Embed Size (px)
Citation preview

Using human umbilical cord cells for tissue engineering: A comparisonwith skin cells
Cindy J. Hayward a,b,c, Julie Fradette a,b,c, Pascal Morissette Martin a,b,c, Rina Guignard a,b,Lucie Germain a,b,c, François A. Auger a,b,c,n
a Centre de recherche en organogénèse expérimentale de l’Université Laval/LOEX, Aile-R, Hôpital de l0Enfant-Jésus, Centre de recherche du CHU de Québec,1401, 18e Rue, Québec, QC, Canada G1J 1Z4b Axe Médecine Régénératrice—Centre de recherche FRQS du CHU de Québec, Québec, QC, Canadac Département de Chirurgie, Faculté de Médecine, Université Laval, Québec, QC, Canada
a r t i c l e i n f o
Article history:Received 20 February 2014Accepted 15 May 2014Available online 11 June 2014
Keywords:Umbilical cordTissue engineeringEpithelial differentiationTissue interactionsEpithelial cellsWharton's jelly cells
a b s t r a c t
The epithelial cells and Wharton's jelly cells (WJC) from the human umbilical cord have yet to beextensively studied in respect to their capacity to generate tissue-engineered substitutes for clinicalapplications. Our reconstruction strategy, based on the self-assembly approach of tissue engineering,allows the production of various types of living human tissues such as skin and cornea from a wide rangeof cell types originating from post-natal tissue sources. Here we placed epithelial cells and WJC from theumbilical cord in the context of a reconstructed skin substitute in combination with skin keratinocytesand fibroblasts. We compared the ability of the epithelial cells from both sources to generate a stratified,differentiated skin-like epithelium upon exposure to air when cultured on the two stromal cell types.Conversely, the ability of the WJC to behave as dermal fibroblasts, producing extracellular matrix andsupporting the formation of a differentiated epithelium for both types of epithelial cells, was alsoinvestigated. Of the four types of constructs produced, the combination of WJC and keratinocytes wasthe most similar to skin engineered from dermal fibroblasts and keratinocytes. When cultured on dermalfibroblasts, the cord epithelial cells were able to differentiate in vitro into a stratified multilayeredepithelium expressing molecules characteristic of keratinocyte differentiation after exposure to air, andmaintaining the expression of keratins K18 and K19, typical of the umbilical cord epithelium. WJC wereable to support the growth and differentiation of keratinocytes, especially at the early stages of air–liquidculture. In contrast, cord epithelial cells cultured on WJC did not form a differentiated epidermis whenexposed to air. These results support the premise that the tissue from which cells originate can largelyaffect the properties and homoeostasis of reconstructed substitutes featuring both epithelial and stromalcompartments.
& 2014 International Society of Differentiation. Published by Elsevier B.V. All rights reserved.
1. Introduction
The development of reconstructed living substitutes has greatlyprogressed in recent years, in particular in the field of skinreconstruction, which is among the most advanced for tissueengineering applications. Achieving tissue homoeostasis withinreconstructed substitutes is especially important for self-renewingtissues such as skin, which features a differentiated epithelialcompartment influenced by the underlying dermal compartment(McLoughlin, 1963). Tissue-engineered substitutes generated usingthe self-assembly method are useful tools for gaining insights intocell interactions and the regulation of tissue homoeostasis. Indeed,being by nature made of cells and the native extracellular matrixthey secrete and organise upon stimulation with ascorbic acid,these living tissues recreate a highly physiological tissue context
Contents lists available at ScienceDirect
journal homepage: www.elsevier.com/locate/diff
Differentiation
http://dx.doi.org/10.1016/j.diff.2014.05.001Join the International Society for Differentiation (www.isdifferentiation.org)0301-4681/& 2014 International Society of Differentiation. Published by Elsevier B.V. All rights reserved.
Abbreviations: DF, Dermal fibroblasts; DMEM, Dulbecco–Vogt modified Eagle'smedium; EDTA, Ethylenediaminetetraacetic acid; EGF, Epidermal growth factor;K, Keratinocytes; K10, Keratin 10; NCS, Newborn calf serum; P, Passage; SEM,Standard error of the mean; TRITC, Tetramethylrhodamine-5-(and-6)-isothiocya-nate; UC, Umbilical cord; ucEpi, Umbilical cord epithelial cells; WJC, Wharton's jellycells
n Corresponding author at: LOEX, Aile-R, Hôpital de l0Enfant-Jésus, Centre derecherche du CHU de Québec, 1401, 18e Rue, Québec, QC, G1J 1Z4.Tel.: þ1 418 990 8255x1662; fax: þ1 418 990 8248.
E-mail addresses: [email protected] (C.J. Hayward),[email protected] (J. Fradette),[email protected] (P. Morissette Martin),[email protected] (R. Guignard),[email protected] (L. Germain),[email protected] (F.A. Auger).
URL: http://www.loex.qc.ca (F.A. Auger).
Differentiation 87 (2014) 172–181

(Paquet et al., 2010). Such substitutes are particularly helpful forunderstanding the contribution of stromal–epithelial interactions todifferentiation patterns and regulation (Trottier et al., 2008; Carrieret al., 2009) in addition to being employed in the treatment of skindefects (Boa et al., 2013).
Umbilical cord-derived cells have spurred increasing interest inrecent years because of their advantageous characteristics, such asa less differentiated status possibly contributing to lower immu-nogenicity (Ennis et al., 2008; Weiss et al., 2008). In addition tocord blood stem cells, cells derived from the umbilical cord solidtissue are of great interest. Of the four cell types that can besuccessfully extracted and cultured from a single umbilical cord(Hayward et al., 2013), endothelial cells and smooth muscle cells ofthe vein and arteries have been extensively used in variousexperimental settings (Jaffe et al., 1973; Owens, 1995; Bachettiand Morbidelli, 2000), including tissue engineering (L’Heureuxet al., 1998; Stephan et al., 2006; Gibot et al., 2010; Guillemetteet al., 2010; Rochon et al., 2010; Tsigkou et al., 2010 Gauvin et al.,2011). The Wharton's jelly cells (WJC) have also become thesubject of intensive research in recent years (reviewed in Marcusand Woodbury (2008), and in Troyer and Weis, (2008)) with anever-increasing number of studies demonstrating their character-istics as multipotent stem cells. The epithelial cells of the umbilicalcord show an interesting pattern of keratin expression, combiningcharacteristics of simple and stratified epithelia (Mizoguchi et al.,2000; Hayward et al., 2013). Recent studies have highlighted theirsimilarity to neonatal keratinocytes in the pattern of K1/K10 anddeltaNp63 expression (Ruetze et al., 2008). They are also able toform a stratified epithelium when seeded on collagen gels popu-lated with fibroblasts (Mizoguchi et al., 2004; Sanmano et al.,2005; Ng et al., 2008; Huang et al., 2011). In this study, weevaluated their behaviour in combination with WJC or dermalfibroblasts in the context of a tissue-engineered skin substitute.
The aim of the present study was to evaluate the impact ofepithelial and stromal cells from two different cell sources on thetissue production of bilayered substitutes by the self-assemblyapproach of tissue engineering. Our results show that the differentcombinations of skin- and umbilical cord-derived cells had a greatimpact on resulting differentiation pattern of the epithelial com-partment as well as on the development of a mature dermo-epidermal junction. Umbilical cord epithelial cells cultured ondermal fibroblasts differentiated in a similar fashion to keratino-cytes when exposed to air, showing the presence of molecularmarkers characteristic of stratified, keratinised epithelia. Kerati-nocytes on WJC also differentiated in a similar fashion, butumbilical cord epithelial cells on WJC did not produce a multi-stratified, skin-like epithelium in air-exposed culture conditions.Thus, not all cell sources are interchangeable, and the particularcombination of cells can have as much impact on the resultingtissue as culture conditions.
