Embed Size (px)
Citation preview

Eur. J . Biochem. 58, 327-337 (1975)
Ultrastructural Organization and Biochemical Characterization of Chromatin - RNA Protein Complexes Isolated from Mammalian Cell Nuclei Jean-Pierre BACHELLERIE, Edmond PUVION, and Jean-Pierre ZALTA
Centre de Recherche en Biochimie et Genetique Cellulaires du Centre National de la Recherche Scientifique, Toulouse, and Institut de Recherches Scientifiques sur le Cancer, Villejuif
(Received February 10/June 9, 1975)
Extranucleolar elements were isolated from mammalian cell nuclei and characterized by correlative biochemical-ultrastructural studies. These complex structures were shown to retain a supra-particle arrangement closely resembling the organizaton in situ of definite nuclear areas. In particular the morphological RNA . protein species, namely perichromatin fibrils and granules, were detected in association with characteristic regions of chromatin. By autoradiographic experi- ments, we were able to show that perichromatin fibrils represent the morphological state of newly formed heterogeneous nuclear RNA (hnRNA).
Chromatin . RNA . protein complexes were further fractionated by means of a treatment dissociating the chromatin component. A minor part of DNA, possibly involved in the biosynthesis and the processing of hnRNA, remains linked to the resulting RNA . protein network.
Most of the mechanisms involved in the control of RNA maturation in eukaryotic cells still remain to be elucidated. It has now been clearly established that a class of heterogeneous nuclear RNA (hnRNA) of high molecular weight, rapidly labeled and with a DNA-like composition [l-'51, presents a number of characteristics of cytoplasmic mRNA precursors [6,7]. Most of this hnRNA is degraded in the nucleus and never reaches the cytoplasm [l, 31. However recent findings indicate that all the hnRNA molecules cannot be considered simply as direct precursors for mRNA
By both cytological [lo, 111 and biochemical [12- 151 studies, this hnRNA has been detected in the form of RNA . protein particles. The biochemical characteri- zation, particularly of the protein moiety of hnRNA- containing RNA . protein complexes, isolated by various procedures, has been undertaken by several groups [12- 151. The ultrastructural study of inter- phase nuclei [lo] has shown that the various morpho- logical RNA . protein species, particularly the so- called perichromatin fibrils, perichromatin and inter- chromatin granules [lo, 16,171, were topographically distributed in situ according to characteristic patterns which could be correlated to various functional states.
The present study was undertaken in order to establish tentative correlation between biochemical and ultrastructural features of nuclear RNA protein particles and their spatial organization in situ.
Abbreviation. hnRNA, heterogeneous nuclear RNA.
~ 9 1 .
We developed an isolation procedure of subnuclear fractions where the ultrastructural organization of definite RNA . protein . chromatin areas appears well preserved. This method could contribute to the characterization of actively transcribed zones in chro- matin as well as to a better definition of the events involved in hnRNA metabolism. We here report direct evidence that perichromatin fibrils do indeed represent the morphological state of newly synthe- sized hnRNA, as previously assumed by autoradio- graphic studies in situ [18].
MATERIALS AND METHODS
Cell Culture and Labeling Zadjela Ascitic Hepatoma Cells. Female Wistar rats
were inoculated intraperitoneally with approximately 2 x lo7 cells. After 3 days, the cells were collected, washed repeatedly in a phosphate-buffered saline medium (NaC10.12 M, KCl4 mM, Na2HP04 7 mM, KH2P0, 1 mM, NaHCO, 16 mM) and resuspended in Fischer [19] medium (Eurobio), supplemented with 5 % calf serum, 100 U/ml penicillin and 50 pg/ml streptomycin. The cell suspension was then incubated at 37 "C in glass flasks at concentrations varying from 5 x 105/ml to 106/ml according to the experiments planned. Cells were labeled after a 5-h incubation in synthetic medium by addition of radioactive pre- cursors.

328 Isolation of Native-Like Chromatin . RNA . Protein Complexes
Chinese Hamster Ovary (CHO) Cells. Chinese hamster cells, kindly provided by Dr L. Siminovitch (Toronto), were grown, in Eagle [2] minimum essen- tial medium (Eurobio) supplemented with 100 U/ml penicillin, 50 pg/ml streptomycin and 10 x calf serum (Flow), in spinner suspension, maintained in the 3 - 6 x 1OS/ml concentration range. Whenever auto- radiographic studies were planned, [3H]uridine was added at 100 pCi/ml.
Labelings were stopped by pouring the suspension on crushed iced Earle's saline [21]. Cells were harvested by centrifugation at 500 x g , and twice washed in cold Earle's saline.
Nuclei isolation and disruption were carried out according to the previously described procedure [22].
Cell Fractionation. All operations were carried out at 0-4"C, unless otherwise stated. 1 ml (3 x lo8) packed cells were resuspended in 10ml buffer N (0.25 M sucrose, 10 mM Tris pH 7.4, 2.5 mM MgCl,, 0.1 mM CaCI,, 20 pg/ml potassium polyvinyl sul- phate) in the presence of non-ionic detergents, cemul- sol NPT 10 (0.4%, v/v) and celanol 251 (0.1 x, v/v), and 0.5 mg/ml collagenase (Calbiochem, Bacterial B gradc). The suspension was homogenized with an Ultra-Turrax (Janke and Kunkel, Staufen, Germany) functioning at 60 V (about 10000 rev./min) until no unbroken cell could be seen under the phase-contrast microscope and the whole nuclei appear freed from cytoplasmic tags (about 3 min). Crude nuclei were collected by a 5-min centrifugation at 150xg in conical glass tubes, resuspended in 10 ml buffer N and the suspension homogenized as previously for 1 min. Purified nuclei were collected by a 5-min centri- fugation at 450 x g . The nuclear pellet was then re- suspended in 12 ml medium A (Ficoll 2.1 %, w/v, 10 mM Tris-HC1 pH 7.4, 5.5 mM MgCl,, 0.25 mM 2-mercaptoethanol, 10 pg/ml potassium polyvinyl sul- phate and the suspension, in a 50-ml polypropylene tube, was submitted to a 90-s sonication (10-s waves with 20-s rest in between) resulting in a complete disruption of nuclei. 12 ml medium B (identical to A except for 0.5 mM Mg') was then added in order to lower the Mg2+ concentration to 3 m M and an additional 90-s sonication step was carried out to achieve the dispersion of most extranucleolar chro- matin aggregates while nucleolar structure were pre- served. Sonication was performed with the Raytheon sonic oscillator (DF 101, 10 kHz, 200 W) with re- frigeration through circulating ethanol at - 15 "C. Fractionation of the dispersed nuclear components was then carried out, by a series of differential centri- fugation, according to Table 1.
