory Au 519 6 30 427 6 12* 440 6 27* F2,15 5 4.097
HippocampusDentate gyrus DG 993 6 29 746 6 36*** 836 6 58** F2,15 5 8.463Ammon’s horn, field 1 CA1 1009 6 121 757 6 31** 821 6 73* F2,15 5 5.906Ammon’s horn, field 2 CA2 768 6 26 665 6 36 697 6 81 F2,15 5 0.969Ammon’s horn, field 3 CA3 688 6 40 554 6 31* 624 6 45 F2,15 5 2.937Stratum oriens Or 991 6 61 783 6 47 782 6 64 F2,15 5 4.303Subiculum Sub 386 6 15 332 6 19* 326 6 13* F2,15 5 4.374
Limbic systemOlfactory tubercle Tu 400 6 24 348 6 39 394 6 39 F2,15 5 0.656Septum SPT 402 6 21 251 6 27*** 268 6 22** F2,14 5 11.236Amygdala Amyg 466 6 12 410 6 62 430 6 28 F2,15 5 0.514
Basal gangliaTotal neostriatum CPu 348 6 15 254 6 20** 261 6 21** F2,15 5 7.635Neostriatum, dorso-medial rNSdm 310 6 7 254 6 30 249 6 21 F2,15 5 2.503Neostriatum, dorso-lateral rNSdl 290 6 9 241 6 21 227 6 19* F2,15 5 3.634Neostriatum, ventro-medial rNSvm 353 6 5 278 6 26* 262 6 22** F2,15 5 5.911Neostriatum, ventro-lateral rNSvl 335 6 5 290 6 24 272 6 26* F2,15 5 2.417caudal Neostriatum, dorsal cNSd 303 6 12 227 6 24* 245 6 21 F2,15 5 4.161caudal Neostriatum, ventral cNSv 314 6 9 243 6 22* 267 6 27 F2,15 5 3.057Lateral globus pallidus LGP 102 6 10 71 6 9 94 6 18 F2,15 5 1.535Nucleus accumbens Acb 388 6 19 300 6 25** 318 6 16* F2,15 5 5.125
ThalamusAnterior ThalA 268 6 23 208 6 36 219 6 33 F2,15 5 1.051Latero-dorsal ThalLD 333 6 22 269 6 33 296 6 29 F2,15 5 1.300Ventro-lateral ThalVL 241 6 26 187 6 35 226 6 25 F2,15 5 0.918Medio-dorsal ThalMD 288 6 46 230 6 33 259 6 30 F2,15 5 0.607Medio-ventral ThalMV 244 6 34 191 6 31 220 6 27 F2,15 5 0.730
Hypothalamus Hyp 178 6 22 136 6 24 161 6 10 F2,15 5 1.158Brainstem
Superior colliculus CS 261 6 23 249 6 14 187 6 32* F2,15 5 2.264Inferior colliculus CI 231 6 25 207 6 25 172 6 21 F2,15 5 1.619Periaqueductal gray PAG 134 6 20 147 6 26 102 6 13 F2,15 5 1.339
CerebellumCerebellar cortex CBctx 107 6 17 87 6 14 82 6 7 F2,15 5 0.873Molecular layer CBmol 46 6 11 54 6 22 38 6 13 F2,15 5 0.233Granular layer CBgran 226 6 18 188 6 16 128 6 17***,† F2,15 5 8.477Cerebellar deep nuclei CBnuc 61 6 14 55 6 9 73 6 10 F2,15 5 0.657
1Values are the means 6 S.E.M. (n 5 6) in femtomoles per milligram of protein (fm/mg protein). Differences between regions in wild type mice (1/1) versusheterozygote (wv/1) and homozygote (wv/wv) weaver mutants were analyzed by ANOVA followed by a post hoc Fisher F-distribution test.*P , 0.05; **P , 0.01, and ***P , 0.001, between wv/1 or wv/wv mutants and 1/1 mice.†P , 0.05, between wv/1 and wv/wv mutant mice.
218 T.A. READER AND J. SENECAL

highest binding levels (ca. 800 fmol/mg protein) were inLEnt, Piri, and M1o, while lower values were measuredin the other cortices. Also, receptor densities weregreater in M1o than in M1i and greater in S1o than inS1i; however, in the case of Pir cortex [3H]AMPA label-ing was higher in Piri than in Piro.
The highest densities of [3H]AMPA binding in 1/1mice were in the hippocampus (Fig. 2D,G) and thelabeling showed a characteristic pattern, with greaterlabeling in CA1 (F3,20 5 8.080). The intrahippocam-pal topology of AMPA receptors was such that thelowest densities were in the CA3 field; compared to thelatter, levels were higher by 32% in DG (F1,10 5 8.635),by 54% in CA1 (F1,10 5 23.668), and by 33% in CA2(F1,10 5 8.958). There was also intense labeling in Orand in Sub (Table II). In wv/1 mutants, [3H]AMPAbinding was slightly higher in all fields of Ammon’shorn (Fig. 2E,H) and, although these differences werenot significant compared to 1/1 mice, they modifiedthe intrahippocampal receptor distribution. Indeed, la-beling in CA1 was now 146% higher than in DG(F1,10 5 16.669; P , 0.01). The lowest densitieswere again found in CA3; levels in CA1 were 157%greater than in CA3 (F1,10 5 23.119; P , 0.001),and 31% higher in CA2 than in CA3 (F1,10 5 6.972;P , 0.05). In the case of wv/wv mutants, compared to1/1 mice, AMPA receptor densities were higher in theCA1-CA2 fields (Fig. 2F,I) but the only significant dif-ference was in CA2 (130%; P , 0.05). The intrahip-pocampal topology revealed that labeling in CA1 was66% greater than in DG (F1,10 5 11.278; P , 0.01)and CA3 (F1,10 5 11.102; P , 0.01), while levelswere 48% higher in CA2 than in CA3 (F1,10 5 5.965;P , 0.05). In limbic system structures, such as Tu,SPT, and Amyg, measurements of [3H]AMPA binding(600–1000 fmol/mg protein) revealed no significant dif-ferences between the values obtained in weaver mu-tants compared to the levels found in 1/1 mice.
In basal ganglia of 1/1 mice, [3H]AMPA binding washomogenous throughout all divisions of rostral andcaudal neostriatum (F5,30 5 4.852). In Acb, receptorlevels were slightly higher than in CPu, and in LGPlabeling was sparse. There were no differences inAMPA receptor levels between 1/1 mice and both mu-tants, which exhibited uniform labeling in all neostria-tal divisions, i.e., wv/1 F5,30 5 1.116 and wv/wvF5,30 5 2.157.
In thalamus, the highest densities of AMPA recep-tors were in ThalA, followed by ThalLD, ThalMD, andThalMV, and the lowest levels in ThalVL (Fig. 2D). Thesame topology was found in the thalamus of weavermutants, with only slightly lower [3H]AMPA binding inwv/wv (Fig. 2F), in ThalVL (220%), ThalMD (225%),and ThalMV (219%), but these changes were signifi-cant only when compared to wv/1 (Fig. 2E). The label-ing in hypothalamus was discrete and somewhathigher than that observed for NMDA receptors; thus, it
was divided into four regions, designated here as pre-optic antero-medial (HypM) and antero-lateral (HypL),medial (Hyp), and mammilary bodies (MamB). In hy-pothalamus of wv/wv, [3H]AMPA binding was lowerthan in wv/1 mutants, but these differences were re-stricted to HypM (230%) and MamB (232%).
In brainstem of wv/1 mutants, AMPA receptor den-sities were slightly higher in PAG (124%) compared to1/1 mice, while in the wv/wv labeling was lower in CS(226%), CI (225%) and PAG (224%), but only whencompared to wv/1 mutants. In cerebellum of 1/1 mice(Figs. 2J, 4G) an ANOVA analysis confirmed the het-erogenous labeling (F2,15 5 194.945), with densitiesin CBmol more than 2-fold higher than in CBgran(P , 0.001) and 7-fold greater than in CBnuc (P ,0.001), while labeling in CBgran was more than 2-foldhigher than in CBnuc (P , 0.001). This labelingpattern was also present in the wv/1 (F2,15 5 135.766),that had receptor levels in CBmol that were also morethan 2-fold higher than in CBgran (P , 0.001) and7-fold greater than in CBnuc (P , 0.001). Also, la-beling in CBgran was more than 2-fold higher than inCBnuc (P , 0.001). When compared to 1/1 mice,[3H]AMPA binding was higher in CBctx (126%) andCBgran (131%) of these wv/1 mutants (Figs. 2K, 4E).In the wv/wv mutants, compared to 1/1 mice, thelevels of AMPA receptors were 59% higher in CBgran,but this relative increase did not affect the pattern oflabeling (F2,15 5 23.690) still conserved in spite ofprofound cerebellar atrophy (Figs. 2L, 4F). Indeed,[3H]AMPA binding in CBmol was still 2-fold higherthan in CBgran (P , 0.01) and 7-fold greater than inCBnuc (P , 0.001), while labeling in CBgran wasnearly four times higher than in CBnuc (P , 0.05).
KA receptors
The distribution of [3H]kainic acid binding (Fig. 3)was also ubiquitous and heterogenous, and in 1/1 micethere was a 16-fold difference in KA receptor levels(Table III) between the regions of lowest (ThalVL) asopposed to the highest binding densities (Endopiriformnucleus; End). In cerebral cortex of 1/1 mice, the high-est [3H]kainic acid binding levels (ca. 400 fmol/mg pro-tein) were measured in lateral entorhinal (LEnt) andAI cortices, and the lowest densities in Pir (.200fmol/mg protein) with intermediate values in the re-maining cortical regions (Fig. 3A,D,G). In primary mo-tor, primary somatosensory and piriform areas, theinner layers M1i, S1i, and Piri had higher levels ofbinding than the outer M1o, S1o, and Piro, respectively.The cortical topology of KA receptors was overall con-served in the wv/1 mutants (Fig. 3B,E,H) but in thewv/wv KA receptor levels were lower (Fig. 3C,F,I)compared to 1/1 mice, particularly in Cg1 (239%),M1 (226%), M1o (233%), M1i (237%), M2 (237%), AI(227%), S1 (231%), S1o (226%), S1i (232%), S2 (232%),Pir (228%), Piro (225%), Piri (225%), RSG (235%),
IONOTROPIC GLUTAMATE RECEPTORS IN WEAVER MICE 219

