Genetic characterization of Trypanosoma brucei gambiense
and clinical evolution of human African trypanosomiasis
in Cote d’Ivoire
V. Jamonneau1, A. Garcia2, S. Ravel3, G. Cuny3, B. Oury4, P. Solano1, P. N’Guessan1, L. N’Dri1, R. Sanon1,
J. L. Frezil3 and P. Truc5
1 Institut de Recherche pour le Developpement (IRD/UR 035), Centre Pierre Richet, Bouake, Cote d’Ivoire2 Institut de Recherche pour le Developpement (IRD/UR 010), Dakar, Senegal3 Institut de Recherche pour le Developpement (IRD/UR 035), Laboratoire de Recherche et de Coordination sur les Trypanosomoses,
Montpellier, France4 Institut de Recherche pour le Developpement (IRD/UR 062), Centre d’Etude sur le Polymorphisme des Microorganismes, Montpellier,
France5 Institut de Recherche pour le Developpement (IRD/UR 035), OCEAC, Yaounde, Cameroon
Summary Human African trypanosomiasis is a parasitic infection caused by protozoa belonging to Trypanosoma
brucei subspecies. The clinical evolution of this disease is complex and might be because of the
parasite itself, as genetic diversity has been observed in T. brucei ssp. We investigated the relationship
between the genetic diversity of trypanosomes and the diversity of clinical patterns in Cote d’Ivoire.
We studied clinical sleeping sickness cases, and genetically analysed the trypanosomes isolated from
these patients. An important genetic monomorphism among stocks isolated in Cote d’Ivoire was
observed by using various markers: isoenzymes electrophoresis, random amplified polymorphism DNA
and PCR of microsatellite sequences. At the same time, the diversity of clinical patterns and evolutions
was confirmed by clinical analysis. The existence of an individual susceptibility to disease (human
trypanotolerance) should be taken into account even if our genetic conclusions might be distorted
because the isolation success rates were particularly poor. In fact, we observed that the isolation success
rate varied significantly depending both on the focus of origin (P ¼ 0.0002) and on the ethnic group
(P ¼ 0.0317) of the patient. Further investigations are required in order to study a possible selective
impact of the use of the kit for in vitro isolation of trypanosomes as an isolation technique.
keywords Trypanosoma brucei gambiense, isoenzymes electrophoresis, RAPD, individual
susceptibility, Cote d’Ivoire
correspondence P. Solano, Institut de Recherche pour le Developpement (IRD), UR 035, Institut Pierre
Richet, 01 BP 1500 Bouake, Cote d’Ivoire. Fax: +225 31 63 27 38; E-mail: [email protected]
Introduction
Human African trypanosomiasis (HAT), or sleeping sick-
ness, is a major public health problem in sub-Saharan
Africa. Approximately 60 million people are daily exposed
to the risk of infection. It is estimated that there are about
500 000 infected but untreated persons (WHO 1998). The
pathogenic agent is the trypanosome Trypanosoma brucei.
Classically, T. brucei is subdivided into three subspecies on
the basis of extrinsic criteria: T. b. gambiense is responsible
for the chronic form in West and Central Africa, T. b.
rhodesiense is the agent of the acute form in East Africa,
and T. b. brucei, a parasite of cattle, is supposed to be non-
pathogenic to humans.
By definition, HAT evolves in two phases: a haemato-
lymphatic stage (first period), for which there are no
specific clinical signs (Jannin et al. 1993; Dumas &
Bouteille 1996), leading to a meningo-encephalitic stage,
usually characterized by neurological disorders (second
period). In the absence of treatment, the disease is
invariably fatal. In T. b. gambiense chronic form, the
duration of the first period may be several years: the
fluctuating parasitaemia remains low and tends to
decrease. The appearance of neurological disorders during
Tropical Medicine and International Health
volume 7 no 7 pp 610–621 july 2002
610 ª 2002 Blackwell Science Ltd

the second period is often progressive. In acute HAT
caused by T. b. rhodesiense, the haemato-lymphatic stage
lasts a few weeks to a few months, passage to the meningo-
encephalitic stage is brutal and death can occur within a
few months of onset of this stage.