RNA Extraction and Analysis
Nucleolar RNAs were extracted according to Nakamura [23]. After homogenization of the isolated
nucleoli in acetate buffer (0.05 M CH3COOK pH 5.1, 0.14 M NaCI, 1 % sodium dodecyl sulphate, 20 pg/ml potassium polyvinyl sulphate, an equal volume of acetate-buffer-saturated phenol containing 0.1 % 8-hydroxyquinoleine was added and the mixture vigor- ously shaken for 10 min at 65 "C.
The emulsion was then broken by centrifugation at room temperature and the aqueous phase again extracted with phenol by shaking for 15 rnin at room temperature. RNAs from extranucleolar fractions (P,,, Ploo and S,,,) were extracted according to Perry et al. [24] by a dodecylsulphate/chloroform treatment at room temperature with strong agitation on aVortex- mixer, This procedure has been shown to prevent considerable loss of poly(A)-containing RNAs and cleavage of the poly(A) sequences in vitro that can occur in the course of this extraction. RNAs from the recovered aqueous phases, brought to 0.2 M CH,COOK (pH 5.1), were precipitated by addition of two volumes of ethanol and allowed to stand over- night at - 20°C. The RNA precipitates collected by centrifugation were dissolved in high-ionic-strength buffer (0.5 M NaCl, 10mM Tris pH 7.4, 50mM MgC1,) and submitted to a 20-min treatment with DNase 1 (Worthington, RNAase-free), (50 pg/ml) at 0 "C before dilution in buffer E (see below) containing 20 % sucrose and analysed by electrophoresis.
Polyacrylamide gel electrophoresis was performed according to Tiollais et al. [25]. Samples of 100 p1 (containing up to 100 pg RNA) were layered on top of 21 x 0.6-cm cylindrical gels made up with 1.7% polyacrylamide/0.5 % agarose in buffer E (containing in 1 1, 10.8 g Tris, 0.93 g Na,EDTA, 5.5 g boric acid, 0.2% sodium dodecyl sulphate, pH 8.3). Separation was achieved by migration, for 290min, under a lO-V/cm electric field, when the bromphenol blue marker reached the bottom of the gel. The radio- activity profile was then determined after a 2-mm slicing of the gel, performed either by the useful manual device described by Tiollais [25] or by the automatic Gilson fractionator. The gel section of RNA was hydrolyzed before counting by an overnight treatment with 1 ml 10% NH,OH.
Ultrastructural Study
Fixation. The pellets were fixed in 1.6% glutar- aldehyde in Sorensen buffer (0.1 M phosphate pH 7.3 at 4°C) for 30-60 min. This time was reduced to 10 min when glycomethacrylate embedding was re- quired for enzymic digestions.
Conventional Techniques. The glutaraldehyde-fixed pellets were embedded in epon after alcohol de- hydratation without osmium postfixation. When some pellets remained dispersed even after prolonged cen- trifugation in the fixative buffer, it was useful to encapsulate them before dehydratation in a droplet

J.-P. Bachellerie, E. Puvion, and J.-P. Zalta 329
of 3% agar at 45°C. For enzymic digestions, pellets were embedded in glycolmethacrylate according to Leduc and Bernhard [26]. In the conventional tech- nique ultrathin sections were stained with uranyl acetate and lead citrate.
Preferential Staining of RNA. Protein and Enzymic Digestions. Epon sections were stained by the uranyl acetate- EDTA- lead citrate sequence according to Bernhard [27]. For ultrathin frozen sections, a small fraction of the pellet was mixed with “Tissue-Tek” [28], frozen and sectioned with the Sorvall MT 2B ultra- microtome equipped with a cryokit. The sections spread on a 50% solution of dimethylsulfoxide in water were then transferred to water by means of plastic rings [29]. After mounting on formvar carbon- coated grids, staining was carried out according to the technique of RNA . protein preferential staining modified by Puvion and Bernhard [30]. Enzymic digestions were performed on ultrathin frozen sections or on ultrathin sections of glycomethacrylate-em- bedded material. In both cases sections in plastic rings were floated for 1 to 5 h on solutions of RNAse (0.1 ”/, in distilled water pH 7) or on solutions of pepsin (0.5 % in 0.1 M HCl) followed by RNAse. After enzymic digestions, final staining was carried out as above. Control sections were floated for the same time on the same solutions without en- zymes.
Autoradiography. Ilford L, emulsion was applied on single grids using the gold interference coloured zone of a platinum loop [31]. The autoradiographs were developed after 9 months (first experiment) or 2 months (second experiment) in a phenidon-con- taining developer. The preferential staining for RNA . protein elements was carried out on dry grids.
Radioisotopes. [Me-3H]Thymidine (52 Ci/mmol) and [5-3H]uridine (23 Ci/mmol) were purchased from C.E.A. (Saclay-France). [14C]Uridine (514 mCi/mmol) was purchased from the Radiochemical Centre (Amersham, England).
RESULTS
Nuclei disruption was achieved by sonication in low-ionic-strength medium containing a suitable Mg2 +
concentration (3 mM). This resulted in a good preser- vation of nucleolar structures and allowed their separation from extranucleolar components. The pres- ence of Ficoll has been shown to favour chromatin dispersion [22]. Nuclear components were separated by a series of differential centrifugation. Purified nucleoli were recovered by a 500 x g centrifugation, the supernatant being freed from remaining nucleoli by a subsequent 1500 x g centrifugation. Extranu- cleolar chromatin fractions, namely P,, and P,,,, were obtained as described in Table 1.
Table 1. Fractionation scheme Isolated nuclci were resuspended in medium A (containing 5.5 m M MgCIJ and submitted to a first sonication cycle. MgCI, concen- tration was then lowered to 3 mM and a second sonication step was carried out
Starting Centrifugation Pellet obtained Super- material natant
Isolated nuclei 5 min Po.5, pure isolated
s0.5 6 min P, 5 , isolated nucleoli at 500 x g nucleoli S0.S
lar chromatin S,,S at 1500 x g and some extranucleo-
s1 .5 10 min PI,, structured extra-
s25 60 min PI,, more dissociated a t 25000 x g nucleolar chromatin SZ5
at 100000 xg extranucleolar chromatin Sloe
Table 2. Distribution of RNA and DNA among subnuclear fractions Chinese hamster ovary cells doubly-labeled by a 4-h pulse with [3H]thymidine (1 pCi/ml) and [’4C]uridine (0.1 pCi/ml) and sub- nuclear fractionation was carried out according to Table 1. Radio- activity precipitable by trichloroacetic acid was determincd on aliquots of each fraction. Results are expressed as percentage of total RNA and DNA in the nuclei
Fraction Nuclear RNA Nuclear DNA
% ~ ~~
Nucleoli P”, + P I S 42 15
Extranucleolar P2S 33.6 56.5 PlUO 23.2 26 s,,, 1.2 2.5
RNA and DNA contents in these nuclear sub- fractions were determined on 4-h [14C]uridine and [3H]thymidine pulsed Chinese hamster ovary cells. The results (Table2) are an average of three deter- minations. This distribution is quite reproducible and was not significantly modified when a shorter pulse time (15 min) was used.