2. Materials and methods
2.1. Cell extraction and culture
Human umbilical cords (N¼6 different donors) were obtainedfrom the obstetrics unit of the Saint-Sacrement Hospital after
normal births, with informed consent from the mothers. Allprotocols were approved by the Research Ethical Committee ofthe Centre hospitalier affilié universitaire de Québec (CHA). Kera-tinocytes were obtained from newborn foreskin as previouslydescribed (Germain et al., 1993), and epithelial cells were extractedfrom the human umbilical cord using a solution of 0.25% trypsin(ICN) with 2.2 mM EDTA in buffered saline solution, as described inHayward et al. (2013). For passaging, these cells were seeded in75 cm2 culture flasks at 1�106 cells per flask with a feeder layer ofirradiated Swiss 3T3 cells (2�104 cells/cm2; Germain et al., 1993)and cultured as described below. Dermal fibroblasts were obtainedfrom an adult skin biopsy (23-year-old female donor) as in a priordescription (Auger et al., 1995). Umbilical cord WJC were isolated asdescribed in Hayward et al. (2013). Both types of stromal cells wereseeded in 75 cm2 culture flasks at 1�104 cells/cm2 and cultured asdescribed below. All cell populations were cryopreserved in liquidnitrogen pending their use in the experiments, then thawed andcultured for at least one passage. All cell cultures and reconstructedtissues were maintained at 37 1C in a humidified atmospherecontaining 8% CO2, and media were changed three times per week.
Keratinocytes and epithelial cells were cultured in Dulbecco–Vogtmodified Eagle's medium (DMEM) combined with Ham's F12 medium(both Gibco BRL, Burlington, Canada) in a 3:1 proportion (referred toas DME–Ham's), supplemented with 10% newborn calf serum (NCS;Fetal Clone II, HyClone, Aurora, Canada), epithelial additives [24.3 μg/mL adenine (Chiron Corp., Emeryville, U.S.A.), 10 ng/mL epidermalgrowth factor (EGF; Austral Biological, San Ramon, U.S.A.), 5 μg/mLinsulin (Sigma Chemical Co., St. Louis, U.S.A.), 0.1 nM cholera toxin (ICNBiomedical, Montréal, Canada), 5 μg/mL transferrin (Boehringer Man-nheim, Laval, Canada), 2 nM 3,30,5 triiodo-L-thyronine (Sigma), 0.4 μg/mL hydrocortisone (Calbiochem, La Jolla, U.S.A.)], and antibiotics[100 IU/mL penicillin G (Sigma), 25 μg/mL gentamicin sulphate (Scher-ing, Pointe-Claire, Canada)]. Fibroblasts and WJC were cultured inDMEM (Gibco BRL), supplemented with 10% foetal calf serum(HyClone) and antibiotics.
2.2. Reconstructed bilayered stromal/epithelial substitutes
Four different reconstructed substitutes were produced in thisstudy, using the following combinations (Table 1): dermal fibro-blasts with skin keratinocytes (DF/K), dermal fibroblasts with cordepithelial cells (DF/ucEpi), WJC with keratinocytes (WJC/K) andWJC with cord epithelial cells (WJC/ucEpi). For each combinationof specific cell populations, up to seven substitutes were prepared,which allowed sampling of two substitutes for final analysis ateach time point. A total of six umbilical cord cell populations(three epithelial and three stromal) were used in five differentepithelial cell–WJC combinations, along with one dermal fibro-blast cell population and one keratinocyte cell population forcomparison purposes.
Bilayered stromal/epithelial substitutes comprising both anepidermal and a stromal layer were prepared using the self-assembly approach of tissue engineering as previously described(Michel et al., 1999). Briefly, dermal fibroblasts or WJC between P3and P7 were seeded in a 25 cm2 culture flasks at a density of2�105 cells per flask with 50 mg/mL sodium L-ascorbate (Sigma) inthe culture medium, which promotes the deposition of extracel-lular matrix material. Fresh ascorbic acid was added at each
Table 1Composition of the bilayered stromal/epithelial substitutes.
DF/K DF/ucEpi WJC/K WJC/ucEpi
Keratinocytes (K) UC epithelial cells (ucEpi) Keratinocytes (K) UC epithelial cells (ucEpi)Dermal fibroblasts (DF) Wharton's Jelly Cells (WJC)
C.J. Hayward et al. / Differentiation 87 (2014) 172–181 173

change of culture medium. Manipulable sheets were obtainedafter a maturation period of at least 14 days for WJC and 28 daysfor dermal fibroblasts. After being gently peeled off the cultureflask surface with forceps, two sheets were superposed in a culturePetri dish and cultured for an additional week in the sameconditions to ensure cohesion of this stromal compartment. Atthis time, keratinocytes or umbilical cord epithelial cells (ucEpi)were dissociated with trypsin-EDTA and seeded on the surface ofthe stromal compartment at a density of 2�105 cells/cm2. Thesubstitutes were cultured in submerged conditions for seven dayswith ascorbic acid still being added at every change of medium.Submerged bilayered stromal/epithelial substitutes were culturedin DME–Ham's, supplemented with 5% NCS, epithelial additivesand antibiotics. Two substitutes per combination were processedfor histological and labelling analyses at this time point, while theothers were further cultured as follows.
The reconstructed bilayered stromal/epithelial substitutes werethen placed on a plastic support allowing the stromal compart-ment to remain in contact with the culture medium while theepithelial compartment was exposed to air. Culture at the air–liquid interface is recognised to induce highly physiological skinepithelial differentiation (Pruniéras et al., 1983; Font et al., 1994;Larouche et al., 2009). Bilayered stromal/epithelial substitutes atthe air–liquid interface were cultured in the same mediumwithout EGF.
2.3. Population doubling calculations
The number of population doublings obtained at the end ofeach subculture was calculated with the formula x¼3.32�(log N� log N0), where N is the number of cells obtained at theend of the subculture, and N0 is the number of viable cellsoriginally seeded (viability being determined by the exclusion oftrypan blue dye). Average doubling times were calculated bydividing the number of hours of subculture by the populationdoublings attained during this time. Data is represented as mean7SEM. For statistical comparisons, a one-way ANOVA was per-formed with Dunnett's post-test, using GraphPad Prism4 software(GraphPad Software, Inc., San Diego, U.S.A.). The confidence inter-val was set at 95% (po0.05).
2.4. Histological analysis and immunofluorescence labelling
Samples from two different constructs of each combinationwere taken on days 0 (immerged), 7 and 21 of culture at the air–liquid interface. A portion of each sample was fixed in Bouin'ssolution (ACP, St. Léonard, Canada) and embedded in paraffin.Cross-sections were mounted on slides and coloured with Mas-son's trichrome staining. Photographs were taken by an AxioCamICc1 camera mounted on a Zeiss Imager M2. The thickness of theepithelial and stromal compartments was determined on histolo-gical sections using the ImageJ software (NIH) for all combinationsat days 0, 7 and 21, using a minimum of 18 measurements perconstruct for each experimental condition. Data is represented asmean 7SEM. Significance was determined after a one-wayANOVA followed by a Tukey post-test.
Other sample portions were quick-frozen in OCT compound(Tissue-Tek, U.S.A.) and conserved at �80 1C until they were cut bya microtome (JUNG RM2035, Leica) into 5 μm cross-sectionswhich were placed on glass slides for immunofluorescence stain-ing. After fixation in acetone, indirect immunofluorescence assayswere performed as previously described (Germain et al., 1988) onthe reconstructed tissue cross-sections as well as on sections ofwhole umbilical cord, cornea and normal human skin as referencetissues. Cell nuclei were subsequently labelled with Hoechstreagent (50 mg/mL, used at 1:100; Sigma) and the samples
examined under a Nikon Eclipse microscope and photographedwith Kodak Tmax 400 ASA film or a Photometrics Sensys CCDcamera. Film images with excessive background were adjustedusing the gamma function in Photoshop.