Fig. 2

Lent (230%), Vis (230%), and Au (231%). These dif-ferences in the wv/wv were also significant, and ofabout the same magnitude, when compared to thewv/1 mutants.
In hippocampus of 1/1 mice (Fig. 3D,G), intermedi-ate [3H]kainic acid binding densities (ca. 280 fmol/mgprotein) were measured in DG; these levels were 92%higher than in CA1 (F1,10 5 32.776; P , 0.001) and104% greater than in CA2 (F1,10 5 37.316; P ,0.001). In contrast, labeling in CA3 was as intense as inDG, and thus 87% and 99% higher than in CA1(F1,10 5 29.432; P , 0.001) and CA2 (F1,10 533.752; P , 0.001), respectively. Moreover, a well-delimited band of labeling (Fig. 3D) was localized instratum lucidum (SLu) of CA3, which contains themossy fibers derived from the axons of granule cells. Inwv/1 mutants, albeit slight nonsignificant lower label-ing, this pattern of [3H]kainic acid binding was main-tained (Fig. 3E,H) and receptor levels in DG were 79%greater than in CA1 (F1,10 5 9.987; P , 0.01) and75% higher than in CA2 (F1,10 5 9.419; P , 0.05).Also, labeling in CA3 was 69% and 66% higher than inCA1 (F1,10 5 29.432; P , 0.001) and CA2 (F1,10 533.752; P , 0.001), respectively, while the highestbinding was again measured in SLu. In wv/wv mu-tants, [3H]kainic acid binding levels were lower in moststructures of the limbic system and in hippocampus(Fig. 3F,I) differences with 1/1 mice were documentedin DG (223%), CA1 (231%) and CA3 (228%) as well asin SLu (230%). There were no changes in receptordensities in Or and Sub, but lower KA receptor levelswere found in other limbic system structures of wv/wvmutants compared to 1/1 mice, i.e., in Tu (228%), End(229%), and Amyg (231%). Since in the latter struc-tures of wv/1 there were no changes in [3H]kainic acidbinding, the differences in wv/wv were also significantwhen compared to wv/1 mutants, i.e., Tu (230%), SPT(232%), End (222%), and Amyg (226%).
In basal ganglia of 1/1 mice, [3H]kainic acid bindingdensities were uniform throughout the divisions of ros-tral (Fig. 3A) and caudal neostriatum (F5,30 5 1.921),with higher levels in Acb, and very low values in theLGP. There were no differences in labeling between1/1 mice and wv/1 mutants; the latter also exhibiteduniform binding (Fig. 3B) in all neostriatal divisions(F5,30 5 1.882). In wv/wv mutants, compared to 1/1mice, KA receptor densities were lower in CPu (234%),
rNSdm (232%), rNSdl (235%), rNSvm (229%), rNSvl(230%), cNSd (244%), cNSv (231%), and Acb (232%).There were no differences in labeling within neostria-tal divisions (Fig. 3C) of the wv/wv mutants (F5,30 51.370). Differences in the basal ganglia structures ofthe wv/wv were also significant, and of about the samemagnitude, when compared to the wv/1 mutants.
In thalamus of 1/1 mice, KA receptor densities weresparse and the lowest levels were measured in ThalVL(Fig. 3D). This distribution was conserved in the mu-tants, but with lower levels in ThalMD (230%) of thewv/1, as well as in ThalLD (237%), ThalMD (244%),and ThalMV (236%) of the wv/wv (Fig. 3F). The retic-ular thalamic nuclei (Rt) showed a discrete band of[3H]kainic acid binding, and although labeling waslower in the mutants, these differences were not sig-nificant. In hypothalamus of 1/1 mice, [3H]kainic acidbinding was slightly higher than in thalamus and wasof the same magnitude in the wv/1 mutants. In thecase of the wv/wv, KA receptor densities were lower(235%) in both HypM and HypL compared to 1/1mice.
Brainstem KA receptors are sparse (100–180fmol/mg protein) and in wv/1 there were no differencesin densities compared to 1/1 mice. In the wv/wv, re-ceptor levels were lower in CS (231%), CI (241%), andPAG (222%). The substantia nigra (SN) was discretelylabeled by [3H]kainic acid and in wv/wv these bindingsites were also lower (229%). In cerebellar cortex of1/1 mice (Figs. 3J, 4G), KA receptor densities werequite homogenous and amounted to about 200 fmol/mgprotein for total cerebellar cortex, or CBctx. TheANOVA analysis (F2,15 5 16.919) revealed no differ-ences between KA receptor densities in CBmol andCBgran, but 2- and 3-fold higher levels between, re-spectively, CBmol and CBnuc, and between CBgranand CBnuc (P , 0.01). There were no differences in[3H]kainic acid binding in the wv/1 when compared to1/1 mice (Figs. 3K, 4H), and in these mutants theintracerebellar differences (F2,15 5 18.043) were thatlabeling was 3-fold greater in CBmol than in CBnuc aswell as in CBgran compared to CBnuc (P , 0.001). Inthe wv/wv, densities of KA receptors were much lowerin CBctx (237%), CBmol (234%), and CBgran (242%)when compared to 1/1 mice (Figs. 3L, 4I). In spite ofthese lower values, the ANOVA analysis (F2,15 54.861) confirmed that labeling was about 2-fold higherin CBmol than in CBnuc as well as in CBgran com-pared to CBnuc (P , 0.05).
DISCUSSIONThe topology of ionotropic glutamate receptor bind-
ing in brains of wild-type mice (1/1) followed the gen-eral pattern described in rat (Greenamyre et al., 1985;Dure and Young, 1995) and other mice strains(Griesser et al., 1982; Olson et al., 1987; Makowiec etal., 1991; Strazielle et al., 2000). In cerebellum of both
Fig. 2. Autoradiograms of [3H]AMPA binding in the forebrain andcerebellum of a wild-type mouse (1/1; A,D,G,J), and from heterozy-gous (wv/1; B,E,H,K) and homozygous (wv/wv; C,F,I,L) weaver mu-tants. Labeling was slightly higher in S1o, S2, and Pir of wv/1 mu-tants (E) compared to 1/1 mice. In the wv/wv mutants (F), bindingwas also higher in S1o, as well as in CA2. In cerebellum, levels ofAMPA receptors were higher in the CBgran layer of both wv/1 (K)and wv/wv (L) mutants compared to 1/1 mice. The calibration stan-dards for [3H]AMPA binding are in femtomoles per milligram ofprotein (80 –2,441 fmol/mg P), and the horizontal bar correspondsto 1 mm.
IONOTROPIC GLUTAMATE RECEPTORS IN WEAVER MICE 221

wv/1 and wv/wv mutants, there were severe receptoralterations, as expected, since this is the site of theneuropathological hallmark of this mutation; how-ever, there were also differences in distribution inmany other brain regions of weaver mutants whencompared to 1/1 mice and will be discussed, when-ever possible, in relation to the regional expression of
the protein subunits that assemble to form gluta-mate receptors.
Cerebral cortex
In 1/1 mice, moderate NMDA receptor levels weremeasured in all cortical regions; the highest densitieswere in the rostralmost MO and Cg1 areas, but also in
TABLE II. Distribution of [3H]AMPA binding sites in brain regions of wild-type (1/1) and in heterozygote (wv/1)and homozygote (wv/wv) weaver mutant mice1
Region 1/1 wv/1 wv/wv F values
Cortical areasOrbital prefrontal MO 693 6 38 836 6 73 759 6 86 F2,15 5 1.072Anterior cingulate Cg1 704 6 36 837 6 75 691 6 54 F2,15 5 1.963Primary motor M1 678 6 37 803 6 79 713 6 57 F2,15 5 1.128Primary motor, outer M1o 811 6 36 962 6 53 890 6 72 F2,15 5 1.814Primary motor, inner M1i 545 6 10 612 6 32 522 6 44 F2,15 5 2.147Secondary motor M2 721 6 30 824 6 69 755 6 65 F2,15 5 0.831Agranular insular AI 731 6 34 911 6 99 823 6 85 F2,15 5 1.323Primary somatosensory S1 605 6 32 745 6 70 710 6 91 F2,15 5 1.128Primary somatosensory, outer S1o 717 6 28 943 6 66** 876 6 37* F2,15 5 6.199Primary somatosensory, inner S1i 498 6 23 584 6 38 499 6 38 F2,15 5 2.146Secondary somatosensory S2 592 6 36 800 6 64* 726 6 66 F2,15 5 3.432Piriform Pir 736 6 33 885 6 62* 767 6 36 F2,15 5 2.983Piriform, outer Piro 654 6 35 780 6 46 700 6 40 F2,15 5 2.468Piriform, inner Piri 842 6 46 944 6 55 804 6 55 F2,15 5 1.939Retrosplenial granular RSG 489 6 32 513 6 14 461 6 64 F2,15 5 0.378Lateral entorhinal LEnt 884 6 80 965 6 54 738 6 95 F2,15 5 2.170Visual Vis 686 6 47 738 6 29 664 6 79 F2,15 5 0.461Auditory Au 713 6 47 791 6 41 703 6 79 F2,15 5 0.693
HippocampusDentate gyrus DG 1105 6 29 1092 6 61 970 6 132 F2,15 5 0.751Ammon’s horn, field 1 CA1 1282 6 79 1576 6 125 1608 6 182 F2,15 5 1.767Ammon’s horn, field 2 CA2 1110 6 91 1318 6 75 1439 6 105* F2,15 5 3.351Ammon’s horn, field 3 CA3 835 6 39 1006 6 56 975 6 103 F2,15 5 1.636Stratum oriens Or 1166 6 90 1470 6 144 1405 6 168 F2,15 5 1.344Subiculum Sub 837 6 80 832 6 121 828 6 66 F2,15 5 0.002
Limbic systemOlfactory tubercle Tu 649 6 42 746 6 64 634 6 36 F2,15 5 1.543Septum SPT 831 6 67 970 6 75 892 6 41 F2,15 5 1.235Amygdala Amyg 824 6 50 934 6 57 764 6 89 F2,15 5 1.632
Basal gangliaTotal neostriatum CPu 535 6 20 597 6 51 484 6 38 F2,15 5 2.072Neostriatum, dorso-medial rNSdm 540 6 25 585 6 58 481 6 37 F2,15 5 1.535Neostriatum, dorso-lateral rNSdl 465 6 18 537 6 51 440 6 34 F2,15 5 1.813Neostriatum, ventro-medial rNSvm 580 6 29 639 6 57 565 6 30 F2,15 5 0.906Neostriatum, ventro-lateral rNSvl 520 6 22 629 6 57 529 6 37 F2,15 5 2.142caudal Neostriatum, dorsal cNSd 446 6 21 496 6 53 406 6 41 F2,15 5 1.242caudal Neostriatum, ventral cNSv 468 6 25 576 6 28 520 6 58 F2,15 5 1.832Lateral globus pallidus LGP 126 6 5 132 6 15 129 6 12 F2,15 5 0.073Nucleus accumbens Acb 631 6 39 736 6 52 605 6 64 F2,15 5 1.742
ThalamusAnterior ThalA 338 6 28 313 6 44 256 6 28 F2,15 5 1.507Latero-dorsal ThalLD 208 6 18 224 6 28 167 6 30 F2,15 5 1.279Ventro-lateral ThalVL 138 6 9 175 6 18 111 6 14‡ F2,15 5 5.173Medio-dorsal ThalMD 257 6 18 301 6 29 192 6 34† F2,15 5 3.879Medio-ventral ThalMV 235 6 20 295 6 28 190 6 29† F2,15 5 4.079
HypothalamusPreoptic, antero-medial HypM 310 6 10 362 6 41 256 6 24† F2,15 5 3.628Preoptic, antero-lateral HypL 211 6 9 236 6 24 209 6 14 F2,15 5 0.829Medial hypothalamus Hyp 283 6 26 308 6 26 242 6 33 F2,15 5 1.329Mammillary bodies MamB 266 6 33 327 6 20 222 6 25† F2,15 5 3.804
BrainstemSuperior colliculus CS 342 6 22 404 6 23 301 6 24‡ F2,15 5 5.088Inferior colliculus CI 300 6 29 310 6 27 231 6 21† F2,15 5 2.690Periaqueductal gray PAG 194 6 12 241 6 8* 187 6 16‡ F2,15 5 5.866
CerebellumCerebellar cortex CBctx 408 6 18 514 6 23* 491 6 48 F2,15 5 2.918Molecular layer CBmol 652 6 33 816 6 50 738 6 106 F2,15 5 1.346Granular layer CBgran 247 6 13 324 6 17* 392 6 38** F2,15 5 8.302Cerebellar deep nuclei CBnuc 90 6 6 111 6 10 104 6 9 F2,15 5 1.568
1Values are the means 6 SEM (n 5 6) in femtomoles per milligram of protein (fm/mg protein). Differences between regions in wild-type mice (1/1) vs. heterozygote(wv/1) and homozygote (wv/wv) weaver mutants were analyzed by ANOVA, followed by a post-hoc Fisher F-distribution test.*P , 0.05 and **P , 0.001, between wv/1 or wv/wv mutants and 1/1 mice.†P , 0.05 and ‡P , 0.01, between wv/1 and wv/wv mutant mice.
222 T.A. READER AND J. SENECAL