The major part of extranucleolar nuclear RNA and DNA was recovered in the form of rapidly sedimenting elements (PZ5) while the remainder was found in the P,,, fraction. Only a very small part, about 2 x, 6f the nuclear nucleic acids were recovered in the elements that did not pellet at 100000 x g .
Ultrastructural Features of Extranucleolar Chromatin . R N A . Protein Elements
After conventional double staining, the two pel- leted fractions appeared similar. They both consisted of heavily stained fibrillar bundles which appeared more dense and irregularly disposed in the faster-
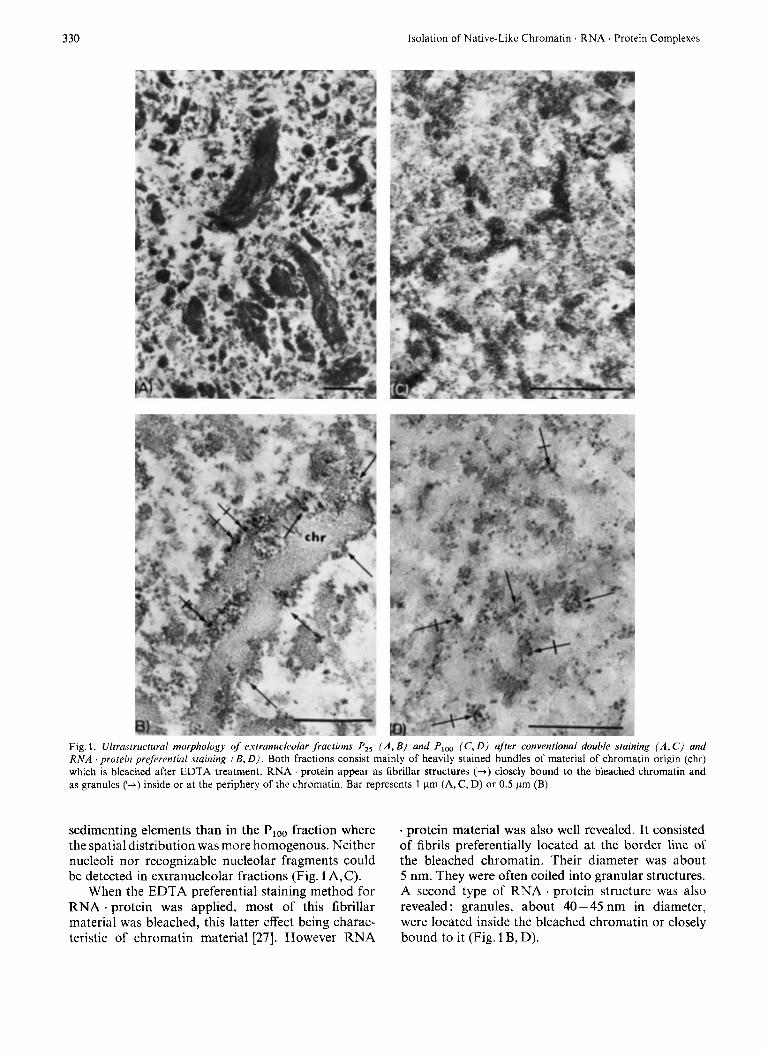
330 Isolation of Native-Likc Chromatin RNA . Protein Complexes
Fig. 1. ULtra.structural morphology oJ extranucleolar Jractions P,, ( A , B ) and Ploo (C , 0) after conventional double staining ( A , C ) and RNA 'protein preferential staining (B , D ) . Both fractions consist mainly of heavily stained bundles of material of chromatin origin (chr) which is bleached after EDTA treatment. RNA . protein appear as fibrillar structures (-+) closely bound to the bleached chromatin and as granules (I-) inside or at the periphery of the chromatin. Bar represents 1 pm (A, C, D) or 0.5 pm (B)
sedimenting elements than in the Plo0 fraction where the spatial distribution was more homogenous. Neither nucleoli nor recognizable nucleolar fragments could be detected in extranucleolar fractions (Fig. 1 A, C).
When the EDTA preferential staining method for RNA . protein was applied, most of this fibrilpar material was bleached, this latter effect being charac- teristic of chromatin material 1271. However RNA
. protein material was also well revealed. It consisted of fibrils preferentially located at the border line of the bleached chromatin. Their diameter was about 5 nm. They were often coiled into granular structures. A second type of RNA . protein structure was also revealed: granules, about 40-45 nm in diameter, were located inside the bleached chromatin or closely bound to it (Fig. 1 B, D).

J.-P. Bachellerie, E. Puvion, and J.-P. Zalta 331
Extranucleolar Origin of Pzs and PI,, Fractions
An important prerequisite for further study was to test whether or not these fractions are really of extra- nucleolar origin. Although ultrastructural observa- tions do not allow the detection of nucleoli or recog- nizable nucleolar fragments, biochemical controls have been undertaken to clear up this point.
The experiment described in Table 3 shows that at least 81 % of the nucleoli initially present in un- broken nuclei are thus recovered in Po,, and PI., in the form of morphologically intact nucleolar struc- tures. The remaining 19% of nucleoli cannot be detected directly in PZ5 and PI,,. We can therefore assume that these nucleoli were lost in the course of decantation or more or less disrupted during the soni- cation and consequently recovered according to their size in the various successive pellets, including the operationally defined nucleolar fractions (Po,s and PIS).
Consequently, the maximum value for a possible nucleolar contamination in extranucleolar fractions is 19%. In relation to this problem more interesting results were achieved by the experiment described in Table 4. When Chinese hamster ovary cells were treated by low doses of actinomycin D, at concentra- tions that are known to inhibit specifically rRNA bio- synthesis [32], a clear uncoupling was obtained in the response of “nucleolar” (Po,, and and “extra- nucleolar” (PZ5 and Plo0) fractions: a 10-min pre- treatment with 0.1 pg/ml actinomycin D resulted in a 60% inhibition of RNA labeling in nucleolar frac- tions, whereas the labeling in extranucleolar fractions was not depressed. (In fact a slight increase, which could be significant, was observed.)