The antibodies used include mouse monoclonal antibodiesagainst human α-smooth muscle actin (DAKO); involucrin, keratins(K) 10 and 14 (all Sigma); type III and VII collagens and laminin5 subunit α2 (Chemicon); filaggrin and transglutaminase (BTI); K12and vimentin (ICN); K18 and K19 (ARP); rabbit antibodies againsttype I collagen (Biodesign); type IV collagen (Chemicon); and K16(gift of Dr. P. Coulombe – Bernot et al., 2002). Secondary antibodieswere TRITC-coupled goat anti-mouse (Chemicon) and TRITC-coupled goat anti-rabbit (Chemicon). Negative controls were per-formed by omission of the primary antibodies.
2.5. Electron microscopy
Specimens were prepared for electron microscopy by fixationin 2.5% glutaraldehyde. They were then washed with 0.2 Mcacodylate buffer, postfixed with 2% OsO4 for 30 min, andembedded in LR White. Thin sections stained with uranyl acetateand lead citrate were observed with a JEOL 1230 transmissionelectron microscope.
3. Results
3.1. Cell differentiation within the substitute's epithelialcompartment is influenced by cell origin
The reference substitute comprising human keratinocytesseeded on dermal fibroblast sheets formed the typical skin sub-stitute extensively characterised in our previous work (Michel etal., 1999; Trottier et al., 2008; Lavoie et al., 2013). It features a well-developed epithelium displaying all the characteristic layers indi-cative of keratinocyte differentiation (Fig. 1A–C). After seven daysof culture in submerged conditions, the keratinocytes formed oneto two layers (Fig. 1A). However, after only seven days of culture atthe air–liquid interface, the stratified epithelium consisted ofseven or more layers due to proliferation of the epithelial cells,with an abundance of desmosome-like structures visible at the celljunctions under light microscopy (Fig. 1B). The basal cells werecuboidal in shape, with cells becoming increasingly flattenednearer the upper surface. Granules were visible in the uppermostcells just under the thin layer of cornified, flat squames at thesurface. A similar structure was observed for the equivalents at day21, except that the nucleated epithelial layers were somewhatthinner, and there were many more layers of cornified cells at theupper surface of the substitutes (Fig. 1C).
Human keratinocytes seeded on WJC (WJC/K substitutes)formed an average of two epithelial layers by day seven ofsubmerged culture (Fig. 1D). Interestingly, the epithelium was fourtimes as thick on average at this time point than that of the controlsubstitute (po0.001; Fig. 1G). Once exposed to the air–liquidinterface and cultured for a week, four to five epithelial layerscould be observed (Fig. 1E). The basal cells were rounded andthose closer to the surface flattened. In general the substituteswere well developed, the epithelium tending to be thinner onaverage than that of the reference skin substitutes at this timepoint (Fig. 1G). After 21 days of exposure at the air–liquid interfacethe epidermis was thinner, with most regions showing thecharacteristics of skin-like epithelial differentiation (Fig. 1F).The epithelium at this stage averaged 43% of the thickness of thecontrol substitute (po0.01, Fig. 1G).
We next compared the proliferation and differentiation poten-tials of umbilical cord epithelial cells (ucEpi). All ucEpi populations
C.J. Hayward et al. / Differentiation 87 (2014) 172–181174
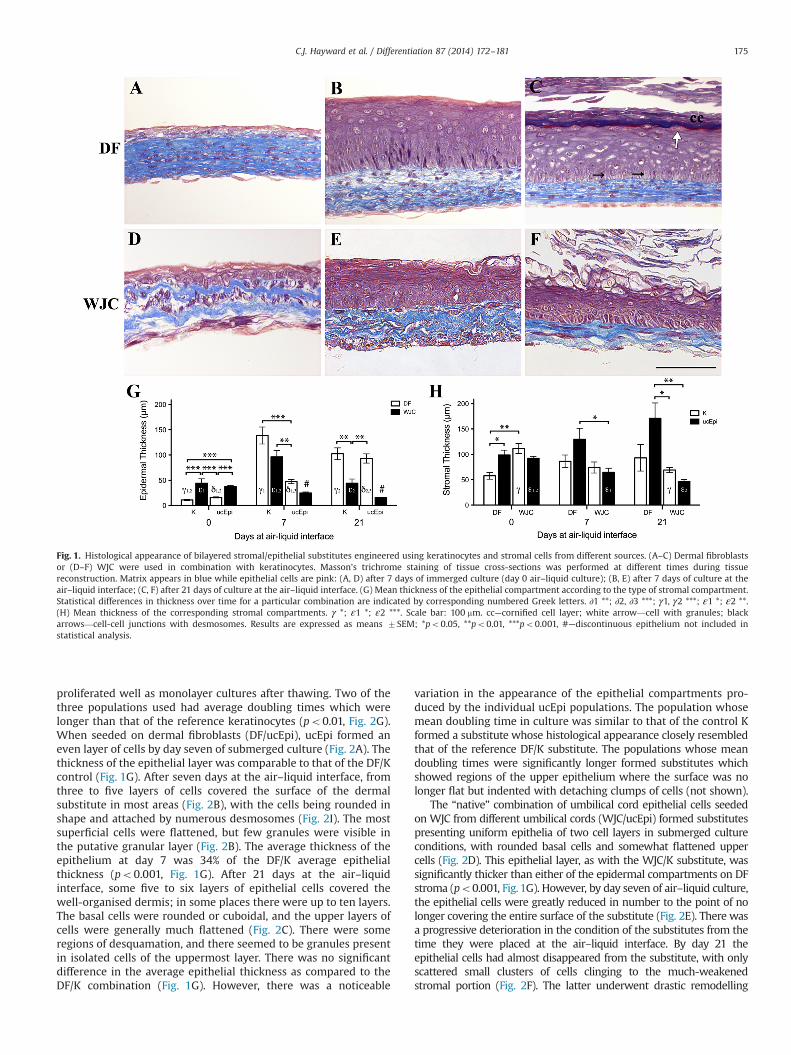
proliferated well as monolayer cultures after thawing. Two of thethree populations used had average doubling times which werelonger than that of the reference keratinocytes (po0.01, Fig. 2G).When seeded on dermal fibroblasts (DF/ucEpi), ucEpi formed aneven layer of cells by day seven of submerged culture (Fig. 2A). Thethickness of the epithelial layer was comparable to that of the DF/Kcontrol (Fig. 1G). After seven days at the air–liquid interface, fromthree to five layers of cells covered the surface of the dermalsubstitute in most areas (Fig. 2B), with the cells being rounded inshape and attached by numerous desmosomes (Fig. 2I). The mostsuperficial cells were flattened, but few granules were visible inthe putative granular layer (Fig. 2B). The average thickness of theepithelium at day 7 was 34% of the DF/K average epithelialthickness (po0.001, Fig. 1G). After 21 days at the air–liquidinterface, some five to six layers of epithelial cells covered thewell-organised dermis; in some places there were up to ten layers.The basal cells were rounded or cuboidal, and the upper layers ofcells were generally much flattened (Fig. 2C). There were someregions of desquamation, and there seemed to be granules presentin isolated cells of the uppermost layer. There was no significantdifference in the average epithelial thickness as compared to theDF/K combination (Fig. 1G). However, there was a noticeable
variation in the appearance of the epithelial compartments pro-duced by the individual ucEpi populations. The population whosemean doubling time in culture was similar to that of the control Kformed a substitute whose histological appearance closely resembledthat of the reference DF/K substitute. The populations whose meandoubling times were significantly longer formed substitutes whichshowed regions of the upper epithelium where the surface was nolonger flat but indented with detaching clumps of cells (not shown).