Vis cortex, in line with the distribution of NMDA-sensitive [3H]-L-glutamic acid binding reported for ro-dents (Monaghan and Cotman, 1985) and mice fromthis same strain (Strazielle et al., 2000). Also, labelingwas greater in the outer layers I–III, so that a lamina-tion pattern in outer (M1o, S1o and Piro) vs. inner (M1i,S1i, and Piri) cortical layers was apparent. Indeed, thisneocortical lamination was better visualized here withthe channel blocker [3H]MK-801 than in previous stud-ies with the NMDA antagonist [3H]CPP, or [propyl-1,2-3H]3-[(6)carboxypiperazine-4-yl)propyl-1-phosphonicacid (Monaghan et al., 1988), or by using NMDA-stim-ulated [3H]-L-glutamic acid (Strazielle et al., 2000). Forexample, comparison of relative binding levels in ratparietal cortex revealed in layer IV greater densities of[3H]CPP than NMDA-sensitive [3H]-L-glutamate bind-ing sites, resulting in a more uniform intracortical dis-tribution, and this lead to the proposal that antagonist-preferring sites are relatively abundant in cerebralcortex (Monagham et al., 1988). In the present studywith [3H]MK-801 using equimolar glutamic acid andglycine concentrations, glycine converted antagonist-preferring sites to the agonist conformation sensitive toglutamate, and could thus be stimulated by the phys-iological agonist. Under these conditions [3H]MK-801bound to a greater proportion of sites within the recep-tor channel, independent of the affinities exhibited bythe agonist (glutamic acid/NMDA) and/or the co-ago-nist (glycine) sites of the NMDA receptor complex, thusrevealing the cortical lamination; this has been re-cently reported but not described in detail or quantified(Reader and Senecal, 2001). Another explanation forthis lamination as well as for regional variations inNMDA receptor binding is a differential intracorticalexpression of the protein subunits that assemble toconstitute NMDA receptor channels. Indeed, in situhybridization (ISH) studies of mice neocortex haveshown that NMDA receptors are mainly constituted byNR1, NR2A, and NR2B subunits, with a faint signal forNR2D (Watanabe et al., 1993; Watanabe, 1997); how-ever, there may also be differences between corticalareas. In rat prefrontal cortex, NR2A expression ishighest in all layers, with a signal that is stronger inthe outer layers II and III than in V and VIa (Rudolf etal., 1996). Also, expression of NR2B is highest in layersII and III and less intense and more uniform in theinner layers. In addition to these regional and intra-cortical variations in protein subunit composition, ex-pression of NMDA receptor genes is highly plastic andmay change in response to environmental modifica-tions, as shown in rat visual cortex. Studies by ISHhave indicated that layer IV pyramidal neurons regu-late NR2A expression in relation to the sensory inputsthey receive (Nase et al., 1999); such an increases inthe ratio NR2A/NR2B reduces the mean channel opentime of the NMDA receptor (Wisden et al., 2000) andcould even modify the binding kinetics of [3H]MK-801,
a ligand that labels the interior of the NMDA receptorchannel complex. In weaver mutants, cortical NMDAreceptor levels were lower than in 1/1 mice, and themost affected were the wv/1, but the regional andintracortical topologies were conserved, indicating ei-ther a change in affinities or a lower expression of allNR1, NR2A, and NR2B subunits. In Lc/1 mutantsNMDA-sensitive [3H]glutamic acid binding was loweronly in Cg1, Pir, M1, and M2 areas (Strazielle et al.,2000), suggesting differences in adaptive mechanismsin these two mutations, caused in Lc/1 by the gain offunction of the d2 orphan protein subunit (Zuo et al.,1997).
The cortical distribution of [3H]AMPA binding alsoshows higher labeling in outer layers (Monaghan et al.,1984; Rainbow et al., 1984; Insel et al., 1990) with someregional variations, since AMPA receptors are moreabundant in piriform than in frontal or parietal corti-ces (Dure and Young, 1995). In rat neocortex, ISHstudies have shown expression of mainly GluR1,GluR2, and GluR3 subunits and but sparse GluR4transcripts (Sato et al., 1993). Most projection neuronsin rat neocortex have Ca21-impermeable AMPA recep-tors made of GluR1/GluR2, or GluR2/GluR3, or GluR1/GluR2/GluR3 assemblies, depending on cell type andcortical layer, while GABAergic interneurons haveCa21-permeable AMPA receptors composed of GluR1/GluR4 subunits leading to fast kinetics and high sin-gle-channel conductance (Conti et al., 1994; Wisden etal., 2000). In contrast to NMDA receptors, densities ofcortical AMPA receptors in weaver mutants wereslightly higher in all areas, and these levels were sig-nificantly different in S1o of wv/1 and wv/wv as well asin S2 and Pir of wv/1. In spite of these changes, bothregional and intracortical distributions were main-tained, as was the case for Lc/1 mutants (Strazielle etal., 2000), suggesting a normal topology, albeit with anincrease, in the expression of GluR1-Glu4 subunits.Moreover, since [3H]AMPA binding was unaffected, oreven was higher than in 1/1 mice, a generalized de-velopmental and/or metabolic anomaly can be ruledout, and it can be proposed that AMPA receptors mayincrease to compensate for the loss of NMDA and/or KAreceptors.
In the case of [3H]kainic acid binding, its distributiondiffers from the topology of NMDA and AMPA recep-tors in that the highest densities are in layer I and inthe deeper VI, with the greatest cortical binding infrontal and insular (Monaghan and Cotman, 1982; Un-nerstall and Wamsley, 1983; Miller et al., 1990). Themost abundant KA receptors in rat neocortex are as-semblies of KA2 and GluR7 subunits, particularly inthe deeper VI lamina. However, other transcripts suchas GluR5, GluR6, and KA1 are also expressed, but to alesser degree (Wisden and Seeburg, 1993). In the caseof the GluR7 gene, it is highly expressed in the innercortical layers, probably by pyramidal neurons. Also,
IONOTROPIC GLUTAMATE RECEPTORS IN WEAVER MICE 223

Fig. 3
224 T.A. READER AND J. SENECAL

there are regional variations in GluR7 expression inrodent neocortex, and a thin but intense sublayer hasbeen described in lamina III of rat (Lomeli et al., 1992;Wisden and Seeburg, 1993) and mice (see fig. 1 inStrazielle et al., 2000). The expression of GluR5 isscattered to a few neurons, perhaps of GABAergic na-ture, and its distribution varies throughout corticalregions and ontogeny, with a certain predominance inouter layers of sensory cortices (Bahn et al., 1994).Binding densities of [3H]kainic acid were lower than in1/1 mice in all cortical areas of wv/wv mutants, butwith no differences in wv/1 mutants; the decreaseswere more generalized than in Lc/1 mutants (Stra-zielle et al., 2000) and with conservation of the regionaland intracortical distributions, indicating decreased af-finities for [3H]kainic acid binding or, alternatively,reduced expression of GluR7 and KA2 transcripts, andwith a conservation of their proportions, thus preserv-ing the higher labeling measured in inner cortical lay-ers. The lack of changes in wv/1 suggests that a certainthreshold has to be attained before KA receptors aremodified, whether in their binding properties or in therelative proportion of their protein subunits.
Hippocampus and limbic structures
The hippocampus presents high numbers of gluta-mate receptors (Halpain et al., 1984; Greenamyre etal., 1985; Dure and Young, 1995), and binding densitiesin this formation were the highest for NMDA andAMPA receptors, while considerable levels of KA siteswere also documented. The NMDA receptor complex ismainly postsynaptic in the perforant pathway to DGgranule cells (Bekenstein et al., 1990) and is present aswell in the Schaffer collaterals to CA1 (Collingridge etal., 1983). The protein subunits that assemble to formNMDA receptors in mouse pyramidal cells of CA1–CA3fields and in DG granule cells are NR1, NR2A, andNR2B (Watanabe, 1997), although there is some ex-
pression of the NR2C gene in small cells, probably ofglial nature (Wisden et al., 2000). Thus, NMDA recep-tors in pyramidal neurons and granule cells of the DGare made most likely of NR1/NR2A, NR1/NR2B, and/orNR1/NR2A/NR2B subunits. Hippocampal GABAergicinterneurons express NR1, NR2A, and NR2B subunits,but in CA1 and CA3 stratum oriens NR2D is alsoexpressed (Standaert et al., 1996). The NR1/NR2D as-semblies activate and deactivate slowly and are lesssensitive to voltage-dependent Mg21 block than themore usual NR1/NR2A of NR1/NR2B assemblies, andwhen glutamate binds they may initiate action poten-tials even when the neuron is not greatly depolarized.As was the case for cortical areas, the lower [3H]MK-801 binding in hippocampus of weaver mutants couldbe explained by overall change in affinities, or a lowerexpression of all NR1, NR2A, and NR2B protein sub-units. Also, as in cortex, the differences were greater inDG, CA1, and CA3 of wv/1 than in wv/wv mutants, butnot significantly different in Or, suggesting a normalexpression of NR2D transcripts. In Lc/1 mutants,NMDA-sensitive [3H]glutamic acid binding densitieswere lower than in 1/1 mice only in the CA2–CA3fields, reflecting a more selective and better labeling ofNMDA-receptor channel by [3H]MK-801 binding or,alternatively, differences in the adaptive mechanismsof these two different gain-of-function mutations. Spa-tial memory and learning in rodents is related to hip-pocampal function (Morris et al., 1982; Whishaw, 1998;Wood et al., 1999) and plasticity (Cotman et al., 1988)and cognitive development has implicated glutamateNMDA receptors within this structure (Liu et al.,2000). The NMDA receptor has been proposed as asynaptic coincidence detector (Bliss and Collingridge,1993) and mice that overexpress NR2B subunits havesuperior ability in learning and memory, showing thatthey are critical to gate the age-dependent thresholdfor plasticity and memory formation (Tang et al., 1999).The lower NMDA receptor levels found in weaver mu-tants may well explain their navigational deficits in theMorris water maze task (Lalonde and Botez, 1986).
In hippocampus, AMPA receptors are present at highlevels in stratum moleculare of DG, in stratum radia-tum and Or of CA1, and in stratum radiatum of CA3(Rainbow et al., 1984; Baudry et al., 1990). In rathippocampus, AMPA receptors are essentially com-posed of GluR1–GluR4 subunits (Sato et al., 1993; Pe-tralia et al., 2000). The GluR1, GluR2, and GluR3 arestrongly expressed by all pyramidal neurons and DGgranule cells (Keinanen et al., 1990), while nonpyrami-dal cells express mainly GluR1 with little GluR2. Thereis also some expression of GluR4, confined to CA1 py-ramidal neurons and DG granule cells. Interneurons inthe hippocampus are labeled with GluR1 and GluR4probes and some cells express GluR3 (Monyer et al.,1991). In most GABAergic interneurons there is alsoGluR2 expression, but at lower levels than in pyrami-
Fig. 3. Autoradiograms of [3H]kainic acid binding in the forebrainand cerebellum of a wild-type mouse (1/1; A,D,G,J), and from het-erozygous (wv/1; B,E,H,K) and homozygous (wv/wv; C,F,I,L) weavermutants. In the wv/wv mutants, KA receptor levels were lower in allcortical areas, notably in Cg1, M1, M2, S1, S2, AI, and Pir (C). Thislevel of section also illustrates lower [3H]kainic acid labeling in theolfactory tubercle (Tu), and in all four quadrants of rostral neostria-tum. At a more posterior plane of section of a 1/1 mouse (D), note theintense labeling in stratum lucidum (SLu) and in the endopirifomnucleus (End) as well as the distinct reticular thalamic nuclei (Rt). Inwv/wv mutants (F), KA receptors are lower in RSG, S1, S2, End, Lent,Piro, and Piri cortices, compared to both 1/1 (D) and wv/1 mutants(E). Also, note in F that labeling was lower in ThalLD, ThalMD,ThalMV, DG, CA1, and CA3, including SLu. Lower densities of KAreceptors were also measured in Vis and Au cortices (I), compared toboth 1/1 (G) and wv/1 mutants (H). Labeling in cerebellum of 1/1mice was uniform in CBctx, albeit slightly higher but not significantlydifferent, in CBgran than in CBmol (J). There were no differences inthe wv/1 mutants (K), but in the wv/wv mutants [3H]kainic acidbinding was much lower (L). Note also decreased labeling in inferiorcolliculus (CI). The calibration standards for [3H]kainic acid bindingare in femtomoles per milligram of protein (39–2,056 fmol/mg P), andthe horizontal bar corresponds to 1 mm.
IONOTROPIC GLUTAMATE RECEPTORS IN WEAVER MICE 225