Table 3. Yield determination ,for isolated nucleoli with our nuclear fructionution procedure Nuclei were isolated from about 2 x 10’ Chinese hamster ovary cells as described in Materials and Methods. Precise determination of the number of nuclei in the nuclear suspension before sonication (1.95 x 10’) was achieved with the help of a counting chamber. The average number of nucleoli per nucleus (1.75) was obtained from 169 determinations on the nuclear suspension. The number of nucleoli recovered in P,, and P,,, was determined as above after resuspension in Tris/Mg/K buffer (Tris-HCI 10 mM, pH 7.4, MgCl, 3 mM, KC1 20 mM)
Fraction Number of nucleoli
Original 3 . 4 0 ~ lo8 (100%)
PI., 1.53 x 10’ (46%) Po., + PI., 2.73 x 10’ (81 %,)
Po 5 1.20 x 10’ (35 %)
Table 4. Selective inhibition of’ R N A biosynthesis in various sub- nuclear,fractions by actinomycin D (0.1 pglml) 10’ Chinese hamster ovary cells were treated by 0.1 pg/ml actino- mycin D for 10 miu before a 15-min pulse with [3H]uridine (2 pCi/ml). A control (lo8 cells) was similarly labeled, actinomycin treatment being omitted. On the other hand, about 10’ Chinese hamster ovary cells were labeled for 4 h with [14C]uridine (0.1 pCi/ ml). Half of the ‘‘C-labeled cells were added to each ’H-labeled cell pellet (control and actinomycin-D-treated) before fractionation, in order to check that the RNA distribution among nuclear sub- fractions was quantitatively reproducible. Acid-precipitable radio- activity was determined on aliquot of each fraction. Results are expressed in relative variations of radioactivity in each fraction (actinomycin-D-treated cells compared with control)
Fraction Radioactivity
”/, Whole nuclei - 17 Nucleoli Po,5 + P,,, - 60 pzi + I p,oo + 18
R N A Analysis of Extrunucleolur Fractions
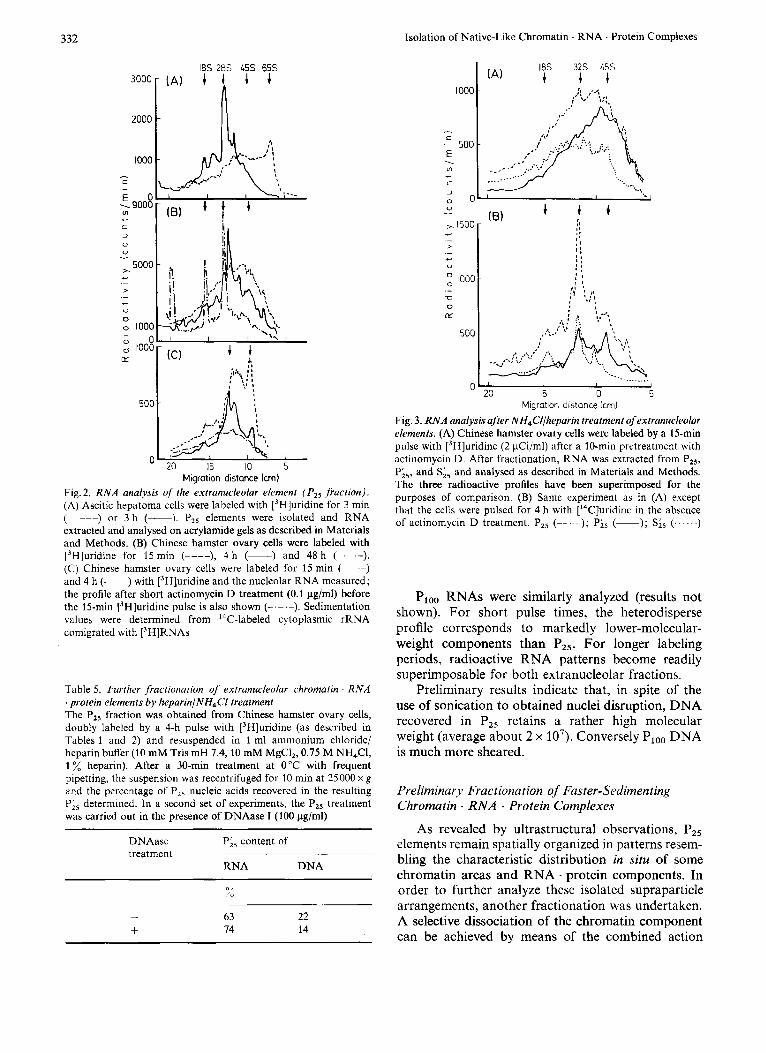
The kinetics of RNA labeling have been studied on both ascitic hepatoma and Chinese hamster ovary cells. RNAs were extracted from different subnuclear fractions and analysed on agarose-polyacrylamide gels. Labeling profiles for hepatoma P,, RNAs are depicted in Fig.2A. For a 3-min pulse with [3H]uridine, the radioactivity pattern was heterodisperse and pre- dominantly located in high-molecular-weight areas (20-70 S). A 65-S radioactivity peak was clearly distinguishable from the polydisperse profile. When cells were labeled by a 3-h pulse with [jH]uridine, the radioactivity pattern was shifted towards lower- molecular-weight areas and the polydispersity was clearly reduced : discrete RNA species coincided with known preribosomal RNA intermediates. The sedi- mentation values were determined from I4C-labeled cytoplasmic rRNA comigrated with P,, [3H]RNAs.
Results obtained for Chinese hamster ovary cells are shown in Fig. 2B. So far the shortest pulse time we have studied is 15 min: the corresponding radio-
activity pattern for RNAs was heterodisperse and slightly shifted towards lower-molecular-weight com- ponents when compared to the profile corresponding to the 3-min pulsed hepatoma cells. For longer pulse times, discrete species became predominantly labeled as observed for hepatoma cells, the 4-h profile in Fig. 2 B being almost superimposable on the 3-h profile in Fig.2A. The 15-min pattern remained exactly the same when Chinese hamster ovary cells were pretreated with 0.1 pg/ml actinomycin D. On the contrary this treatment has been shown to severely inhibit nucleolar labeling (Table 4).
As a control, nucleolar RNA patterns (Po.5 + from Chinese hamster ovary cells) are shown inFig.2C. Characteristic preribosomal RNA species are clearly detected, the shift of labeling from the 45-S to 28-S area in the 0.25 - 4-h interval corresponding to the known maturational cleavages of preribosomal RNAs.
The effect of actinomycin D treatment on the nucleolar RNA pattern can also be seen in Fig. 2C.