The “native” combination of umbilical cord epithelial cells seededon WJC from different umbilical cords (WJC/ucEpi) formed substitutespresenting uniform epithelia of two cell layers in submerged cultureconditions, with rounded basal cells and somewhat flattened uppercells (Fig. 2D). This epithelial layer, as with the WJC/K substitute, wassignificantly thicker than either of the epidermal compartments on DFstroma (po0.001, Fig. 1G). However, by day seven of air–liquid culture,the epithelial cells were greatly reduced in number to the point of nolonger covering the entire surface of the substitute (Fig. 2E). There wasa progressive deterioration in the condition of the substitutes from thetime they were placed at the air–liquid interface. By day 21 theepithelial cells had almost disappeared from the substitute, with onlyscattered small clusters of cells clinging to the much-weakenedstromal portion (Fig. 2F). The latter underwent drastic remodelling
Fig. 1. Histological appearance of bilayered stromal/epithelial substitutes engineered using keratinocytes and stromal cells from different sources. (A–C) Dermal fibroblastsor (D–F) WJC were used in combination with keratinocytes. Masson's trichrome staining of tissue cross-sections was performed at different times during tissuereconstruction. Matrix appears in blue while epithelial cells are pink: (A, D) after 7 days of immerged culture (day 0 air–liquid culture); (B, E) after 7 days of culture at theair–liquid interface; (C, F) after 21 days of culture at the air–liquid interface. (G) Mean thickness of the epithelial compartment according to the type of stromal compartment.Statistical differences in thickness over time for a particular combination are indicated by corresponding numbered Greek letters. ∂1 **; ∂2, ∂3 ***; γ1, γ2 ***; ε1 *; ε2 **.(H) Mean thickness of the corresponding stromal compartments. γ *; ε1 *; ε2 ***. Scale bar: 100 mm. cc—cornified cell layer; white arrow—cell with granules; blackarrows—cell-cell junctions with desmosomes. Results are expressed as means 7SEM; *po0.05, **po0.01, ***po0.001, #—discontinuous epithelium not included instatistical analysis.
C.J. Hayward et al. / Differentiation 87 (2014) 172–181 175
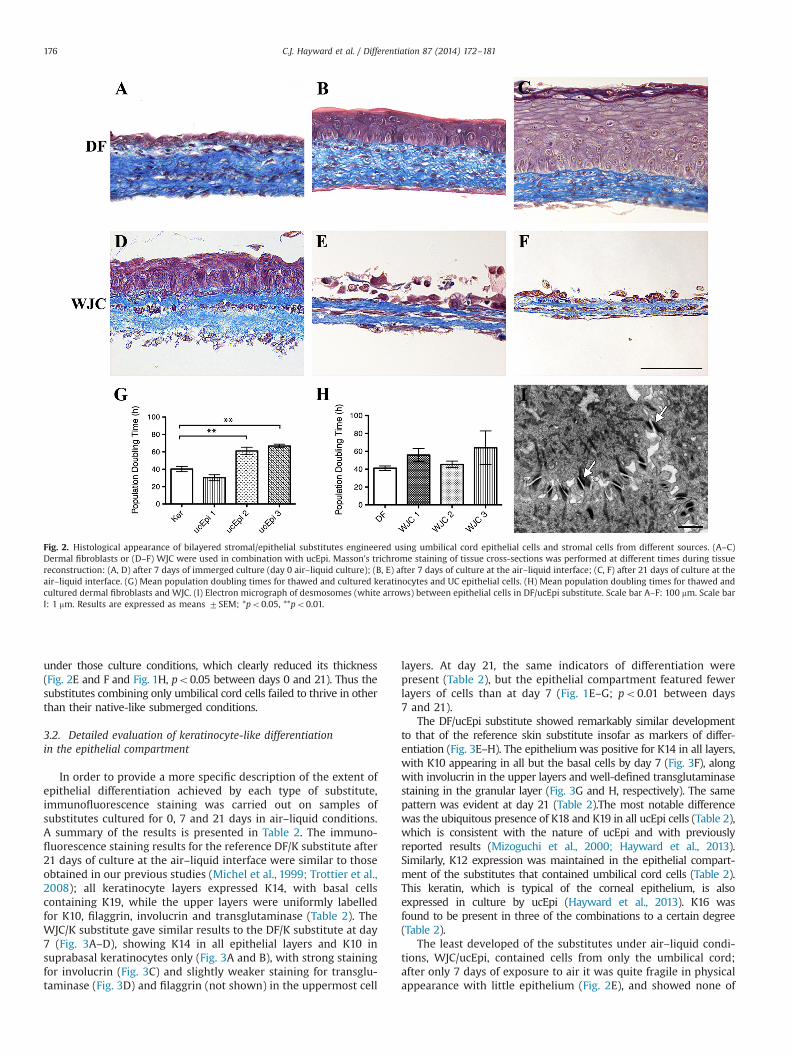
under those culture conditions, which clearly reduced its thickness(Fig. 2E and F and Fig. 1H, po0.05 between days 0 and 21). Thus thesubstitutes combining only umbilical cord cells failed to thrive in otherthan their native-like submerged conditions.
3.2. Detailed evaluation of keratinocyte-like differentiationin the epithelial compartment
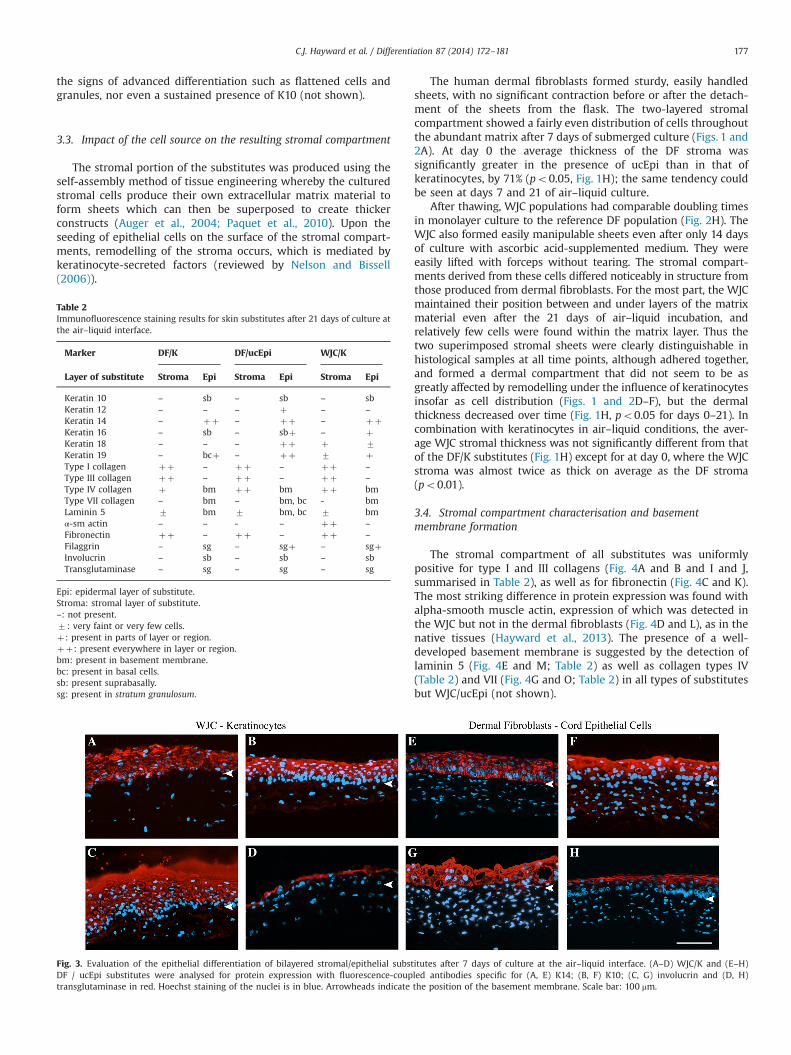
In order to provide a more specific description of the extent ofepithelial differentiation achieved by each type of substitute,immunofluorescence staining was carried out on samples ofsubstitutes cultured for 0, 7 and 21 days in air–liquid conditions.A summary of the results is presented in Table 2. The immuno-fluorescence staining results for the reference DF/K substitute after21 days of culture at the air–liquid interface were similar to thoseobtained in our previous studies (Michel et al., 1999; Trottier et al.,2008); all keratinocyte layers expressed K14, with basal cellscontaining K19, while the upper layers were uniformly labelledfor K10, filaggrin, involucrin and transglutaminase (Table 2). TheWJC/K substitute gave similar results to the DF/K substitute at day7 (Fig. 3A–D), showing K14 in all epithelial layers and K10 insuprabasal keratinocytes only (Fig. 3A and B), with strong stainingfor involucrin (Fig. 3C) and slightly weaker staining for transglu-taminase (Fig. 3D) and filaggrin (not shown) in the uppermost cell
layers. At day 21, the same indicators of differentiation werepresent (Table 2), but the epithelial compartment featured fewerlayers of cells than at day 7 (Fig. 1E–G; po0.01 between days7 and 21).