dal neurons or DG granule cells (Racca et al., 1996).The higher relative expression of GluR1/GluR4, but notGluR2 or GluR3 (Catania et al., 1995), suggests thathippocampal GABAergic interneurons, as was the casefor interneurons in cerebral cortex, have highly Ca21-
permeable AMPA receptors with fast kinetics and highsingle-channel conductance (Conti et al., 1994; Wisdenet al., 2000). In weaver mutants, compared to 1/1mice, there were no differences in [3H]AMPA labelingexcept for a slight increase in the CA2 field, thus indi-
TABLE III. Distribution of [3H]kainic acid binding sites in brain regions of wild-type (1/1) and in heterozygote (wv/1)and homozygote (wv/wv) weaver mutant mice1
Region 1/1 wv/1 wv/wv F values
Cortical areasOrbital prefrontal MO 352 6 30 367 6 37 260 6 30† F2,15 5 3.163Anterior cingulate Cg1 387 6 22 365 6 36 238 6 68**,‡ F2,15 5 7.620Primary motor M1 335 6 16 286 6 24 213 6 25**,† F2,15 5 7.587Primary motor, outer M1o 217 6 7 195 6 18 143 6 15**,† F2,15 5 6.873Primary motor, inner M1i 377 6 17 327 6 32 239 6 25***,† F2,15 5 7.708Secondary motor M2 378 6 17 328 6 33 238 6 22***,† F2,15 5 8.038Agranular insular AI 416 6 16 415 6 42 304 6 29*,† F2,15 5 4.339Primary somatosensory S1 281 6 13 259 6 22 194 6 21**,† F2,15 5 5.794Primary somatosensory, outer S1o 165 6 11 158 6 15 122 6 13* F2,15 5 3.080Primary somatosensory, inner S1i 359 6 15 329 6 26 246 6 24**,† F2,15 5 7.033Secondary somatosensory S2 282 6 9 266 6 27 193 6 23**,† F2,15 5 5.065Piriform Pir 184 6 14 181 6 18 133 6 14*,† F2,15 5 3.256Piriform, outer Piro 149 6 10 137 6 12 112 6 10* F2,15 5 3.009Piriform, inner Piri 197 6 9 192 6 19 147 6 17* F2,15 5 3.038Retrosplenial granular RSG 215 6 17 170 6 23 139 6 13** F2,15 5 4.527Lateral entorhinal LEnt 427 6 25 396 6 36 299 6 29**,† F2,15 5 4.873Visual Vis 297 6 24 296 6 30 208 6 23*,† F2,15 5 3.846Auditory Au 325 6 21 305 6 27 223 6 29*,† F2,15 5 4.425
HippocampusDentate gyrus DG 280 6 22 235 6 26 188 6 17** F2,15 5 4.355Ammon’s horn, field 1 CA1 146 6 12 131 6 19 101 6 7* F2,15 5 2.870Ammon’s horn, field 2 CA2 137 6 12 134 6 20 110 6 12 F2,15 5 0.931Ammon’s horn, field 3 CA3 273 6 18 222 6 27 196 6 17* F2,15 5 3.461Stratum lucidum SLu 704 6 66 631 6 59 496 6 41* F2,15 5 3.537Stratum oriens Or 133 6 9 128 6 17 98 6 7 F2,15 5 2.623Subiculum Sub 188 6 18 206 6 16 150 6 21 F2,15 5 2.302
Limbic systemOlfactory tubercle Tu 434 6 30 451 6 34 314 6 30*,‡ F2,15 5 5.690Septum SPT 299 6 15 318 6 33 215 6 32† F2,15 5 3.798Endopiriform nucleus End 737 6 36 672 6 52 524 6 52**,† F2,15 5 5.275Amygdala Amyg 348 6 16 321 6 32 239 6 24**,† F2,15 5 5.189
Basal gangliaTotal Neostriatum CPu 397 6 15 368 6 31 262 6 28**,‡ F2,15 5 7.675Neostriatum, dorso-medial rNSdm 368 6 17 364 6 28 249 6 27**,‡ F2,15 5 7.621Neostriatum, dorso-lateral rNSdl 351 6 28 352 6 29 230 6 25**,‡ F2,15 5 6.579Neostriatum, ventro-medial rNSvm 350 6 31 361 6 29 250 6 32*,† F2,15 5 3.931Neostriatum, ventro-lateral rNSvl 431 6 31 433 6 40 303 6 36*,† F2,15 5 4.176caudal Neostriatum, dorsal cNSd 407 6 44 323 6 19 227 6 25***,† F2,15 5 8.442caudal Neostriatum, ventral cNSv 451 6 29 407 6 25 311 6 39**,† F2,15 5 5.115Lateral globus pallidus LGP 77 6 9 85 6 11 76 6 15 F2,15 5 0.156Nucleus accumbens Acb 554 6 43 523 6 40 377 6 51*,† F2,15 5 4.381
ThalamusAnterior ThalA 165 6 23 135 6 11 111 6 18 F2,15 5 2.208Latero-dorsal ThalLD 86 6 13 60 6 7 54 6 5* F2,15 5 3.562Ventro-lateral ThalVL 47 6 14 37 6 9 35 6 8 F2,15 5 0.329Medio-dorsal ThalMD 179 6 19 126 6 10* 101 6 8*** F2,15 5 8.822Medio-ventral ThalMV 121 6 11 106 6 10 78 6 8** F2,15 5 4.965Reticular thalamic nuclei Rt 177 6 23 151 6 12 121 6 22 F2,15 5 2.042
HypothalamusPreoptic, antero-medial HypM 286 6 18 283 6 24 186 6 25**,‡ F2,15 5 6.161Preoptic, antero-lateral HypL 221 6 22 189 6 18 143 6 24* F2,15 5 3.314Medial hypothalamus Hyp 201 6 14 178 6 25 143 6 17 F2,15 5 2.236Mammillary bodies MamB 189 6 26 191 6 19 140 6 19 F2,15 5 1.830
BrainstemSubstantia nigra SN 105 6 11 132 6 10 75 6 11‡ F2,15 5 6.988Superior colliculus CS 177 6 16 193 6 19 123 6 15*,† F2,15 5 4.602Inferior colliculus CI 145 6 24 135 6 18 86 6 9* F2,15 5 3.036Periaqueductal gray PAG 147 6 10 164 6 17 114 6 18† F2,15 5 2.792
CerebellumCerebellar cortex CBctx 202 6 23 202 6 18 128 6 21*,† F2,15 5 4.095Molecular layer CBmol 205 6 19 215 6 24 135 6 22*,† F2,15 5 3.946Granular layer CBgran 226 6 26 221 6 25 131 6 19*,† F2,15 5 5.084Cerebellar deep nuclei CBnuc 78 6 10 66 6 9 66 6 9 F2,15 5 0.520
1Values are the means 6 SEM (n 5 6) in femtomoles per milligram of protein (fm/mg protein). Differences between regions in wild-type mice (1/1) vs. heterozygote(wv/1) and homozygote (wv/wv) weaver mutants were analyzed by ANOVA, followed by a post-hoc Fisher F-distribution test.*P , 0.05, **P , 0.01, and ***P , 0.001, between wv/1 or wv/wv mutants and 1/1 mice.†P , 0.05 and ‡P , 0.01, between wv/1 and wv/wv mutant mice.
226 T.A. READER AND J. SENECAL