332 Isolation of Native-Like Chromatin . RNA . Protein Complexes
2000
1000 - c - E D \9000 m
c 3
0 U
I
- 5000 x
c .- > - - U
0 0 1000 G O 0 1000
LT
50C
C 0 15 10 5
Migration distance lcm) Fig. 2. R N A analysis of !he extrunucleolar element (P , , fraction). (A) Ascitic hepatoma cells were labeled with [3H]uridine for 3 min (-----) or 3 h (-). P,, elements wcre isolated and RNA extracted and analysed on acrylamide gels as described in Materials and Methods. (B) Chinese hamster ovary cells were labeled with [3H]uridine for 15 min (----), 4 h (-) and 48 h (-.-.-). (C) Chinese hamster ovary cells were labeled for 15 min ( --) and 4 h (-) with [3H]uridine and the nucleolar RNA measured; the profile after short actinomycin D treatment (0.1 pg/ml) before the 15-min [3H]uridine pulse is also shown (-.-.-). Sedimentation values were determined from 14C-labeled cytoplasmic rRNA comigrated with ['HH]RNAs
Table 5. Further fractionation of' extranucleolar chromatin . R N A . protein elements by heparin/NH,CI treatment The P,, fraction was obtained from Chinese hamster ovary cells, doubly labeled by a 4-h pulse with [3H]uridine (as described in Tables 1 and 2) and resuspended in 1 ml ammonium chloride/ heparin buffer (10 mM Tris mH 7.4,lO mM MgCI,, 0.75 M NH4CI, 1 heparin). After a 30-min treatment at 0°C with frequent pipetting, the suspension was recentrifuged for 10 min at 25000 x g and the percentage of P,, nucleic acids recovered in the resulting P;5 determined. In a second set of experiments, the P,, treatment was carried out in the presence of DNAase I (100 pg/ml)
DNAasc Pi, content of treatment
RNA DNA
- +
63 22 14 14
1000 :: ,'4 -
I
C .- 500 E
-
Ln . c 3
.... .... .... - 0 0; ' I I U I
1500 c - > .- c
U
1000 ._ m 0 IX
500
0 .
::
20 15 10 5 Migration distance Icm)
Fig. 3. RNA analysis after NH,C'l/lieparin treatmeni of extranucleolar elements. (A) Chinese hamster ovary cells were labeled by a 15-min pulse with [3H]uridine (2 pCi/ml) after a 10-min pretreatment with actinomycin D. After fractionation, RNA was extracted from P,,, P;5, and S;, and analysed as described in Materials and Methods. The three radioactive profiles have been superimposed for the purposes of comparison. (B) Same experiment as in (A) except that the cells were pulsed for 4 h with [14C]uridine in the absence of actinomycin D trcatment. PIS (-- ); Pis (-); S;, (...... )
Ploo RNAs were similarly analyzed (results not shown). For short pulse times, the heterodisperse profile corresponds to markedly lower-molecular- weight components than PZ5. For longer labeling periods, radioactive RNA patterns become readily superimposable for both extranucleolar fractions.
Preliminary results indicate that, in spite of the use of sonication to obtained nuclei disruption, DNA recovered in Pz5 retains a rather high molecular weight (average about 2 x lo7). Conversely Ploo DNA is much more sheared.
Preliminary Fractionution of Faster-Sedirnenting Chromatin . RNA . Protein Complexes
As revealed by ultrastructural observations, PZs elements remain spatially organized in patterns resem- bling the characteristic distribution in situ of some chromatin areas and RNA . protein components. In order to further analyze these isolated supraparticle arrangements, another fractionation was undertaken. A selective dissociation of the chromatin component can be achieved by means of the combined action

J.-P. Bachellerie, E. Puvion, and J.-P. Zalta 333
of a high concentration (0.75 M) of NH4Cl (a treat- ment known to result in the release of F1 histone from chromatin) and of heparin, a polyanion that has strong affinities with basic proteins. As indicated in Table 5, the 25 000 x g pellet recovered from the treated suspen- sion of PZs elements, namely P;5, contains the major part of 4-h labeled RNA and only a minor part of DNA : the RNA/DNA ratio is about three times higher than in primitive PZ5. When this treatment was carried out in the presence of 100 pg/ml of DNAase I, the uncoupling between DNA and RNA separation was more pronounced: RNA/DNA ratio in Pi5 was five times higher than in primitive P25. It is important to note that, even in this latter case, a significant portion (14%) of the DNA remained linked to the complex structures recovered in P;,.
However, for a shorter labeled uridine pulse (15 min, after a 0.1 g / m l actinomycin D treatment) the percentage of P2, labeled RNA released in S;, was significantly reduced: 24 % instead of 37 % for 4-h labeled cells. This discrepancy was observed in an experiment in which the two batches of Chinese hamster ovary cells (pulsed 15 min and 4 h) were labeled respectively with [3H]uridine and [14C]uridine. This enabled us to carry out the fractionation proce- dure on mixed batches and thus to obtain more significant results.
The analysis of RNA recovered in Pi, and S;, is shown in Fig.3 for the two incorporation times studied. It would then appear that Pi5 profiles roughly correspond to a rather higher molecular-weight dis- tribution than S;, ; this result may be correlated to the preferential recovcry of newly synthesized RNA in Pis as compared to long-labeled RNA species.
Ultrastructural Characterization of Pi5 Elements
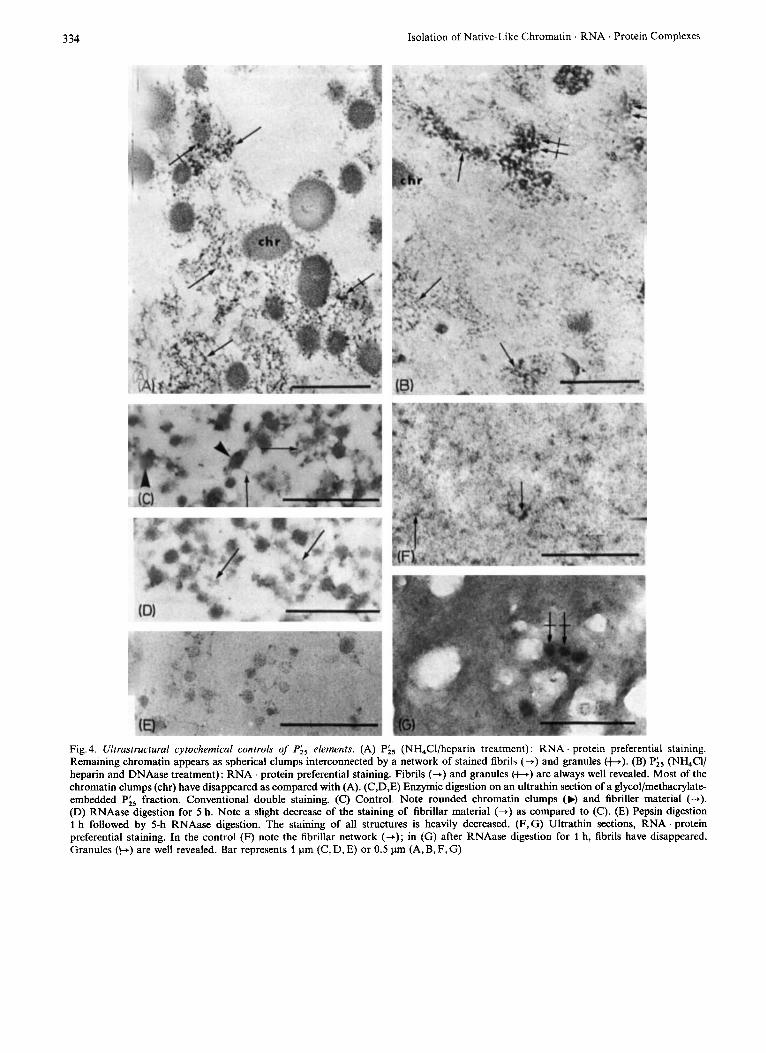
The remaining chromatin in Pi5 was detected in the form of spherical “amorphous” clumps inter- connected by a network of fibrils of about 6nm in diameter. These fibrils appear closely linked to the rounded “amorphous” structures of chromatin and are sometimes located at their borderline. Among the fibrillar network, granules of about 40 nm in diameter can be observed; they are either isolated or grouped in small clusters. Both fibrils and granules were re- vealed by EDTA preferential staining for RNA . pro- tein, although the spherical chromatin structures are only slightly bleached (Fig. 4A). If the NH,Cl/heparin treatment is carried out in the presence of DNase I (100 pg/ml), the characteristic spherical structures disappear almost completely. RNA . protein fibrils and granules are quite well preserved and remain visible after EDTA preferential staining (Fig. 4 B). These morphological features were found to be similar For the two cell species studied, Chinese hamster ovary and ascitic hepatoma cells. Moreover, when applied
to PI,,,, fraction, NH4Cl heparin treatment results in almost identical morphological patterns for Pioo elements, as compared to Pi5.