The DF/ucEpi substitute showed remarkably similar developmentto that of the reference skin substitute insofar as markers of differ-entiation (Fig. 3E–H). The epitheliumwas positive for K14 in all layers,with K10 appearing in all but the basal cells by day 7 (Fig. 3F), alongwith involucrin in the upper layers and well-defined transglutaminasestaining in the granular layer (Fig. 3G and H, respectively). The samepattern was evident at day 21 (Table 2).The most notable differencewas the ubiquitous presence of K18 and K19 in all ucEpi cells (Table 2),which is consistent with the nature of ucEpi and with previouslyreported results (Mizoguchi et al., 2000; Hayward et al., 2013).Similarly, K12 expression was maintained in the epithelial compart-ment of the substitutes that contained umbilical cord cells (Table 2).This keratin, which is typical of the corneal epithelium, is alsoexpressed in culture by ucEpi (Hayward et al., 2013). K16 wasfound to be present in three of the combinations to a certain degree(Table 2).
The least developed of the substitutes under air–liquid condi-tions, WJC/ucEpi, contained cells from only the umbilical cord;after only 7 days of exposure to air it was quite fragile in physicalappearance with little epithelium (Fig. 2E), and showed none of
Fig. 2. Histological appearance of bilayered stromal/epithelial substitutes engineered using umbilical cord epithelial cells and stromal cells from different sources. (A–C)Dermal fibroblasts or (D–F) WJC were used in combination with ucEpi. Masson's trichrome staining of tissue cross-sections was performed at different times during tissuereconstruction: (A, D) after 7 days of immerged culture (day 0 air–liquid culture); (B, E) after 7 days of culture at the air–liquid interface; (C, F) after 21 days of culture at theair–liquid interface. (G) Mean population doubling times for thawed and cultured keratinocytes and UC epithelial cells. (H) Mean population doubling times for thawed andcultured dermal fibroblasts and WJC. (I) Electron micrograph of desmosomes (white arrows) between epithelial cells in DF/ucEpi substitute. Scale bar A–F: 100 mm. Scale barI: 1 mm. Results are expressed as means 7SEM; *po0.05, **po0.01.
C.J. Hayward et al. / Differentiation 87 (2014) 172–181176
the signs of advanced differentiation such as flattened cells andgranules, nor even a sustained presence of K10 (not shown).
3.3. Impact of the cell source on the resulting stromal compartment
The stromal portion of the substitutes was produced using theself-assembly method of tissue engineering whereby the culturedstromal cells produce their own extracellular matrix material toform sheets which can then be superposed to create thickerconstructs (Auger et al., 2004; Paquet et al., 2010). Upon theseeding of epithelial cells on the surface of the stromal compart-ments, remodelling of the stroma occurs, which is mediated bykeratinocyte-secreted factors (reviewed by Nelson and Bissell(2006)).
The human dermal fibroblasts formed sturdy, easily handledsheets, with no significant contraction before or after the detach-ment of the sheets from the flask. The two-layered stromalcompartment showed a fairly even distribution of cells throughoutthe abundant matrix after 7 days of submerged culture (Figs. 1 and2A). At day 0 the average thickness of the DF stroma wassignificantly greater in the presence of ucEpi than in that ofkeratinocytes, by 71% (po0.05, Fig. 1H); the same tendency couldbe seen at days 7 and 21 of air–liquid culture.
After thawing, WJC populations had comparable doubling timesin monolayer culture to the reference DF population (Fig. 2H). TheWJC also formed easily manipulable sheets even after only 14 daysof culture with ascorbic acid-supplemented medium. They wereeasily lifted with forceps without tearing. The stromal compart-ments derived from these cells differed noticeably in structure fromthose produced from dermal fibroblasts. For the most part, the WJCmaintained their position between and under layers of the matrixmaterial even after the 21 days of air–liquid incubation, andrelatively few cells were found within the matrix layer. Thus thetwo superimposed stromal sheets were clearly distinguishable inhistological samples at all time points, although adhered together,and formed a dermal compartment that did not seem to be asgreatly affected by remodelling under the influence of keratinocytesinsofar as cell distribution (Figs. 1 and 2D–F), but the dermalthickness decreased over time (Fig. 1H, po0.05 for days 0–21). Incombination with keratinocytes in air–liquid conditions, the aver-age WJC stromal thickness was not significantly different from thatof the DF/K substitutes (Fig. 1H) except for at day 0, where the WJCstroma was almost twice as thick on average as the DF stroma(po0.01).
3.4. Stromal compartment characterisation and basementmembrane formation
The stromal compartment of all substitutes was uniformlypositive for type I and III collagens (Fig. 4A and B and I and J,summarised in Table 2), as well as for fibronectin (Fig. 4C and K).The most striking difference in protein expression was found withalpha-smooth muscle actin, expression of which was detected inthe WJC but not in the dermal fibroblasts (Fig. 4D and L), as in thenative tissues (Hayward et al., 2013). The presence of a well-developed basement membrane is suggested by the detection oflaminin 5 (Fig. 4E and M; Table 2) as well as collagen types IV(Table 2) and VII (Fig. 4G and O; Table 2) in all types of substitutesbut WJC/ucEpi (not shown).
Table 2Immunofluorescence staining results for skin substitutes after 21 days of culture atthe air–liquid interface.
Marker DF/K DF/ucEpi WJC/K
Layer of substitute Stroma Epi Stroma Epi Stroma Epi
Keratin 10 – sb – sb – sbKeratin 12 – – – þ – –
Keratin 14 – þþ – þþ – þþKeratin 16 – sb – sbþ – þKeratin 18 – – – þþ þ 7Keratin 19 – bcþ – þþ 7 þType I collagen þþ – þþ – þþ –
Type III collagen þþ – þþ – þþ –
Type IV collagen þ bm þþ bm þþ bmType VII collagen – bm – bm, bc - bmLaminin 5 7 bm 7 bm, bc 7 bmα-sm actin – – - – þþ –
Fibronectin þþ – þþ – þþ –
Filaggrin – sg – sgþ – sgþInvolucrin – sb – sb – sbTransglutaminase – sg – sg – sg
Epi: epidermal layer of substitute.Stroma: stromal layer of substitute.–: not present.7: very faint or very few cells.þ: present in parts of layer or region.þþ: present everywhere in layer or region.bm: present in basement membrane.bc: present in basal cells.sb: present suprabasally.sg: present in stratum granulosum.
Fig. 3. Evaluation of the epithelial differentiation of bilayered stromal/epithelial substitutes after 7 days of culture at the air–liquid interface. (A–D) WJC/K and (E–H)DF / ucEpi substitutes were analysed for protein expression with fluorescence-coupled antibodies specific for (A, E) K14; (B, F) K10; (C, G) involucrin and (D, H)transglutaminase in red. Hoechst staining of the nuclei is in blue. Arrowheads indicate the position of the basement membrane. Scale bar: 100 mm.