cating a seemingly normal expression of all GluR1–GluR4 transcripts. In Lc/1 mutants, however, levels of[3H]AMPA binding were higher in CA1 as well as inCA2–CA3 fields (Strazielle et al., 2000), but the overallhippocampal distribution of AMPA receptors was con-served in both mutations.
Densities of KA receptors in hippocampus are some-what lower, except in SLu of CA1 and in stratum mo-leculare of DG (Monaghan and Cotman, 1982; Unner-stall and Wamsley, 1983). The KA receptor subunitsexpressed in rat hippocampus within the CA1 field are
basically made from KA2 subunits, although there issome expression of GluR5, GluR6, GluR7, and KA1transcripts. In contrast, in CA3 they are made up ofKA1/KA2 subunits with some lower expression ofGluR6 and GluR7 transcripts (Wisden and Seeburg,1993), and in DG there are high levels of KA1 and KA2expression with moderate amounts of GluR6 andGluR7. Thus, the possible KA receptor configurationsin CA1 pyramidals are GluR6/KA2, while in CA3 py-ramidals their composition could be GluR6/KA2, orGluR6/KA1, or GluR6/KA1/KA2 and in DG granule
Fig. 4. Comparison of the labeling with [3H]MK-801 (A,B,C),[3H]AMPA (D,E,F), and [3H]kainic acid (G,H,I) in sections throughthe brainstem and cerebellum that include the deep cerebellar nuclei(CBnuc). The histological images shown in all right panels are fromthe same sections that were used for the ligand binding incubations,
postfixed and stained (Nissl) with cresyl-violet. The calibration scalesfor each ligand are in femtomoles per milligram of protein: 137–2,721fmol/mg P for [3H]MK-801, 80–2,441 fmol/mg P for [3H]AMPA, and39–2,056 fmol/mg P for [3H]kainic acid. The horizontal bar corre-sponds to 1 mm.
IONOTROPIC GLUTAMATE RECEPTORS IN WEAVER MICE 227

cells the receptors derive from KA1, KA2, GluR6, andGluR7 (Wisden et al., 2000). In addition, in Sub, in Orof CA1, and in DG a punctate pattern of GluR5 expres-sion has been attributed to oriens-alveus-lacunosum-moleculare interneurons, while a few GluR-5-positivecells in stratum radiatum and in the pyramidal celllayer may be GABAergic in nature. Most GABAergicinterneurons of the pyramidal layer also expressGluR6 and there are but few GluR7 mRNA-positivecells in mouse Or and radiatum (Bureau et al., 1999).In wv/wv, but not in wv/1 mutants, as well as in Lc/1mice (Strazielle et al., 2000), [3H]kainic labeling waslower than in 1/1 mice for all hippocampal divisions,suggesting altered binding properties and/or expres-sion of KA1, KA2, GluR6, and GluR7 subunits. Thedecreased labeling in SLu, attributed to presynapticreceptors on the mossy fibers of granule cells, couldalso be explained by changes in KA1, KA2, GluR6, andGluR7 protein subunits.
Basal ganglia and related structures
In rodent basal ganglia, the highest amounts ofNMDA-sensitive glutamate receptors are found in CPuand Acb, with very low levels in LGP, subthalamicnucleus, substantia nigra, or ventral tegmental area(Sakurai et al., 1991; Albin et al., 1992; Dure andYoung, 1995; Strazielle et al., 2000). It has been shownby ISH that mice express the NR1, NR2A, and NR2Bsubunits (Watanabe, 1997), while rats express in addi-tion low levels of NR2C in some neurons and glial cells(Standaert et al., 1999). Also, cholinergic interneuronsappear to express NR1/NR2B/NR2D transcripts (Stan-daert et al., 1996, 1999). In LGP, there is some expres-sion of NR1, NR2A, NR2B, and NR2D (Watanabe,1997; Petralia et al., 2000). Compared to 1/1 mice,[3H]MK-801 labeling was lower in rNSvm, cNSd, andcNSv of wv/1 mutants and in rNSdl, rNSvm, andrNSvl of wv/wv, but such differences were not found byNMDA-stimulated [3H]glutamic acid binding in Lc/1mutants (Strazielle et al., 2000), perhaps a reflection ofthe depletions of DA (Lane et al., 1977; Schmidt et al.,1982; Roffler-Tarlov and Graybiel, 1984) and the 5-HThyperinnervations (Stotz et al., 1993; Dewar, 1999;Reader et al., 2001) found in weaver but not in Lc/1mice. In the Acb, levels of [3H]MK-801 binding werelower in wv/1 and wv/wv mutants than in 1/1; thisstructure also shows a profound dopaminergic impair-ment (Reader et al., 1999). Thus, this suggests that insome CNS regions there could be a compensatorydownregulation of NMDA receptors in response to adopaminergic deficiency, but whether these changesare due to interactions between neurotransmitters or aconsequence of increased depolarization caused by thegain-of-function of mutated wvGIRK2 channels (Lesterand Karshin, 2000), or to a direct effect of the weavergene on the genomic expression of NMDA receptorprotein subunits still remains to be examined.
In the case of [3H]AMPA binding, the highest densi-ties are in Acb and CPu (Albin et al., 1992), and al-though relative levels of AMPA sites in LGP are some-what higher than NMDA receptors, the latter structurewas very sparsely labeled (Strazielle et al., 2000). Ithas been shown by ISH that medium spiny neurons inCPu express GluR1 and GluR2 transcripts (Sato et al.,1993), while GluR3 is localized to medium spiny neu-rons coexpressing enkephalin and substance P, andGluR4 is expressed by cholinergic interneurons (Ste-fani et al., 1998). In LGP, the main transcripts areGluR1 and GluR4, with lower levels of expression forGluR2 and GluR3 (Sato et al., 1993; Petralia et al.,2000). In contrast to Lc/1 mutants, which show slightincreases in [3H]AMPA binding (Strazielle et al., 2000),there were no differences in labeling of neostriatum,LGP, and Acb of wv/1 and wv/wv, indicating a conser-vation of all GluR1–Glu4 transcripts.
Finally, KA receptors in the basal ganglia also aremoderate to sparse, with relatively higher [3H]kainicacid labeling in Acb followed by CPu and much lowerKA receptor densities in LGP (Albin et al., 1992; Dureand Young, 1995; Strazielle et al., 2000). In rat CPu,the highest expression is for KA2 followed by GluR6 inall medium-sized neurons and cholinergic neurons, andmuch lower expression of GluR5, GluR7, and KA1,while in LGP there are very low levels of GluR5,GluR7, KA1, and KA2 transcripts (Wisden and See-burg, 1993). As in cerebral cortex and hippocampus,[3H]kainic acid binding densities were lower in wv/wvthan in 1/1 mice, but there were no differences inwv/1 mutants; this was documented in neostriatumand Acb, indicating that affinities for [3H]kainic aciddiminished or, alternatively, that expression of GluR7and KA2 subunits was downregulated to compensatefor the dopaminergic deficit, or in response to excessivedepolarization, similar to that documented for cerebel-lar granule cells (Kofuji et al., 1996; Tong et al., 1996)of this gain-of-function mutation expressing anomalouswvGIRK2 channels. Similar lower [3H]kainic acid bind-ing was found in Lc/1 mutants (Strazielle et al., 2000),and since in this mutation there are no apparent DAdeficits but a d2 gain-of-function that produces consti-tutively-active glutamate receptors (Zuo et al., 1998;Lester and Karshin, 2000), it can be now proposed thatan excessive and sustained basal depolarization thatreaches a certain threshold can lead to a compensatorydecrease in the expression of KA receptors.
Thalamus and hypothalamus
Only moderate to low levels of glutamate receptorshave been described in thalamus of rodents, and evenlower amounts in hypothalamus (Monaghan and Cot-man, 1985; Albin et al., 1992; Strazielle et al., 2000). Inmedial thalamus of rat, the binding properties of asubset of NMDA receptors may differ from the classicalagonist- and antagonist-preferring forms (Beaton et
228 T.A. READER AND J. SENECAL

al., 1992), while binding studies with MK801 have ledto the proposal that these receptors may be regulateddifferently (Sakurai et al., 1993). The distribution ofNMDA receptor subunits varies in the different tha-lamic nuclei, and in the mouse the main transcripts areNR1, NR2A, NR2B, with much lower levels of NR2Cand NR2D (Watanabe, 1997; Petralia et al., 2000).Other ionotropic glutamate receptors are moderate tosparse (Monagham et al., 1984; Baudry et al., 1990),except for intense labeling in the Rt (Miller et al., 1990;and see fig. 1C in Strazielle et al., 2000). Many tha-lamic nuclei express mainly GluR1, GluR2, and GluR4,with a predominance of GluR3 and GluR4 in ventraland lateral groups, and a prevalence of GluR4 in Rt(Sato et al., 1993; Petralia et al., 2000). In the case ofKA receptors, GluR7 is highly expressed in Rt togetherwith lesser amounts of GluR6, KA1, and KA2, GluR5 isvery apparent in anteroventral and midline nuclei withsome expression of GluR7 and KA2 (Wisden and See-burg, 1993; Petralia et al., 2000).
Brainstem and cerebellum
Glutamate receptors in brainstem are sparse andwere quantified in a few identified regions. For NMDAreceptors, levels of [3H]MK-801 binding were lower inCS of weaver mutants, but this difference was signifi-cant only in the wv/wv as if the loss of sites was depen-dent on the homozygous phenotype. AMPA receptordensities in PAG were higher in wv/1 mutants than in1/1 mice, but levels in the wv/wv were maintained. ForKA receptors, there were differences in the wv/wv com-pared to 1/1 mice, with lower levels in CS and CI. TheSN was well-labeled with [3H]kainic acid in both parscompacta and pars reticulata, and there was a trend tohigher levels in wv/1 and somewhat lower in wv/wv, areflection of the changes in dendritic fields of dopami-nergic neurons and loss of DA neurons, respectively.
It is now well established that in cerebellum gluta-mate is the neurotransmitter in synapses of mossyfibers onto granule cells, climbing fibers onto Purkinjeneurons, parallel fibers onto Purkinje neurons, andstellate/basket cells, as well as between mossy andclimbing fibers with Golgi interneurons (Oertel, 1993;Voogd et al., 1996). NMDA receptor levels are moder-ate in the CBgran, with lower densities in CBmol layer;they have been located on granule cells as well as onstellate or basket cells (Monaghan and Cotman, 1985;Olson et al., 1987; Strazielle et al., 2000). Purkinjeneurons of adult mice express only the NR1 gene (Wa-tanabe et al., 1994; Watanabe, 1997) but there seems tobe some variability for mouse lines, including mutantsand the mismatch between NR1 and NR2 transcriptsleading to no responses or to low-conductance NMDAreceptors, which has eluded explanation (Cull-Candyet al., 1998), but NR1 subunits could associate with d2orphan subunit to form a glutamate receptor channel(Wisden et al., 2000). Adult granule cells express the
highest NR2C gene transcripts in brain (Watanabe etal., 1994). Interestingly, during development there arechanges in expression, so that young pre- and postmi-gratory granule cells express NR1 and NR2B tran-scripts, while in the second postnatal week NR2A andNR2C genes are turned on with a decline in mRNAlevels for NR2B (Monyer et al., 1994). The stellate andbasket cells express NR1 and NR2D genes (Watanabeet al., 1994), as observed for other GABAergic inter-neurons, while Golgi cells have been proposed to ex-press also the NR1 and NR2D transcripts (Cull-Candyet al., 1998). There is also some expression of mRNAsfor NR2B and NR1 genes in Bergmann glial cells(Luque and Richards, 1995). In wv/wv mutants, NMDAreceptor levels were considerably lower in CBgran,compared to the wv/1 and to 1/1 mice; this was ex-pected since glutamate is the neurotransmitter in syn-apses between mossy fibers and granule cells; the lat-ter are lost in this mutant (Rakic and Sidman,1973a,b,c; Sotelo and Changeux, 1974; Smeyne andGoldowitz, 1989) as was the case for the lower labelingwith NMDA stimulate-[3H]glutamic acid observed inthe Lc/1 (Strazielle et al., 2000). The residual [3H]MK-801 binding in weaver mutants can be attributed toremaining granule and other cells, i.e., Golgi interneu-rons and Bergmann glia that also express NMDA re-ceptors.
AMPA receptors in the Cbmol are presumably ondendrites of Purkinje neurons (Olson et al., 1987; Ma-kowiec et al., 1991; Strazielle et al., 2000). Purkinjecells express mainly GluR2 and GluR3 transcripts, butthere is also some GluR1 and GluR4 (Sato et al., 1993);multiple types of AMPA receptors on Purkinje neuronscould be differentially located at parallel and climbingfiber synapses. Granule cells contain only GluR2 andsome GluR4, the GABAergic interneurons of the stel-late and basket cell types express GluR2 and GluR3,and Golgi cells possibly GluR3 (Keinanen et al., 1990;Sato et al., 1993). Bergmann glial cells contain GluR1and GluR4 mRNAs (Keinanen et al., 1990; Monyer etal., 1991; Sato et al., 1993) and since the GluR2 gene isnot expressed, their AMPA receptors are Ca21-perme-able with fast kinetics and high single-channel conduc-tance (Geiger et al., 1995). In the CBmol layer, AMPAreceptor levels were higher in weaver mutants than in1/1 mice; these differences were significant only forthe wv/1 and there was but a trend in the wv/wv. Thealtered migration of Purkinje cells in weaver mutantscauses changes in dendritic trees, but these neuronsmay still express AMPA receptors. However, theirmain presynaptic input, i.e., parallel fibers, are de-creased because of the profound loss of granule cells,and this lack of glutamatergic neurotransmission maylead to the upregulation of AMPA postsynaptic recep-tors located on Purkinje cell dendrites, particularly inthe wv/1 that shows much higher levels of [3H]AMPAbinding than the wv/wv. Although there is some Pur-
IONOTROPIC GLUTAMATE RECEPTORS IN WEAVER MICE 229