Ultrastructural Cytochemical Controls
When an RNase digestion was performed on a glycolmethacrylate-embedded Fraction the RNAase effect was slight, even after a 5-h action, as compared to the control (Fig.4C,D).
When RNAase digestion was performed on ultra- thin frozen sections, the effect was more important: after preferential RNA . protein staining, the complete disappearance of fibrils was obtained. On the other hand, the contrast of granules remained unchanged (Fig. 4F,G).
The double digestion, pepsin + RNAase, could not be carried out on ultrathin frozen sections, due to their extreme sensibility to proteolytic action. On glycolmethacrylate-embedded material, the staining contrast of all structures was heavily decreased (Fig. 4 E).
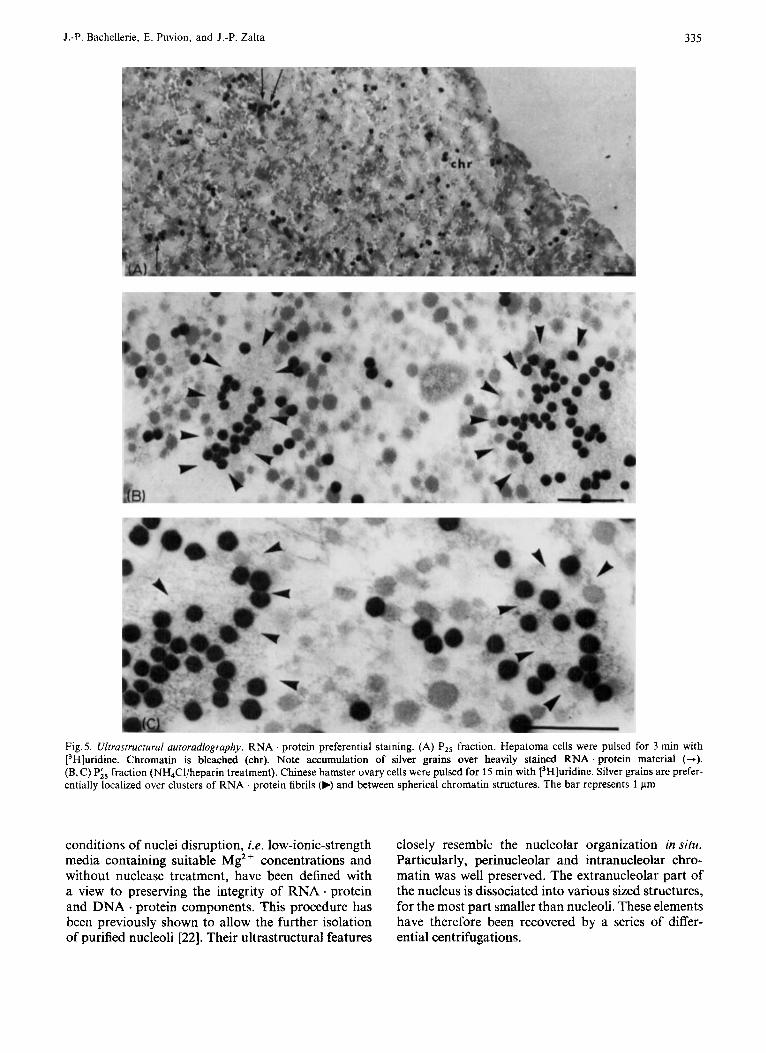
Ultrastructural Autoradiography In a first experiment on ascitic hepatoma cells,
after a 3-min pulse with [3H]uridine, cells were frac- tionated and extranucleolar nuclear chromatin . RNA protein (P2, and Ploo) was processed for autoradio-
graphic observation as described in Materials and Methods. After prolonged exposure significant label- ing in both fractions was revealed.
Although the resolving power is not sufficient to assume that silver grains are precisely located on RNA . protein particles, accumulations of silver grains upon densely packed and heavily stained RNA pro- tein structures could clearly be detected (Fig. 5A).
In a second series of experiments, Chinese hamster ovary cells were pulsed-labeled for 15 min with [3H]- uridine. After a shorter exposure time for autorddio- graphs, extranucleolar nuclear elements were well revealed the labeling pattern being qualitatively similar to the preceding experiment. These elements re- covered after NH4Cl/heparin treatment, were also observed in the conditions described above. In this case, a preferential labeling of the clusters of RNA fibrils and granules clearly appears. It is important to observe that between the spherical “amorphous” chromatin structures, the silver grains appear pre- dominantly located on the RNA . protein fibrillar component which presents similar characteristics to the so-called perichromatin fibrils in situ (Fig. 5 B, C).