A diversity of clinical evolutions has been observed for
T. b. gambiense, from some chronic forms to asympto-
matic forms (Jamonneau et al. 2000a). At one extreme, a
patient from Togo carried trypanosomes (T. b. gamb-
iense) for 21 years with no clinical signs or neurological
disorders (Lapeyssonnie 1960). At the other extreme,
some patients detected in Cote d’Ivoire presented a
clinical evolution characteristic of acute HAT (Truc et al.
1997a). This clinical diversity does not correspond to the
traditional definition of the T. b. gambiense clinical forms
of HAT.
Genetic diversity within T. b. gambiense has been
demonstrated by several studies using molecular markers
(Gibson 1986; Paindavoine et al. 1986; Godfrey et al.
1990; Hide et al. 1990), but its influence on the clinical
evolution of the HAT remains unproven. We intended
to investigate the relationships between the genetic
diversity of the parasite and that of the clinical patterns.
With this aim, we launched a clinical study on patients
diagnosed in Cote d’Ivoire together with a genetic
analysis of the populations of trypanosomes isolated
from these patients.
Patients and methods
Study areas and patients
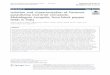
This study was conducted from 1996 to 1999 in two active
HAT foci of Cote d’Ivoire: the western-central part of the
country (Daloa, Vavoua, Bouafle, Sinfra, and Bonon) and
the south-east (Aboisso) (Figure 1). We included patients
detected (parasitological evidence) either actively (during
five medical surveys) or passively (patients presenting
themselves for treatment). All patients were treated in the
three centres specialized in HAT treatment: the Projet de
Recherches Cliniques sur les Trypanosomoses in Daloa,
and the local health centres in Bouafle and Aboisso.
Patients were told of the objectives and protocol of the
study, and only those who gave their consent were included
in the study (patients younger than 10 years were not
included).
Epidemiological data collection
For each patient, the following data were recorded: sex,
age, nationality, ethnic group, geographical origin
(south-east or mid-west), HAT focus of provenance (Sinfra,
Bonon, Daloa, Bouafle or Aboisso), occupation, existence
of family history of HAT, time and mode of diagnosis
(passive or active), and treatment schedule. We distin-
guished two ethnic grouping groups: natives (including
patients born in the study areas) and migrants (including
subjects from northern Cote d’Ivoire, Burkina Faso and
Mali).
Clinical assessment before treatment
Objective clinical signs that were looked for were:
• palpation (hepatomegaly, splenomegaly, swollen lateral
cervical lymph nodes);
• cardiovascular investigation (dysrhythmia, heart
murmurs and low blood pressure);
• dermatological examination (search for initial lesion of
inoculation, trypanids);
• assessing possible endocrinological disorders (impotence,
facial oedema, amenorrhea, abortion);
• neurological examination (alteration of mental state,
abnormal reflexes, tone disorders, sensory disorders,
coordination disorders).
A questionnaire presented to patients covered subjective
clinical signs such as asthenia, anorexia, cachexia, fevers,
repeated headache, nausea, pruritus, cutaneous rash, sleep
disturbances (alteration of circadian rhythm). Questions
about the approximate date of appearance of the initial
symptoms and the mode of evolution of the disease were
also included.
On the basis of clinical signs, patients were classified
according to the existence and the importance of the
following: infectious or inflammatory syndrome, cardio-
vascular syndrome, digestive syndrome, dermatological
syndrome and neuro-psychiatric syndrome (Table 1).
Serological and parasitological examinations,
and stage determination
Each patient underwent serological and parasitological
investigations. For serology, the Card Agglutination Test
for Trypanosomiasis (T. b. gambiense) was performed
using whole blood and plasma (Magnus et al. 1978).