kinje cell loss in weaver mutants (Smeyne and Goldo-witz, 1990), it is less profound than in Lc/1 mice.Indeed, in Lc/1 mutants AMPA receptor levels werelower in both CBmol (Strazielle et al., 2000) because ofthe massive loss of Purkinje cells and their dendriteswhere these receptor have a postsynaptic location, andin CBgran due to the very heavy loss of granule cells inthis mutant compared to weaver mice.
The distribution of KA binding sites has been ini-tially reported to be similar to NMDA-sensitive bind-ing, i.e., higher labeling in CBgran, presumably locatedon granule cells (Olson et al., 1987); however, recentstudies with [3H]kainic acid in 1/1 mice have revealedhigher labeling in CBmol than in CBgran, and thistopology was conserved in the Lc/1 mutant, whichsuffers considerable degeneration of granule cells(Strazielle et al., 2000), concordant with the earlierproposal that KA receptors may also be present onpresynaptic parallel fibers (Henke et al., 1981). It isnow established that Purkinje neurons express GluR5and KA1 subunits (Bettler et al., 1990; Wisden andSeeburg, 1993) and may assemble their receptor asKA1/GluR5. In rodent Purkinje cells there are alsohigh levels of the d2 orphan subunit mRNA, specificallyat synapses between their dendritic spines and parallelfibers (Zuo et al., 1997) and this synaptic organizationcontributes to the phenomenon of long-term depression(LTD). The d2 subunit may assemble with the NR1subunit, or even with AMPA or kainate. In knockoutmice, lack of the d2 subunit causes locomotor ataxia,altered Purkinje cell synapse formation, and suppres-sion of LTD (Hirano et al., 1995; Kashiwabuchi et al.,1995), while in the mutation Lc/1 gain-of-function ofthe d2 leads to excitotoxic death of Purkinje cells (Zuoet al., 1997). Granule cells assemble KA2/GluR6 recep-tors (Wisden and Seeburg, 1993), and based on KA2immunoreactivity on parallel fibers it has been sug-gested that some of the granule cell GluR6/KA2 recep-tors may have a presynaptic function (Petralia et al.,1994). The GABAergic interneurons, i.e., basket andstellate cells, only express moderate amounts of GluR7transcripts (Lomeli et al., 1992; Wisden and Seeburg,1993) and such homomeric receptor channels have alow sensitivity towards glutamate (Schiffer et al.,1997). In the wv/wv there were much lower levels of[3H]kainic acid binding than in 1/1 mice in both theCBgran and CBmol layers and this can be imputed tothe loss of granule cells. Indeed, decreases of differingseverity have been reported in murine mutants withvariable losses of cerebellar granule cells, such asweaver, staggerer, and reeler (Grieser et al., 1982) aswell as in Lc/1 (Strazielle et al., 2000), in line with theproposal that KA receptors are located on presynapticparallel fibers (Henke et al., 1981). In addition, KA2and GluR6 protein subunits, which form high-affinityKA receptors, are expressed in granule cells (Wisdenand Seeburg, 1993), while the existence of GluR7 tran-
scripts in basket and stellate interneurons could ac-count for the residual binding in both layers.
The deep cerebellar nuclei receive inhibitory affer-ents from Purkinje cells and excitatory afferents fromclimbing and mossy fibers (Oertel, 1993). Decreases inglutamate concentrations observed in these nuclei ofweaver and staggerer mice but not in the nervous mu-tant (Roffler-Tarlov and Sidman, 1978; Roffler-Tarlovand Turey, 1982) have been explained by the loss ofgranule cells, perhaps causing mossy fiber hypofunc-tion. The main NMDA protein subunit transcripts ex-pressed in the deep cerebellar nuclei are NR1, NR2A,and NR2D (Watanabe et al., 1994), as well as GluR2,GluR3, and GluR4 transcripts (Sato et al., 1993).Although binding of [3H]MK-801, [3H]AMPA and[3H]kainic acid was very sparse in 1/1 mice andweaver mutants, its levels were still detectable abovebackground and there were no differences in labeling.In contrast, NMDA receptor binding was increased inthe deep nuclei of Lc/1 mutants, which have a moder-ate cell loss of approximately 30% (Heckroth, 1994),but in this mutant there is an additional loss of allPurkinje cell axons leading to a structural atrophy ofabout 45%. When the labeling was corrected by surfacearea it was similar in Lc/1 and 1/1 mice, indicating,as is the case for weaver mutants studied here, a rela-tive preservation of NMDA and AMPA receptors, lo-cated on the spared cells of the deep nuclei; however,lower levels of KA receptors in Lc/1 mutants wereattributed (Strazielle et al., 2000) to the loss of Pur-kinje cell axons and/or to degeneration of intrinsic cellscontaining this receptor subtype.
Concluding remarks
The lower levels of NMDA receptor-channels in cer-ebellar granular layer of weaver mutants can be attrib-uted to the postnatal loss of granule cells and Purkinjeneurons, as was the case for the Lc/1 mutant. How-ever, decreased [3H]MK-801 labeling in other brainregions, notably in neocortical areas associated withmotor control and somatosensory perception, as well asin neostriatum and nucleus accumbens, may representcompensatory adaptations to the increased overall neu-ronal excitability caused in the wvGIRK2 gain-of-func-tion mutation, or by an altered developmental expres-sion of glutamate receptor subunits. In hippocampus,the relative lack of NMDA receptors will have specificconsequences for cognitive processes, particularly inplasticity, spatial memory, and learning. AlthoughAMPA receptor levels were relatively well conserved,there were generalized losses of KA receptors that af-fected cerebellar but also extracerebellar structures,particularly in cortical areas, hippocampus, limbic sys-tem, basal ganglia, and diencephalon. Thus, the weaverphenotype is not only the outcome of an agranularcerebellum combined with a DA deficit but, in addition,presents severe alterations in glutamate receptor to-
230 T.A. READER AND J. SENECAL