C.J. Hayward et al. / Differentiation 87 (2014) 172–181 177
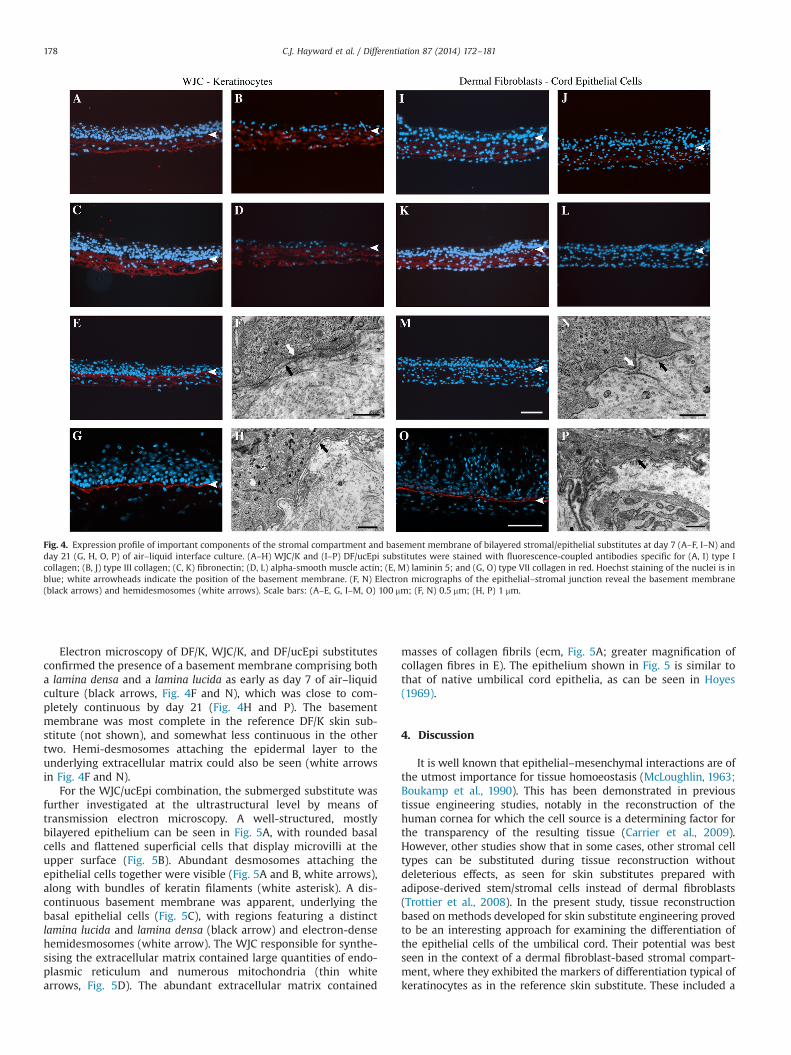
Electron microscopy of DF/K, WJC/K, and DF/ucEpi substitutesconfirmed the presence of a basement membrane comprising botha lamina densa and a lamina lucida as early as day 7 of air–liquidculture (black arrows, Fig. 4F and N), which was close to com-pletely continuous by day 21 (Fig. 4H and P). The basementmembrane was most complete in the reference DF/K skin sub-stitute (not shown), and somewhat less continuous in the othertwo. Hemi-desmosomes attaching the epidermal layer to theunderlying extracellular matrix could also be seen (white arrowsin Fig. 4F and N).
For the WJC/ucEpi combination, the submerged substitute wasfurther investigated at the ultrastructural level by means oftransmission electron microscopy. A well-structured, mostlybilayered epithelium can be seen in Fig. 5A, with rounded basalcells and flattened superficial cells that display microvilli at theupper surface (Fig. 5B). Abundant desmosomes attaching theepithelial cells together were visible (Fig. 5A and B, white arrows),along with bundles of keratin filaments (white asterisk). A dis-continuous basement membrane was apparent, underlying thebasal epithelial cells (Fig. 5C), with regions featuring a distinctlamina lucida and lamina densa (black arrow) and electron-densehemidesmosomes (white arrow). The WJC responsible for synthe-sising the extracellular matrix contained large quantities of endo-plasmic reticulum and numerous mitochondria (thin whitearrows, Fig. 5D). The abundant extracellular matrix contained
masses of collagen fibrils (ecm, Fig. 5A; greater magnification ofcollagen fibres in E). The epithelium shown in Fig. 5 is similar tothat of native umbilical cord epithelia, as can be seen in Hoyes(1969).
4. Discussion
It is well known that epithelial–mesenchymal interactions are ofthe utmost importance for tissue homoeostasis (McLoughlin, 1963;Boukamp et al., 1990). This has been demonstrated in previoustissue engineering studies, notably in the reconstruction of thehuman cornea for which the cell source is a determining factor forthe transparency of the resulting tissue (Carrier et al., 2009).However, other studies show that in some cases, other stromal celltypes can be substituted during tissue reconstruction withoutdeleterious effects, as seen for skin substitutes prepared withadipose-derived stem/stromal cells instead of dermal fibroblasts(Trottier et al., 2008). In the present study, tissue reconstructionbased on methods developed for skin substitute engineering provedto be an interesting approach for examining the differentiation ofthe epithelial cells of the umbilical cord. Their potential was bestseen in the context of a dermal fibroblast-based stromal compart-ment, where they exhibited the markers of differentiation typical ofkeratinocytes as in the reference skin substitute. These included a
Fig. 4. Expression profile of important components of the stromal compartment and basement membrane of bilayered stromal/epithelial substitutes at day 7 (A–F, I–N) andday 21 (G, H, O, P) of air–liquid interface culture. (A–H) WJC/K and (I–P) DF/ucEpi substitutes were stained with fluorescence-coupled antibodies specific for (A, I) type Icollagen; (B, J) type III collagen; (C, K) fibronectin; (D, L) alpha-smooth muscle actin; (E, M) laminin 5; and (G, O) type VII collagen in red. Hoechst staining of the nuclei is inblue; white arrowheads indicate the position of the basement membrane. (F, N) Electron micrographs of the epithelial–stromal junction reveal the basement membrane(black arrows) and hemidesmosomes (white arrows). Scale bars: (A–E, G, I–M, O) 100 mm; (F, N) 0.5 mm; (H, P) 1 mm.
C.J. Hayward et al. / Differentiation 87 (2014) 172–181178

multilayered keratinised epithelium with increasingly flattenedcells near the surface, and the sustained suprabasal presence ofK10, involucrin, transglutaminase and filaggrin. The development ofa cornified layer was absent in the epithelial compartment,although some desquamation was evident. The basement mem-brane markers were well developed, with a sustained presence oflaminin 5 and types IV and VII collagens along the dermo-epidermal junction. We were also able to verify the ultrastructureof the basement membrane by electron microscopy, confirming thepresence of hemidesmosomes anchoring the basal epithelial cells tothe stroma, and of both a lamina lucida and a lamina densa as earlyas day 7 of air–liquid culture. The continuity and maturity of thecomplete basement membrane increased with time in culture. Thisis in contrast to previous studies (Mizoguchi et al., 2004; Huang etal., 2011) in which ucEpi were seeded on fibroblast-populatedcollagen gels, and where only intermittent type IV collagen andlittle to no type VII collagen were noted in the basement membraneafter ten days of exposure to air. The former study found noultrastructural evidence of basement membrane components; onlyby transplanting the substitutes to athymic mice were the investi-gators able to observe complete basement membrane structures(Sanmano et al., 2005). The very physiological nature of the self-assembled dermal compartments of our substitutes is likely acontributing factor to the proper development of the basementmembrane in vitro (Lavoie et al., 2013).
The connective tissues forming native skin and umbilical cordare dense, with stromal cells distributed throughout the matrix, andthe dermal substitutes formed from dermal fibroblasts reproducedthis pattern. The stromal cells of the Wharton's jelly, however, couldnot always be maintained in culture with ascorbic acid for theintended 28 days of tissue production, as they eventually retractedinto a small wad after a certain time of culture, which ranged from10 to 30 days depending on the cell population used. This is likelydue to the ubiquitous presence of α-smooth muscle actin in theWJCpopulations, which may play a role in the contraction of theumbilical cord blood vessels in vivo (Takechi et al., 1993). Extendedsheets of WJC were easily lifted, and once detached from the culture
flasks and placed in submerged culture as dermal substitutes,these sheets remained stable and showed no further excessivelycontractile tendencies. Most unexpectedly, though, the stromal cellsdid not redistribute themselves throughout the matrix they hadproduced, but mostly remained clustered in distinct and separatelayers, each underlying its band of collagen-rich matrix material. Asimilar result was obtained with a reconstructed adventitia madefrom sheets of WJC (unpublished results, see also L’Heureux et al.(1998)).