DISCUSSION
In the present work, the dispersion of nuclear components was achieved through the sonication of a purified nuclear suspension. Our physicochemical
334 Isolation of Native-Like Chromatin RNA . Protein Complexes
Fig. 4. Ultrasrructural cyrochemical controls of Pis elements. (A) Pis (NH,Cl/heparin treatment) : RNA protein preferential staining. Remaining chromatin appears as spherical clumps interconnected by a network of stained fibrils (+) and granules (+). (B) Pi5 (NH,Cl/ heparin and DNAase treatment): RNA . protein preferential staining. Fibrils (-+) and granules (t.) are always well revealed. Most of the chromatin clumps (chr) have disappeared as compared with (A). (C,D,E) Enzymic digestion on an ultrathin section of a glycol/methacrylate- embedded Pi, fraction. Conventional double staining. (C) Control. Note rounded chromatin clumps (N and fibriller material (+). (D) RNAase digestion for 5 h. Note a slight decrease of the staining of fibrillar material (+) as compared to (C). (E) Pepsin digestion 1 h followed by 5-h RNAase digestion. The staining of all structures is heavily decreased. (F,G) Ultrathin sections, RNA . protein preferential staining. In the control (F) note the fibrillar network (+); in (G) after RNAase digestion for 1 h, fibrils have disappeared. Granules (b) are well revealed. Bar represents 1 pm (C, D, E) or 0.5 pm (A, B, F, G)
J.-P. Bachellerie, E. Puvion, and J.-P. Zalta 335
Fig. 5 . Ultrustructurul uutorodiogruphy. RNA . protein preferential staining. (A) PZ5 fraction. Hepatoma cells were pulsed for 3 min with [3H]uridine. Chromatin is bleached (chr). Note accumulation of silver grains over heavily stained RNA . protein material (+). (B, C) Pi5 fraction (NH,Cl/heparin treatment). Chinese hamster ovary cells were pulsed for 15 min with [3H]uridine. Silver grains are prefer- entially localized over clusters of RNA . protein fibrils (b) and between spherical chromatin structures. The bar represents 1 pm
conditions of nuclei disruption, i.e. low-ionic-strength media containing suitable Mg2 + concentrations and without nuclease treatment, have been defined with a view to preserving the integrity of RNA . protein and DNA . protein components. This procedure has been previously shown to allow the further isolation of purified nucleoli [22]. Their ultrastructural features
closely resemble the nucleolar organization in situ. Particularly, perinucleolar and intranucleolar chro- matin was well preserved. The extranucleolar part of the nucleus is dissociated into various sized structures, for the most part smaller than nucleoli. These elements have therefore been recovered by a series of differ- ential centrifugations.

336 Isolation of Native-Like Chromatin . RNA . Protein Complexes
PZ5 fraction contains the major part of extra- nucleolar nuclear RNA and DNA. Furthermore its RNA component presents some characteristic proper- ties of hnRNA described elsewhere: it presents a heterogeneous high-molecular-weight distribution as well was the early labeling of the largest molecular species, and is distinct from rRNAs (according to chain length distribution or response to actinomycin D treatment).
The morphological study of both extranucleolar fractions have shown the presence of definite RNA . protein . chromatin areas resembling chardcter- istic patterns of spatial organization in situ.
As for RNA . protein complexes, two species were detected, respectively of fibrillar and granular type.
Fibrillar RNA ’ Protein Particles
From both cytochemical properties and topo- graphical distribution, the characteristics of fibrillar RNA . protein particles are analogous with those of the so-called “perichromatin fibrils” previously ob- served in situ by Monneron and Bernhard [lo]. They appear to be more sensitive to RNAase digestion then the granular component, and disappear almost completely after a combined pepsin/RNAase action. Moreover, the fibrillar component is always revealed in close association with chromatin, lying as a small rim at the periphery of the chromatin areas in native P,, elements or closely bound to the spherical amor- phous clumps of chromatin after NH,Cl heparin treatment. The average diameter of these fibrillar elements ( 5 - 10 nm) is about the same as the values determined in situ for perichromatin fibrils. Up till now only indirect evidence indicated that perichroma- tin fibrils could represent the morphological state of hnRNA: after very short pulses with [3H]uridine and autoradiographic observations, Fakan and Bernhard [ 181 were indeed able to observe a preferential labeling over the regions where perichromatin fibrils are known to be located, that is at the border line be- tween condensed and EDTA-bleached chromatin. Furthermore, Petrov and Bernhard [33] were able to induce the appearance of perichromatin fibrils in hepatocyte of starved rats some 30 min after cortisone injection, a treatment known to enhance RNA syn- thesis. Conversely, the blocking of extranucleolar RNA synthesis by a-amanitin is correlated to a striking decrease in the amount of perichromatin fibrils in the nucleus of rat liver cells [34]. While it was not possible to demonstrate in situ a specific labeling of peri- chromatin fibrils for lack of resolving power, we were able to show unequivocally this predominant labeling of the fibrillar areas on the isolated extranucleolar elements after NHLCl heparin treatment. This is a direct evidence of the rapid labeling of the fibrillar
component which presents identical characteristics with the so-called perichromatin fibrils in situ.
Granular RNA . Protein Particles
Granules, about 40- 50 nm in diameter, were detected in both extranucleolar fractions, inside or around the bleached chromatin clumps. Small clusters of these granules can be detected after NH,Cl/heparin treatment among the fibrillar areas. These granules are found to be resistant to RNAase digestion, even in ultrathin frozen sections. These characteristics are similar to those of perichromatin granules in situ, not only demonstrated [lo] at the periphery of the con- densed chromatin but also inside, after bleaching by EDTA treatment. From indirect evidence it was sug- gested that perichromatin granules could be involved in the transfer of genetic information from nucleus to cytoplasm, possibly corresponding to be “RNA- informofers” described by Georgiev’s group [ 121. They have indeed similar ultrastructural and cyto- chemical characteristics as the Balbiani granules of giant chromosomes of Dipterus [35], which were seen crossing the nuclear envelope [36]. Moreover, they increase in number when nuclear extranucleolar RNA cannot normally be transferred to the cytoplasm as observed in rat liver cells treated by aflatoxin, lasio- carpin or low doses of actinomycin D [37]. Con- sidering the biochemical results of RNA analysis, the detection after rather prolonged labeling periods by [3H]uridine, of discrete RNA species of ribosomal or preribosomal types over the heterodisperse profile of high-molecular-weight RNAs has to be discussed. A first possibility is the presence of nucleolar frag- ments, and consequently nucleolar RNAs in the so- called extranucleolar fractions. However, the total labeling found in these ribosomal-like RNAs can hardly be attributed to such a contamination, even in the case where the actual amount of nucleolar debris present in Pzs reaches the upper limit which we have experimentally determined (see results of Table 2).
The possibility of a leakage in vitro, in the course of the fractionation, of some nucleolar RNA-con- taining elements and further recovery in PZ5 and PI,, fractions is indeed difficult to exclude. In this hypo- thesis this leakage would be the greater for more mature RNA species. Pz5 RNA patterns after short pulse labelings do not contain discrete 45-S species and are not modified when cells are pretreated with low doses of actinomycin D. Furthermore, for inter- mediate pulse periods, no clear 28-S RNA peak can be detected in extranucleolar fractions, whereas nu- cleolar 2 8 3 RNA is already significantly labeled. Consequently, the presence of ribosomal-like RNA species in extranucleolar fractions may have a physio- logical significance and could correspond to the newly-

J.-P. Bachellcrie, E. Puvion, and J.-P. Zalta 331
formed preribosomes flowing from the nucleolus to cytoplasmic areas. It is noteworthy that the major part of these “extranucleolar” ribosomal-like RNA species are bound to the rapidly sedimenting chro- matin . RNA * protein elements (P2& Under these conditions, free ribosomes should not pellet; they should be recovered almost exclusively in slower sedimenting fractions. If this linkage with elements containing newly synthesized hnRNA as a physio- logical significance, the result could be put together with the hypothesis of Harris [38] that the nucleolus may play a part in the control of the processing of mRNA precursors and the transport of mRNA sequences to the cytoplasm, through the linkage of newly formed ribosomes flowing onto nascent hnRNA chains. It is interesting to note that these ribosomal- like RNAs seem to be preferentially removed from RNA . protein network isolated after NH,Cl/heparin treatment. This problem of the presence of rRNAs species is now being examined.