Trypanosomes were detected using the mini Anion
Exchange Centrifugation Technique (Lumsden et al. 1979)
and by direct microscopic examination of the lymphatic
fluid if lymph nodes were swollen.
For stage determination, the tests used on the cereb-
rospinal fluid (CSF) were: trypanosome detection by
double centrifugation (Cattand et al. 1988) and leukocyte
counting using a Nageotte counting chamber. A low
leukocyte count (£ 5 cells/ll) combined with absence of
Tropical Medicine and International Health volume 7 no 7 pp 610–621 july 2002
V. Jamonneau et al. Human African trypanosomiasis in Cote d’Ivoire (Ivory Coast)
ª 2002 Blackwell Science Ltd 611

trypanosomes in CSF is defined as the first stage of the
disease. An elevated leukocyte count (>5 cells/ll), with or
without trypanosomes in CSF, forms the basis for second-
stage diagnosis (WHO 1998).
Isolation of trypanosomes
The stocks were isolated by using the KIVI (Kit for In
vitro Isolation of trypanosomes, Aerts et al. 1992) and
were then multiplied using semidefined culture medium
(Cunningham 1977) according to the protocol described
by Truc et al. (1992). For each stock, two pellets of
trypanosomes were obtained by centrifugation and stored
in liquid nitrogen. Reference stocks (Tables 2 and 3) had
been isolated with the KIVI method during or after
1991, the others having been isolated by rodent inocu-
lation.
Multilocus enzyme electrophoresis
Proteins were extracted from one of the two pellets (Truc
et al. 1991). Stocks were characterized by the technique of
multilocus enzyme electrophoresis (MLEE) on cellulose
acetate plates and 11 enzymatic systems were revealed:
ALAT (EC.2.6.1.2), GOT (EC.2.6.1.1), Nhi (EC.3.2.2.1),
Nhd (EC.3.2.2.1), ME (EC.1.1.1.40), PEP-2 (EC.3.4.11),
MALI
GUINEA
BURKINA FASO
Bouake
Vavoua
BononDaloa
Sinfra
Bouafle Yamoussoukro
LIBERIA
GHANA
Aboisso
Abidjan
ATLANTIC OCEAN
LEGEND:RiverLake
Towns of the study areaOther towns
Scale0 25 50 75 100 km
N
Figure 1 Geographic location of the studyarea.
Tropical Medicine and International Health volume 7 no 7 pp 610–621 july 2002
V. Jamonneau et al. Human African trypanosomiasis in Cote d’Ivoire (Ivory Coast)
612 ª 2002 Blackwell Science Ltd

MDH (EC.1.1.1.37), IDH (1.1.1.42), TDH (1.1.1.103),
PGM (EC.2.7.5.1) according to Truc et al. (1991) and
Truc and Tibayrenc (1993), and SOD (EC.1.15.1.1)
according to Stevens et al. (1989).
Random amplified polymorphism DNA
The DNA was extracted from the second pellet according to
the protocol described by Oury et al. (1998). Stocks were
characterized by the RAPD technique (random amplified
polymorphism DNA, Welsh & McClelland 1990; Williams
et al. 1990; Tibayrenc et al. 1993). Seven primers were used:
A2 (5¢-TGCCGAGCTG-3¢), A4 (5¢-AATCGGGCTG-3¢),A7 (5¢-GAAACGGGTG-3¢), A8 (5¢-GTGACGTAGG-3¢),A10 (5¢-GTGATCGCAG-3¢), A11 (5¢-CAATCGCCGT-3¢)and A18 (5¢-AGGTGACCGT-3¢).
Polymorphism of chain reaction of microsatellite DNA
sequences
From the same DNA extraction, microsatellite sequences
(PCR/microsatellite) were amplified with four primer pairs:
M6C8-CAF/R, MT30-33F/R (Biteau et al. 2000),
TRBPA1/2 (Simo et al. 2000) and TBDAC1/2 (Truc et al.