Virtually all the basal cells of the reference DF/K substitutecontained K19, due to the fact that newborn foreskin keratinocyteswere used, which have a much higher percentage of K19-positiveepithelial-stem-cell-like cells than do the keratinocytes of adultskin (Michel et al., 1996). We chose newborn keratinocytes for thisexperiment as being closest in gestational age to the cells of thefull-term umbilical cord, and their similarity has been documentedin previous publications (Ng et al., 2008; Ruetze et al., 2008).The umbilical cord epithelial cells were consistently positive forK19 in all layers of the substitutes, as well as for K12 and K13, asthey are in vivo (Hayward et al., 2013).
Keratinocytes seeded on WJC dermal substitutes showed pro-gress towards epithelial differentiation at day 7, and at day 21 mostareas of these substitutes had a relatively thin stratified anddifferentiated epithelium with flattened, desquamating superficialcells. The basement membrane components were also present inthese substitutes, although with less complete coverage than in thereference DF/K substitute, demonstrating that WJC can function as adermal substrate for skin-like differentiation where keratinocytesare concerned. Cord epithelial cells, however, developed similarskin-like differentiation patterns only when cultured on DF dermalsubstitutes; air exposure alone was not sufficient to promote suchdifferentiation when only UC-derived cell types were included inthe substitutes. In contrast, under submerged conditions both cordepithelial cells and keratinocytes formed thicker, more even layerson the WJC than on dermal fibroblasts. Thus, it would seem evidentthat both substrate (cell source and matrix) and environment (airexposure) play a role in the differentiation of a skin-like epithelium.
Fig. 5. Ultrastructural analysis of the WJC/ucEpi substitute after 7 days of immerged culture conditions. (A) Transmission electron micrographs show the full thickness of theepithelium and part of the stroma. (B) Details of the epithelial surface layer are shown, including microvilli, bundles of keratin filaments, mitochondria and desmosomes.(C) Electron-dense hemidesmosomes appear along the basement membrane, and (D) abundant endoplasmic reticulum and mitochondria are evident in the cytoplasm of thestromal cells. (E) Masses of collagen fibrils compose the extracellular matrix of the stromal compartment of the substitutes. Scale bars: (A) 4 mm; (B, C) 0.5 mm; (D, E) 1 mm.ecm—extracellular matrix; epi—epithelial cell; f—fibroblastic cell; g—Golgibody; n—nucleus; wide black arrow—basement membrane; wide white arrow—desmosomes (A),hemidesmosomes (C); thin white arrow—mitochondria; white *—keratin bundles.
C.J. Hayward et al. / Differentiation 87 (2014) 172–181 179

It is clear that not all combinations of cells would lead to theproduction of a human substitute suitable for grafting in a skinreplacement setting. The properties shown by umbilical cord cell-containing substitutes in submerged conditions would be moreeffective as a replacement for mucosal tissues in a moist environment,where these cells thrive. The WJC form a supportive stroma in theseconditions, where even after only 7 days of co-culture hemidesmo-somes have formed along the epidermal–stromal interface.
In conclusion, these results support the premise that the tissuefrom which cells originate can greatly affect the properties andhomoeostasis of reconstructed substitutes featuring both epithelialand stromal compartments. The epithelial cells of the umbilical corddemonstrate an ability to adopt a multi-stratified, keratinised differ-entiated state when placed in adequate conditions and in thepresence of stimuli from dermal fibroblasts. However, in combinationwith WJC, these cells produce a non-keratinised epithelium, which isunable to sustain exposure to air. In contrast, keratinocytes culturedon WJC are able to differentiate as they do on dermal fibroblasts,with a slightly diminished response. Thus, by varying the combina-tions of cell sources and culture conditions, substitutes featuringdifferent degrees of epithelial differentiation could be produced,ranging from stratified, keratinised skin substitutes to replacementsfor epithelia in internal, moist environments.
Acknowledgements
The authors would like to thank Kathleen Baker, Eric Grand-bois, Cindy Perron, Véronique Racine, Israël Martel and SophieRoberge for their technical expertise, Claude Marin for his photo-graphic assistance, Richard Janvier and the electron microscopefacility staff at Laval University for sample preparation and theircourteous advice for taking the micrographs, and Dan Lacroix forcritical reading and constructive suggestions during the prepara-tion of the manuscript. This work was supported by the CanadianInstitutes for Health Research (CIHR) Grant #MOP-14364 to FAA,Les Fonds de la Recherche du Québec – Santé (FRQS) and LaFondation des Pompiers du Québec pour les Grands Brûlés (FAA),the Réseau Thécell du FRQS, and in part by the CIHR Grant#111233 to JF. CJH was the recipient of a scholarship from theNational Science and Engineering Research Council. PMM is therecipient of a CIHR scholarship. JF is an FRQS Junior 2 Career Awardscholar.
References
Auger, F.A., López Valle, C.A., Guignard, R., Tremblay, N., Noël, B., Goulet, F.,Germain, L., 1995. Skin equivalent produced with human collagen. In Vitr. Cell.Dev. Biol. Anim. 31, 432–439.
Auger, F.A., Berthod, F., Moulin, V., Pouliot, R., Germain, L., 2004. Tissue-engineeredskin substitutes: from in vitro constructs to in vivo applications. Biotechnol.Appl. Biochem. 39, 263–275.
Bachetti, T., Morbidelli, L., 2000. Endothelial cells in culture: a model for studyingvascular functions. Pharmacol. Res. 42, 9–19.
Bernot, K.M., Coulombe, P.A., McGowan, K.M., 2002. Keratin 16 expression defines asubset of epithelial cells during skin morphogenesis and the hair cycle. J.Investig. Dermatol. 119, 1137–1149.
Boa, O., Cloutier, C.B., Genest, H., Labbé, R., Rodrigue, B., Soucy, J., Roy, M., Arsenault, F.,Ospina, C.E., Dubé, N., Rochon, M.H., Larouche, D., Moulin, V.J., Germain, L., Auger,F.A., 2013. Prospective study on the treatment of lower-extremity chronic venousand mixed ulcers using tissue-engineered skin substitute made by the self-assembly approach. Adv. Skin Wound Care 26, 400–409.
Boukamp, P., Breitkreutz, D., Stark, H.J., Fusenig, N.E., 1990. Mesenchyme-mediatedand endogenous regulation of growth and differentiation of human skinkeratinocytes derived from different body sites. Differentiation 44, 150–161.
Carrier, P., Deschambeault, A., Audet, C., Talbot, M., Gauvin, R., Giasson, C.J., Auger, F.A.,Guerin, S.L., Germain, L., 2009. Impact of cell source on human cornea reconstructedby tissue engineering. Investig. Ophthalmol. Vis. Sci. 50, 2645–2652.
Ennis, J., Gotherstrom, C., Le Blanc, K., Davies, J.E., 2008. in vitro immunologicproperties of human umbilical cord perivascular cells. Cytotherapy 10, 174–181.
Font, J., Braut-Boucher, F., Pichon, J., Noel-Hudson, M.S., Muriel, M.P., Bonnet, M.,Wepierre, J., Aubery, M., 1994. A new three-dimensional culture of humankeratinocytes: optimization of differentiation. Cell Biol. Toxicol. 10, 353–359.
Gauvin, R., Guillemette, M., Galbraith, T., Bourget, J.M., Larouche, D., Marcoux, H.,Aube, D., Hayward, C., Auger, F.A., Germain, L., 2011. Mechanical properties oftissue-engineered vascular constructs produced using arterial or venous cells.Tissue Eng. Part A 17, 2049–2059.
Germain, L., Noël, M., Gourdeau, H., Marceau, N., 1988. Promotion of growth anddifferentiation of rat ductular oval cells in primary culture. Cancer Res. 48,368–378.