The biochemical characterization of the minor DNA fraction which remains linked to the fibrillar RNA . protein network recovered after preferential chromatin dispersion, as well as the analysis of this network is now under investigation.
We gratefully acknowledge the constant support of Dr W. Bern- hard (Director of the Laboratoire de Microscopie Electronique de I’Institut de Recherches sur le Cancer Villejuif) who favoured the development of this ultrastructural study in his laboratory and provided us with much valuable advice throughout this work. The skilful technical assistance of Mrs M. J. Burglen and M. Nicoloso is greatly appreciated. These investigations were aided by a contract from I.N.S.E.R.M. (no. 74.1.226.2).
REFERENCES
1. Scherrer, K., Latham, H. & Darnell, J. E. (1963) Proc. Nut1
2. Gcorgiev, G. P., Samarina, 0. P., Lerman, M. L. & Smirnov, M. N. & Severtzov, A. N. (1963) Nuture (Land.) 200. 1291 - 1294.
3. Scherrer, K. & Marcaud, L. (1965) Bull. Soc. Chim. Biol. 47,
4. Scherrer, K . , Marcaud, L., Zajdela, F., London, I. M. & Gros, F. (1966) Proc. Natl Acad. Sci. U.S.A. 56, 1571- 1578.
5. Warner, J. R., Soeiro, R., Birnboim, H. C., Girard, M. & Dar- ncll, J. E. (1966) J. Mol. Biol. 19, 349-361.
Acud. Sci. U.S.A. 49, 240-248.
1967- 1973.
6.
7.
8.
9.
10.
11.
12.
13. 14. 15. 16.
17.
18. 19. 20. 21. 22.
23.
24.
2s.
26.
21. 28.
29. 30. 31.
32.
33.
34.
3s.
36. 37.
38.
Imaizumi, T., Diggelmann, H. & Schemer, K . (1973) Proc.
Darnell, J. E., Philipson, L., Wall, R. & Adesnik, M. (1971)
Perry, R. P., Kclley, D. E. & Latorre, J. (1974) J . Mol. Biol.
Price, R. P., Ransom, L. & Penman, S. (1974) Cell, 2, 253-
Monncron, A. & Bernhard, W. (1969) .I. tiltra.struct. Res, 27,
Miller, 0. L. & Bakken, A. (1972) Fifth Kurolinska Symposium on Research Methods in Reproductive Endocrinology: Gene Transcription in Reproductive Tissues, pp. 155 - 177. Bog- trykkerit Forum, Copenhagen.
Samarina, 0. P., Lukanidin, E. M., Molnar, J. & Georgicv, G . P. (1968) J . Mol. B i d . 33, 251-263.
Stevenin, J . &Jacob, M. (1972) Eur. J . Biochem. 29,480-488. Stevenin, J. & Jacob, M. (1974) Eur. J. Biochem. 47, 129- 137. Pcdcrson, T. (1974) J . Mol. Biol. 83, 163-183. Bernhard, W. & Granboulan, N. (1963) Exp. Cell Res. Supp.
Granboulan, M. & Bernhard, W. (1961) Slances Soc. Biol. Fil.
Fakdn, S. & Bernhard, W. (1971) E x p . Cell Rex. 67, 129- 141. Fischer, G. A. (1958) Ann. N.Y. Acad. Sci. 76, 673-680. Eagle, H. (1959) Science (Wash. O.C.) 130,432-437. Earlc, W. R. (1943) J. Nut. Cancer Inst. 4, 165-212. Zalta, J., Zalta, J. P. & Simdrd, R. (1971) J . Cell Bid . 51,
Nakamura, T., Rapp, F. & Busch, H. (1967) Cancer Rex 77,
Perry, R. P., Latorre, J . , Kelley, D. E. & Greenberg, J . R.
Tiollais, P., Galibert, F., Lepctit, A. & Augcr, M. A. (1972)
Leduc, E. H. & Bernhard, W. (1967) J . tiltrastruct. Res. 19,
Bcrnhard, W. (1969) J . Ultrastruct. Res. 27, 250-265. Benichou, J. C. & Ryter, A. (1973) J . Microsc. (Paris) 17,
Bemhard, W. & Viron, A. (1971) J . Cell Biol. 49, 731 - 746. Puvion, E. & Bernhard, W. (1975) J. Cell Biol. in press. Haase, G . & Jung, G. (1964) Naturwissenschaften, 51, 404-
Perry, R. P. & Kelley, D. E. (1968) J . Cell. Physiol. 72, 235
Petrov, P. & Bernhard, W. (1971) J . tiltrastruct. Rc.s. 35,
Petrov, P. & Sekeris, C. E. (1971) E x p . Cell. Res. 69, 393-
Vasquez-Nin, G. & Bernhard, W. (1971) J . Ultrastruct. Res.
Stevens, B. J . & Swift, H. (1966) J . Cell Bid. 31, 55-77. Monneron, A. (1968) Hebd. Siances Acad. Sci. Sir . D . Sci.
Harris, H. (1974) Nucleus and Cytoplasm, p. 138, Clarendon
NatlAcad. Sci. U.S.A. 70, 1122-1126.
Science (Wash. D.C.) 174, 507-510.
82,315-331.
258.
266 - 288.
9, 19-53.
155, 1767 - 1719.
563 - 568.
1084- 1091.
(1972) Biochim. Biophy.7. Actu, 262, 220 - 226.
Biochimie (Paris) 54 , 339- 354.
196-199.
221 - 232.
405.
- 246.
386-402.
401.
36,842- 860.
Nut. (Paris) 267,2053-2056.
Press, Oxford.
J.-P. Bachellerie and J.-P. Zalta, Centre de Recherche de Biochimie et de Genetique Cellulairesdu C.N.R.S., 118 Route de Narbonne, F-31077 Toulouse-Cedex, France
E. Puvion, Laboratoire de Microscopie Electronique du C.N.R.S., Institut de Recherches Scientifiques sur le Cancer, Boite postale 8, F-94800 Villejuif, France

