pology. These changes may subtend extensive modifi-cations in the delicate balance between ionotropicglutamate receptor subtypes directly affecting gluta-matergic transmission, and could thus explain the va-riety of behavioral and cognitive deficits observed inthis mutant.
ACKNOWLEDGMENT
The authors thank Mrs. Daniel Cyr and Claude Gau-thier for technical assistance with the illustrations.
REFERENCES
Albin RL, Makowiec RL, Hollingsworth ZR, Dure LS IV, Penney JB,Young AB. 1992. Excitatory amino acid binding sites in the basalganglia of the rat: a quantitative autoradiographic study. Neuro-science 46:35–48.
Bahn S, Volk B, Wisden W. 1994. Kainate receptor gene expression inthe developing rat brain. J Neurosci 14:5525–5547.
Barlow RB. 1983. Biodata handling with microcomputers. Amster-dam: Elsevier Science Publishers.
Baudry M, Monaghan D, Cotman C, Altar CA. 1990. Regional distri-bution of a-[3H]amino-3-hydroxy-5-methylisoxazole-4-propionic acidbinding sites in the rat brain: effect of chemical modification ofSH-groups in tissue sections. J Neurochem 54:1682–1688.
Bayer SA, Wills KV, Triarhou LC, Verina T, Thomas JD, Ghetti B.1995. Selective vulnerability of late-generated dopaminergic neu-rons of the substantia nigra in weaver mutant mice. Proc Natl AcadSci USA 92:9137–9140.
Beaton JA, Stemsrud K, Monaghan DT. 1992. Identification of a novelN-methyl-D-aspartate receptor population in the rat medial thala-mus. J Neurochem 59:754–757.
Bekenstein JW, Bennett Jr JP, Wooten GF, Lothman EW. 1990.Autoradiographic evidence that NMDA receptor-coupled channelsare located postsynaptically and not presynaptically in the per-forant path-dentate granule cell system of the rat hippocampalformation. Brain Res 514:334–342.
Bettler B, Boulter J, Hermans-Borgmeyer I, O’Shea-Greenfield A,Deneris ES, Moll C, Borgmeyer U, Hollmann M, Heinemann S.1990. Cloning of a novel glutamate receptor subunit, GluR5; expres-sion in the nervous system during development. Neuron 5:583–595.
Bliss TV, Collingridge GL. 1993. A synaptic model of memory: long-term potentiation in the hippocampus. Nature 361:31–39.
Bureau I, Bischoff S, Heinemann SF, Mulle C. 1999. Kainate receptor-mediated responses in the CA1 field of wild-type and GluR6-defi-cient mice. J Neurosci 19:653–663.
Catania MV, Tolle T, Seeburg PH, Monyer H. 1995. Differentialexpression of AMPA receptor subunits in NOS-positive neurons ofcortex, striatum and hippocampus. J Neurosci 15:7046–7061.
Collingridge GL, Kehl SJ, McLennan H. 1983. Excitatory amino acidsin synaptic transmission in the Schaffer-commisural pathway of therat hippocampus. J Physiol 334:33–46.
Conti F, Minelli A, Brecha NC. 1994. Cellular localization and lami-nar distribution of AMPA glutamate receptor subunit mRNAs andproteins in the rat cerebral cortex. J Comp Neurol 350:241–259.
Cotman CW, Monaghan DT, Ganong AH. 1988. Excitatory amino acidneurotransmission: NMDA receptors and Hebb-type synaptic plas-ticity. Annu Rev Neurosci 11:61–80.
Cull-Candy SG, Brickley SG, Misra C, Feldmeyer D, Momiyama A,Farrant M. 1998. NMDA receptor diversity in the cerebellum: iden-tification of subunits contributing to functional receptors. Neuro-pharmacology 37:1369–1380.
Dewar KM. 1999. Alterations in serotonin receptors in the neostria-tum of weaver mutant mice. Neurochem Res 24:1449–1454.
Dure IV LS, Young AB. 1995. The distribution of glutamate receptorsubtypes in mammalian central nervous system using quantitativein vitro autoradiography. In: Stone TW. editor. CNS neurotrans-mitters and neuromodulators: glutamate. Boca Raton, FL: CRC. p83–94.
Frank L, Althoen SC. 1994. Statistics: concepts and applications. NewYork: Cambridge University Press.
Franklin KBJ, Paxinos G. 1997. The mouse brain in stereotaxic coor-dinates. San Diego: Academic Press.
Fuller TA, Russchen FT, Price JL. 1987. Sources of presumptiveglutamatergic/aspartatergic afferents to the rat ventral striatopal-lidal region. J Comp Neurol 258:317–338.
Geiger JRP, Melcher T, Koh DS, Sakmann B, Seeburg PH, Jonas P,Monyer H. 1995. Relative abundance of subunit mRNAs determinesgating and Ca21 permeability of AMPA receptors in principal neu-rons and interneurons in rat CNS. Neuron 15:193–204.
Greenamyre JT, Olson JMM, Penney JB, Young AB. 1985. Autora-diographic characterization of N-methyl-D-aspartate-, quisqualate-and kainate-sensitive glutamate binding sites. J Pharmacol ExpTher 233:254–263.
Griesser CAV, Cuenod M, Henke H. 1982. Kainic acid receptor sites inthe cerebellum of nervous, Purkinje cell degeneration, reeler, stag-gerer and weaver mice mutant strains. Brain Res 246:265–271.
Gupta M, Felten DL, Ghetti B. 1987. Selective loss of monoaminergicneurons in weaver mutant mice: an immunocytochemical study.Brain Res 402:379–382.
Halpain S, Wieczorek CM, Rainbow TC. 1984. Localization of L-glu-tamate receptors by quantitative autoradiography. J Neurosci4:2247–2258.
Hebert C, Habimana A, Elie R, Reader TA. 2001. Effect of chronicantidepressant treatments on 5-HT and NA transporters in ratbrain: an autoradiographic study. Neurochem Int 38:63–74.
Heckroth JA. 1994. Quantitative morphological analysis of the cere-bellar nuclei in normal and Lurcher mutant mice. I. Morphologyand cell number. J Comp Neurol 343:173–182.
Henke H, Beaudet A, Cuenod M. 1981. Autoradiographic localizationof specific kainic acid binding sites in pigeon and rat cerebellum.Brain Res 219:95–105.
Hirano T, Kasono K, Araki K, Mishina M. 1995. Supression of LTD incultured Purkinje cells dedicient in the glutamate receptor delta 2subunit. NeuroReport 6:524–526.
Insel TR, Miller LP, Gelhard RE. 1990. The ontogeny of excitatoryamino acid receptors in rat forebrain. I. N-methyl-D-aspartate andquisqualate receptors. Neuroscience 35:31–43.
Kashiwabuchi K, Ikeda K, Araki K, Hirano T, Shibuki K, TakayamaC, Inoue Y, Kutsuwada T, Yagi T, Kang Y, Aizawa S, Mishina M.1995. Impairment of motor coordination, Purkinje cell synapse for-mation, and cerebellar long-term depression in GluRd2 mutantmice. Cell 81:245–252.
Keinanen K, Wisden W, Sommer B, Werner P, Herb A, Verdoorn TA,Sakmann B, Seeburg PH. 1990. A family of AMPA-selective gluta-mate receptors. Science 249:556–560.
Kofuji P, Hofer M, Millen KJ, Millonig JH, Davidson N, Lester HA,Hatten ME. 1996. Functional analysis of the mutant weaver GIRK2potassium channel and rescue of weaver granule cells. Neuron16:941–952.
Lalonde R. 1986. Acquired immobility response in weaver mutantmice. Exp Neurol 94:808–811.
Lalonde R. 1987. Motor abnormalities in weaver mutant mice. ExpBrain Res 65:479–481.
Lalonde R, Botez MI. 1986. Navigational deficits in weaver mutantmice. Brain Res 398:175–177.
Lalonde R, Manseau M, Botez MI. 1988. Spontaneous alternation andexploration in weaver mutant mice. Behav Brain Res 31:111–114.
Lane JD, Nadi NS, McBride WJ, Aprison MH, Kusano K. 1977.Contents of serotonin, norepinephrine and dopamine in the cere-brum of the ‘staggerer’, ‘weaver’, and ‘nervous’ neurologically mu-tant mice. J Neurochem 29:349–350.
Lester HA, Karschin A. 2000. Gain of function mutants: ion channelsand G protein-coupled receptors. Annu Rev Neurosci 23:89–125.
Liu D, Diorio J, Day JC, Francis DD, Meaney MJ. 2000. Maternalcare, hippocampal synaptogenesis and cognitive development inrats. Nat Neurosci 3:799–806.
Lomeli H, Wisden W, Kohler M, Keinanen K, Sommer B, Seeburg PH.1992. High-affinity kainate and domoate receptors in rat brain.FEBS Lett 307:139–143.
Luque JM, Richards JG. 1995. Expression of NMDA 2B receptorsubunit mRNA in Bergmann glia. Glia 13:228–232.
Makowiec RL, Cha JJ, Penney JB, Young AB. 1991. Cerebellar exci-tatory amino acid binding sites in normal, granuloprival, and Pur-kinje cell-deficient mice. Neuroscience 42:671–681.
Miller LP, Johnson AE, Geldhard RE, Insel TR. 1990. The ontogeny ofexcitatory amino acid receptors in the rat forebrain. II. Kainic acidreceptors. Neuroscience 35:45–51.
Mjaatvedt AE, Cabin DE, Cole SE, Long LJ, Breitwieser GE, ReevesRH. 1995. Assessment of a mutation in the H5 domain of GIRK2 asa candidate for the weaver mutation. Genome Res 5:453–463.
Monaghan DT, Cotman CW. 1982. The distribution of [3H]kainic acidbinding sites in rat CNS as determined by autoradiography. BrainRes 252:91–100.
Monaghan DT, Cotman CW. 1985. Distribution of N-methyl-D-aspar-tate-sensitive L-[3H]-glutamate-binding sites in rat brain. J Neuro-sci 5:2909–2919.
IONOTROPIC GLUTAMATE RECEPTORS IN WEAVER MICE 231