Germain, L., Rouabhia, M., Guignard, R., Carrier, L., Bouvard, V., Auger, F.A., 1993.Improvement of human keratinocyte isolation and culture using thermolysin.Burns 19, 99–104.
Gibot, L., Galbraith, T., Huot, J., Auger, F.A., 2010. A preexisting microvascularnetwork benefits in vivo revascularization of a microvascularized tissue-engineered skin substitute. Tissue Eng. Part A 16, 3199–3206.
Guillemette, M.D., Gauvin, R., Perron, C., Labbe, R., Germain, L., Auger, F.A., 2010.Tissue-engineered vascular adventitia with vasa vasorum improves graftintegration and vascularization through inosculation. Tissue Eng. Part A 16,2617–2626.
Hayward, C.J., Fradette, J., Galbraith, T., Remy, M., Guignard, R., Gauvin, R., Germain,L., Auger, F.A., 2013. Harvesting the potential of the human umbilical cord:isolation and characterisation of four cell types for tissue engineering applica-tions. Cells Tissues Organs 197, 37–54.
Hoyes, A.D., 1969. Ultrastructure of the epithelium of the human umbilical cord.J. Anat. 105, 149–162.
Huang, L., Wong, Y.P., Gu, H., Cai, Y.J., Ho, Y., Wang, C.C., Leung, T.Y., Burd, A., 2011.Stem cell-like properties of human umbilical cord lining epithelial cells and thepotential for epidermal reconstitution. Cytotherapy 13, 145–155.
Jaffe, E.A., Hoyer, L.W., Nachman, R.L., 1973. Synthesis of antihemophilic factorantigen by cultured human endothelial cells. J. Clin. Investig. 52, 2757–2764.
L’Heureux, N., Pâquet, S., Labbé, R., Germain, L., Auger, F.A., 1998. A completelybiological tissue-engineered human blood vessel. FASEB J. 12, 47–56.
Larouche, D., Paquet, C., Fradette, J., Carrier, P., Auger, F.A., Germain, L., 2009.Regeneration of skin and cornea by tissue engineering. Methods Mol. Biol. 482,233–256.
Lavoie, A., Fugère, C., Beauparlant, A., Goyer, B., Larouche, D., Paquet, C., Desgagné, M.,Sauvé, S., Robitaille, H., Dunnwald, M., Kim, Dong H., Pouliot, R., Fradette, J.,Germain, L., 2013. Human epithelial stem cells persist within tissue-engineeredskin produced by the self-assembly approach. Tissue Eng. Part A 19, 1023–1038.
Marcus, A.J., Woodbury, D., 2008. Fetal stem cells from extra-embryonic tissues: donot discard. J. Cell Mol. Med. 12, 730–742.
McLoughlin, C.B., 1963. Mesenchymal influences on epithelial differentiation.Symp. Soc. Exp. Biol. 17, 359–388.
Michel, M., L’Heureux, N., Pouliot, R., Xu, W., Auger, F.A., Germain, L., 1999.Characterization of a new tissue-engineered human skin equivalent with hair.In Vitr. Cell Dev. Biol. Anim. 35, 318–326.
Michel, M., Tor̈ok̈, N., Godbout, M.-J., Lussier, M., Gaudreau, P., Royal, A., Germain, L.,1996. Keratin 19 as a biochemical marker of skin stem cells in vivo and in vitro:keratin 19 expressing cells are differentially localized in function of anatomicsites, and their number varies with donor age and culture stage. J. Cell Sci. 109,1017–1028.
Mizoguchi, M., Ikeda, S., Suga, Y., Ogawa, H., 2000. Expression of cytokeratins andcornified cell envelope-associated proteins in umbilical cord epithelium: acomparative study of the umbilical cord, amniotic epithelia and fetal skin.J. Invest. Dermatol. 115, 133–134.
Mizoguchi, M., Suga, Y., Sanmano, B., Ikeda, S., Ogawa, H., 2004. Organotypic cultureand surface plantation using umbilical cord epithelial cells: morphogenesis andexpression of differentiation markers mimicking cutaneous epidermis.J. Dermatol. Sci. 35, 199–206.
Nelson, C.M., Bissell, M.J., 2006. Of extracellular matrix, scaffolds, and signaling:tissue architecture regulates development, homeostasis, and cancer. Ann. Rev.Cell Biol. 22, 287–309.
Ng, W., Nishiyama, C., Mizoguchi, M., Nakano, N., Suga, Y., Ikeda, S., Itoh, S.,Kinoshita, K., Okumura, K., Ogawa, H., 2008. Human umbilical cord epithelialcells express Notch1: implications for its epidermal-like differentiation.J. Dermatol. Sci. 49, 143–152.
Owens, G.K., 1995. Regulation of differentiation of vascular smooth muscle cells.Physiol. Rev. 75, 487–517.
Paquet, C., Larouche, D., Bisson, F., Proulx, S., Simard-Bisson, C., Gaudreault, M.,Robitaille, H., Carrier, P., Martel, I., Duranceau, L., Auger, F.A., Fradette, J.,Guérin, S.L., Germain, L., 2010. Tissue engineering of skin and cornea: devel-opment of new models for in vitro studies. Ann. N. Y. Acad. Sci. 1197, 166–177.
Pruniéras, M., Régnier, M., Woodley, D., 1983. Methods for cultivation of keratino-cytes with an air–liquid interface. J. Invest. Dermatol. 81, 28s–33s.
Rochon, M.H., Fradette, J., Fortin, V., Tomasetig, F., Roberge, C.J., Baker, K., Berthod, F.,Auger, F.A., Germain, L., 2010. Normal human epithelial cells regulate the sizeand morphology of tissue-engineered capillaries. Tissue Eng. Part A 16,1457–1468.
Ruetze, M., Gallinat, S., Lim, I.J., Chow, E., Phan, T.T., Staeb, F., Wenck, H., Deppert, W.,Knott, A., 2008. Common features of umbilical cord epithelial cells and epidermalkeratinocytes. J. Dermatol. Sci. 50, 227–231.
Sanmano, B., Mizoguchi, M., Suga, Y., Ikeda, S., Ogawa, H., 2005. Engraftment ofumbilical cord epithelial cells in athymic mice: in an attempt to improvereconstructed skin equivalents used as epithelial composite. J. Dermatol. Sci. 37,29–39.
C.J. Hayward et al. / Differentiation 87 (2014) 172–181180

Stephan, S., Ball, S.G., Williamson, M., Bax, D.V., Lomas, A., Shuttleworth, C.A.,Kielty, C.M., 2006. Cell-matrix biology in vascular tissue engineering. J. Anat.209, 495–502.
Takechi, K., Kuwabara, Y., Mizuno, M., 1993. Ultrastructural and immunohisto-chemical studies of Wharton's jelly umbilical cord cells. Placenta 14,235–245.
Trottier, V., Marceau-Fortier, G., Germain, L., Vincent, C., Fradette, J., 2008. IFATScollection: using human adipose-derived stem/stromal cells for the productionof new skin substitutes. Stem Cells 26, 2713–2723.
Troyer, D.L., Weiss, M.L., 2008. Wharton's jelly-derived cells are a primitive stromalcell population. Stem Cells 26, 591–599.
Tsigkou, O., Pomerantseva, I., Spencer, J.A., Redondo, P.A., Hart, A.R., O’Doherty, E.,Lin, Y., Friedrich, C.C., Daheron, L., Lin, C.P., Sundback, C.A., Vacanti, J.P.,Neville, C., 2010. Engineered vascularized bone grafts. Proc. Natl. Acad. Sci.USA 107, 3311–3316.
Weiss, M.L., Anderson, C., Medicetty, S., Seshareddy, K.B., Weiss, R.J., VanderWerff, I.,Troyer, D., McIntosh, K.R., 2008. Immune properties of human umbilical cordWharton's jelly-derived cells. Stem Cells 26, 2865–2874.
C.J. Hayward et al. / Differentiation 87 (2014) 172–181 181