2002). Amplifications followed the protocol described by
Truc et al. (2002).
Analysis procedures
Whenever possible, a comparison of two qualitative
variables was performed by means of Pearson chi-square
tests. Alternatively, we performed Fisher’s exact test. The
level of significance retained for the tests was 5%. These
Table 1 Clinical signs and corresponding syndromes
Syndrome Clinical signs Expected answers
Inflammatory/infectious Temperature at inclusion Quantitativesyndrome Asthenia Absence/presence
Anorexia Absence/presenceWeight loss Absence/presence(feeling) Fever Absence/presenceRecurrent headache Absence/presenceCervical lymph nodes Absence/presence
Digestive syndrome Nausea Absence/presenceVomiting Absence/presenceHepatomegaly Absence/presenceSplenomegaly Absence/presence
Cardiovascular syndrome Arrhythmia (heart rhythm) Normal/abnormalHeart rate Normal/abnormalBlood pressure Normal/abnormalSubjective heart troubles Absence/presence
Dermatological syndrome Chancre Absence/presenceTrypanids Absence/presenceOedema Absence/presencePruritus Absence/presenceSkin rash Absence/presence
Neuro-psychiatric syndrome Sleep disorders Absence/diurnal drowsiness/night insomniaEating disorders Absence/lack of appetite/compulsive eatingThirst disorders Absence/polydipsiaSexual disorders Absence/lower libido/lost libido/sexual impotenceSensibility disorders Absence/hyperpathyConsciousness disorders Absence/lower consciousness/mental confusion/comaBehavioural disorders Absence/agitation/indifferenceEmotional disorders Absence/euphoria/sadness/aggressivenessMotor disorders Absence/lower motility/no motilityCoordination disorders Absence/impaired coordinationMuscular tonus disorders Absence/hypertonicity/hypotonicityArchaic reflexes Absence/presenceOsteo-tendinous reflexes Normal/excessive/abolitionPlantar-skin reflexes Normal/excessive/abolitionObjective sensitivity Normal/insensitivity
Tropical Medicine and International Health volume 7 no 7 pp 610–621 july 2002
V. Jamonneau et al. Human African trypanosomiasis in Cote d’Ivoire (Ivory Coast)
ª 2002 Blackwell Science Ltd 613

analyses were done using BMDP software (BMDP statis-
tical Software, University of California, Los Angeles, CA,
USA).
Unweighted Pair Group Method Analysis (UPGMA)
dendrograms were built, starting with the Jacquard genetic
distances (Jacquard 1973) calculated from MLEE and
RAPD results for visualizing the relationships between
stocks (Sneath & Sokal 1973). Reference stocks of T. b.
rhodesiense, T. b. brucei and T. b. gambiense groups 1 and
bouafle were included, as well as stocks of T. congolense-
like groups for the UPGMA comparison (Tables 2 and 3).
For the PCR/microsatellite DNA, the four primers
were specific to T. b. gambiense group 1; thus, only
two reference stocks, Jua and Peya (Table 3), were
used. Each band obtained, defined by its molecular
weight (in base pairs or bp), corresponds to an allele.
For each stock, two bands per primer were revealed
(X bp/Y bp).