Monaghan DT, Yao D, Cotman CW. 1984. Distribution of [3H]AMPAbinding sites in rat brain as determined by quantitative autoradiog-raphy. Brain Res 324:160–164.
Monaghan DT, Olverman HJ, Nguyen L, Watkins JC, Cotman CW.1988. Two classes of N-methyl-D-aspartate recognition sites: differ-ential distribution and differential regulation by glycine. Proc NatlAcad Sci USA 85:9836–9840.
Monyer H, Seeburg PH, Wisden H. 1991. Glutamate-operated chan-nels: developmentally early and mature forms arise by alternativesplicing. Neuron 6:799–810.
Monyer H, Burnashev N, Laurie DJ, Sackmann B, Seeburg PH. 1994.Developmental and regional expression in the rat brain and func-tional properties of four NMDA receptors. Neuron 12:529–540.
Morris RGM, Garrard P, Rawlins JNP, O’Keefe J. 1982. Place navi-gation is impaired in rats with hippocampal lesions. Nature 297:681–683.
Muller T, Grosche J, Ohlemeyer C, Kettermann H. 1993. NMDA-activated currents in Bergmann glial cells. Neuroreport 4:671–674.
Nase G, Weishaupt J, Stern P, Singer W, Monyer H. 1999. Geneticand epigenetic regulation of NMDA receptor expression in the ratvisual cortex. Eur J Neurosci 11:4320–4326.
Navarro B, Kennedy ME, Velimirovic B, Bhat D, Peterson AS,Clapham DE. 1996. Non-selective and Gbg-insensitive weaver K1
channels. Science 272:1950–1953.Oertel WH. 1993. Neurotransmitters in the cerebellum: scientific
aspects and clinical relevance. Adv Neurol 61:33–75.Olson JM, Greenamyre JT, Penney JB, Young AB. 1987. Autoradio-
graphic localization of cerebellar excitatory amino acid binding sitesin the mouse. Neuroscience 22:913–923.
Patil N, Cox DR, Bhat D, Faham M, Myers RM, Peterson AS. 1995. Apotassium channel mutation in weaver mice implicates membraneexcitability in granule cell differentiation. Nat Genet 11:126–129.
Petralia RS, Wang Y-X, Wenthold RJ. 1994. Histological and ultra-structural localization of the kainate receptor subunits KA2 andGluR6/7, in the rat nervous system using selective antipeptideantibodies. J Comp Neurol 349:85–110.
Petralia RS, Rubio ME, Wang Y-X, Wenthold RJ. 2000. Regional andsynaptic expression of ionotropic glutamate receptors. In: OttersenOP, Storm-Mathisen JT, editors. Handbook of chemical neuroanat-omy, vol. 18. Amsterdam: Elsevier. p 145–182.
Racca C, Catania MV, Monyer H, Sakmann B. 1996. Expression ofAMPA-glutamate receptor B subunit in rat hippocampal GABAer-gic interneurons. Eur J Neurosci 8:1580–1590.
Rainbow TC, Wieczorek CM, Halpain S. 1984. Quantitative autora-diography of binding sites for [3H]AMPA, a structural analogue ofglutamic acid. Brain Res 309:173–177.
Rakic P, Sidman RL. 1973a. Sequences of developmental abnormali-ties leading to granule cells deficit in cerebellar cortex of weavermutant mice. J Comp Neurol 152:103–132.
Rakic P, Sidman RL. 1973b. Organization of cerebellar cortex second-ary to deficits of granular cells in weaver mutant mice. J CompNeurol 152:133–161.
Rakic P, Sidman RL. 1973c. Weaver mutant mouse cerebellum: de-fective neuronal migration secondary to abnormality of Bergmannglia. Proc Natl Acad Sci USA 70:240–244.
Reader TA, Dewar KM. 1999. Effects of denervation and hyperinner-vation on dopamine and serotonin systems in the rat neostriatum:implications for human Parkinson’s disease. Neurochem Int 34:1–21.
Reader TA, Senecal J. 2001. Distribution of glutamate receptors of theNMDA subtype in brains of heterozygous and homozygous weavermutant mice. Neurochem Res 26:579–589.
Reader TA, Strazielle C. 1999. Quantitative autoradiography of mono-amine uptake sites and receptors in rat and mouse brain. In: Boul-ton AA, Bateson A, Baker GB, editors. Neuromethods, vol. 33. Cellneurobiology techniques. Totowa, NJ: Humana Press. p 1–51.
Reader TA, Ase AR, Hebert C, Amdiss F. 1999. Distribution ofdopamine, its metabolites, and D1 and D2 receptors in heterozy-gous and homozygous weaver mutant mice. Neurochem Res 24:1455–1470.
Reader TA, Hebert C, Ase AR, Le Marec N. 2001. Distribution ofserotonin, its metabolites and 5-HT transporters in the neostri-atum of Lurcher and weaver mutant mice. Neurochem Int 39:169 –177.
Reeves RH, Crowley MR, Lorenzon N, Pavan WJ, Smeyne RJ, Gold-owitz D. 1989. The mouse neurological mutant weaver maps withinthe region of chromosome 16 that is homologous to human chromo-some 21. Genomics 5:522–526.
Rezai Z, Yoon CH. 1972. Abnormal rate of granule cell migration inthe cerebellum of “weaver” mutant mice. Dev Biol 29:17–26.
Richter JA, Stotz EH, Ghetti B, Simon JR. 1992. Comparison ofalterations in tyrosine hydroxylase, dopamine level and dopamine
uptake in the striatum of the weaver mutant mouse. NeurochemRes 17:437–441.
Roffler-Tarlov S, Graybiel AM. 1984. Weaver mutation has differen-tial effects on the dopamine-containing innervation of the limbicand nonlimbic striatum. Nature 307:62–66.
Roffler-Tarlov S, Graybiel AM. 1987. The postnatal development ofthe dopamine-containing innervation of dorsal and ventral stria-tum: effects of the weaver gene. J Neurosci 7:2364–2372.
Roffler-Tarlov S, Sidman RL. 1978. Concentrations of glutamic acid incerebellar cortex and deep nuclei of normal mice and weaver, stag-gerer and nervous mutants. Brain Res 142:269–283.
Roffler-Tarlov S, Turey M. 1982. The content of amino acids in thedeveloping cerebellar cortex and deep cerebellar nuclei of granulecell deficient mutant mice. Brain Res 247:65–73.
Rudolf GD, Cronin CA, Landwehrmeyer GB, Standaert DG, PenneyJr JB, Young AB. 1996. Expression of N-methyl-D-aspartate gluta-mate receptor subunits in the prefrontal cortex of the rat. Neuro-science 73:417–427.
Sakurai SY, Cha J-HJ, Penney JB, Young AB. 1991. Regional distri-bution and properties of [3H]MK-801 binding sites determined byquantitative autoradiography in rat brain. Neuroscience 40:533–543.
Sakurai SY, Penney JB, Young AB. 1993. Regionally distinct N-methyl-D-aspartate receptors distinguished by quantitative autora-diography of [3H]MK-801 binding in rat brain. J Neurochem 60:1344–1353.
Sato K, Kiyama H, Tohyama M. 1993. The differential expression ofmessenger RNAs encoding non-N-methyl-D-aspartate glutamate re-ceptor subunits (GluR1–4) in the rat brain. Neuroscience 52:515–539.
Schiffer HH, Swanson GT, Heinemann SF. 1997. Rat GluR7 and acarboxy-terminal splice variant, GluR7b, are functional kainatereceptor subunits with a low sensitivity to glutamate. Neuron 19:1141–1146.
Schmidt MJ, Sawyer BD, Perry KW, Fuller RW, Foreman MM, GhettiB. 1982. Dopamine deficiency in the weaver mutant mouse. J Neu-rosci 2:376–380.
Sidman RL. 1968. Development of interneuronal connections inbrains of mutant mice. In: Carlson FD, editor. Physiological andbiochemical aspects of nervous integration. Englewood Cliffs, NJ:Prentice Hall. p 163–193.
Slesinger PA, Patil N, Liao YJ, Jan LY, Cox DR. 1996. Functionaleffects of the weaver mutation on G protein-gated inwardly rectify-ing K1 channels. Neuron 16:321–331.
Smeyne RT, Goldowitz D. 1989. Development and death of externalgranular layer cells in the weaver mouse cerebellum: a quantitativestudy. J Neurosci 9:1608–1620.
Smeyne RT, Goldowitz D. 1990. Purkinje cell loss is due to a directaction of the weaver gene in Purkinje cells: evidence from chimericmice. Dev Brain Res 52:211–218.
Sotelo C. 1975. Anatomical, physiological and biochemical studies ofthe cerebellum from mutant mice. II. Morphological study of cere-bellar cortical neurons and circuits in the weaver mouse. Brain Res94:19–44.
Sotelo C, Changeux JP. 1974. Bergmann fibres and granular cellmigration in the cerebellum of homozygous weaver mutant mouse.Brain Res 77:484–491.
Standaert DG, Landwehrmeyer GB, Kerner JA, Penney Jr JB, YoungAB. 1996. Expression of NMDAR2D glutamate receptor subunitmRNA in neurochemically identified interneurons in the rat neo-striatum, neocortex and hippocampus. Mol Brain Res 42:89–102.
Standaert DG, Friberg IK, Landwehrmeyer GB, Young AB, Penney JrJB. 1999. Expression of NMDA glutamate receptor subunit mRNAsin neurochemically identified projection and interneurons in thestriatum of the rat. Mol Brain Res 64:11–23.
Stefani A, Chnen Q, Flores-Hernandez J, Jiao Y, Reiner A, SurmeiirDI. 1998. Physiological and molecular properties of AMPA/kainatereceptors expressed by striatal spiny neurons. Dev Neurosci 20:242–252.
Stotz EH, Triarhou LC, Ghetti B, Simon JR. 1993. Serotonin contentis elevated in the dopamine deficient striatum of the weaver mutantmouse. Brain Res 606:267–272.
Strazielle C, Lalonde R, Amdiss F, Botez MI, Hebert C, Reader TA.1998. Distribution of dopamine transporters in basal ganglia ofcerebellar ataxic mice by [125I]RTI-121 quantitative autoradiogra-phy. Neurochem Int 32:61–68.
Strazielle C, Lalonde R, Reader TA. 2000. Autoradiography of gluta-mate receptor binding in adult Lurcher mutant mice. J NeuropatholExp Neurol 59:707–722.
Surmeier DJ, Mermelstein PG, Goldowitz D. 1996. The weaver mu-tation of GIRK2 results in a loss of inwardly rectifying K1 current
232 T.A. READER AND J. SENECAL

in cerebellar granule cells. Proc Natl Acad Sci USA 93:11191–11195.
Tang Y-P, Shimizu E, Dube GR, Rampon C, Kerchner GA, Zhuo M,Liu G, Tsien JZ. 1999. Genetic enhancement of learning and mem-ory in mice. Nature 401:63–69.
Tong Y, Wei J, Zhang S, Strong JA, Dlouhy SR, Hodes ME, Ghetti B,Yu L. 1996. The weaver mutation changes the ion selectivity of theaffected inwardly rectifying potassium channel GIRK2. FEBS Lett390:63–68.
Triarhou LC, Ghetti B. 1987. Neuroanatomical substrate of behav-ioural impairment in weaver mutant mice. Exp Brain Res 68:434–436.
Triarhou LC, Norton J, Ghetti B. 1988. Mesencephalic dopaminedeficit involves areas A8, A9 and A10 in weaver mutant mice. ExpBrain Res 70:256–265.
Unnerstall JR, Wamsley JK. 1983. Autoradiographic localization ofhigh-affinity [3H]kainic acid binding sites in the rat forebrain. EurJ Pharmacol 86:361–371.
Unnerstall JR, Niehoff DL, Kuhar MJ, Palacios JM. 1982. Quantita-tive receptor autoradiography using [3H]ultrafilm: application tomultiple benzodiazepine receptors. Neurosci Meth 6:59–73.
Verina T, Norton J, Sorbel JJ, Triarhou LC, Laferty D, Richter JA,Simon JR, Ghetti B. 1997. Atrophy and loss of dopaminergic mes-encephalic neurons in heterozygous weaver mice. Exp Brain Res113:5–12.
Verney C, Febvret-Muzerelle A, Gaspar P. 1995. Early postnatalchanges of the dopaminergic mesencephalic neurons in the weavermutant mouse. Dev Brain Res 89:115–119.
Voogd J, Jaarsma D, Marani E. 1996. The cerebellum: chemoarchi-tecture and anatomy. In: Swanson LW, Bjorklund A, Hokfelt T,
editors. Handbook of chemical neuroanatomy: integrated systems ofthe CNS. Part III, cerebellum, basal ganglia, olfactory system.Amsterdam: Elsevier. p 1–369.
Watanabe M. 1997. Developmental dynamics of gene expression forNMDA receptor channel. In: Monaghan DT, Wenthold RJ, editors.The ionotropic glutamate receptors. Totowa, NJ: Humana Press. p189–218.
Watanabe M, Inoue Y, Sakimura K, Mishina M. 1993. Distinct dis-tributions of five N-methyl-D-aspartate receptor channel subunitmRNAs in the forebrain. J Comp Neurol 338:377–390.
Watanabe M, Mishima M, Inoue Y. 1994. Distinct spatiotemporalexpressions of the NMDA receptor channel subunit mRNAs in thecerebellum. J Comp Neurol 343:513–519.
Whishaw IQ. 1998. Place learning in hippocampal rats and the pathintegration hypothesis. Neurosci Biobehav Rev 22:209–220.
Wisden W, Seeburg PH. 1993. A complex mosaic of high-affinitykainate receptors exists in rat brain. J Neurosci 13:3582–3598.
Wisden W, Seeburg PH, Monyer H. 2000. AMPA, kainate and NMDAionotropic glutamate receptor expression—an in situ hybridizationatlas. In: Ottersen OP, Storm-Mathisen JT, editors. Handbook ofchemical neuroanatomy, vol. 18. Glutamate. Amsterdam: Elsevier.p 99–143.
Wood ER, Dudchenko PA, Eichenbaum H. 1999. The global record ofmemory in hippocampal neuronal activity. Nature 397:561–563.
Zhao HM, Wenthold RJ, Petralia RS. 1998. Glutamate receptor tar-geting to synaptic populations on Purkinje cells is developmentallyregulated. J Neurosci 18:5517–5528.
Zuo J, De Jager PL, Takahashi KA, Jiang W, Linden DJ, Heintz N.1997. Neurodegeneration in Lurcher mutant mice caused by muta-tion in d2 glutamate receptor gene. Nature 388:769–773.
IONOTROPIC GLUTAMATE RECEPTORS IN WEAVER MICE 233









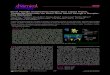






![Design, synthesis and application of carbazole macrocycles ...€¦ · Anion receptors containing carbazole and amide functionalities were investigated in numerous works [6-9]. In](https://img.pdfslide.fr/doc/110x75/605e9713d39a752cd71609f2/design-synthesis-and-application-of-carbazole-macrocycles-anion-receptors-containing.jpg)