Results
Patients
A total of 139 patients (harbouring trypanosomes) partici-
pated in the study; nine came from Aboisso in the South-
East, and of the 130 remaining patients from the midwest, 82
Table 2 Reference stocks used for MLEE characterization
Stock Host Year Country Focus Zymodeme Species Reference
DAL072 Human 1978 CI Vavoua 1 T. b. g 1 1Trazie Human 1991 CI Sinfra 2 T. b. g 1 2Sique Human 1991 CI Sinfra 3 T. b. g 1 2SH017 Human 1989 CI Aboisso 6 T. b. bfl 2SH196 Human 1990 CI Daloa 7 T. b. bfl 2SH276 Human 1992 CI Daloa 10 T. b. bfl 2SINF1 Human 1992 CI Sinfra 11 T. b. g 1 2SINF5 Human 1992 CI Sinfra 12 T. b. g 1 2TH2 Human 1978 CI Daloa 14 T. b. bfl 3TSW53 Pig 1982 CI Bouafle 15 T. b. bfl 1TSW103 Pig 1977 Liberia Sanniquelle 27 T. congo 4132 Kob 1993 CI Comoe 30 T. b. bfl 5KK39 Kob 1980 CI Comoe 33 T. b. bfl 6AB14 Hartebeest 1980 CI Comoe 37 T. b. bfl 11972 Human 1993 CI Sinfra 38 T. b. g 1 2Bub6 Hartebeest 1994 CI Marahoue 40 T. b. g 1 2
CI, Cote d’Ivoire; T. b. g 1, Trypanosoma brucei gambiense group 1; T. b. bfl, Trypanosoma brucei bouafle group; T. congo, Trypanosomacongolense.References: 1. Stevens et al. (1992); 2. Truc et al. (1997a); 3. Mehlitz et al. 1982; 4. Gashumba et al. (1988); 5. Truc et al. (1997b);6. Young and Godfrey (1983).
Stock Host Year Country Specie Reference
Jua* Human 1979 Cameroon T. b. g 1 1Peya* Human 1980 Congo T. b. g 1 1KP465 Pig 1991 Cote d’Ivoire T. b. bfl 2TSW65 Pig 1982 Cote d’Ivoire T. b. bfl 3058clA3 Human 1974 Zambia T. b. rh 4Eatro 1125 w.m. 1966 Uganda T. b. b 5TRPZ105 Dog 1981 Zambia T. congo 6
w.m., Wild mammal; T. b. g 1, Trypanosoma brucei gambiense group 1; T. b. bfl,Trypanosoma brucei bouafle group; T. b. rh, Trypanosoma brucei rhodesiense; T. b. b,Trypanosoma brucei brucei; T. congo., Trypanosoma congolense.* For PCR of microsatellites sequences, only Jua and Peya were used as reference stocks.References: 1. Truc et al. (1991); 2. Truc et al. (1997a); 3. Stevens et al. (1992);4. Gibson et al. (1980); 5. Hide et al. (1990); 6. Gashumba et al. (1988).
Table 3 Reference stocks used for RAPDand PCR/microsatellite characterization*
Tropical Medicine and International Health volume 7 no 7 pp 610–621 july 2002
V. Jamonneau et al. Human African trypanosomiasis in Cote d’Ivoire (Ivory Coast)
614 ª 2002 Blackwell Science Ltd

came from Sinfra, 27 from Bonon, 13 from Bouafle and 8
from Daloa. Sixty-eight per cent of patients were detected
passively and 71 actively through five medical surveys.
Thirty-nine patients were natives (28%) and 100 migrants
(72%). This unequal distribution is traditional in Cote
d’Ivoire as most of the migrants work and live close to the
cocoa and coffee plantations, which constitute areas of
greater HAT transmission risk. One-hundred and nine
patients (78.4%) were farmers and 30 (21.6%) declared a
family history of HAT.
Isolation of trypanosomes
Of the 139 KIVIs performed before treatment, only
62 (44%) gave a positive result allowing the in vitro
culture and multiplication of procyclics. To isolate a
maximum of stocks, another KIVI was inoculated for
46 patients (some of whom had already given positive
KIVI in the first round). Only 20 of these 46 KIVI
(43%) were positive. In total, 64 stocks (46%) were
isolated and cultured for genetic identification.
These isolation success rates were particularly poor.
Thus, we checked whether some of the host characteristics
could have affected the success of isolation. We compared
the epidemiological parameters of the human population
from whom the stocks were isolated (population ‘KIVI
positive’) with the population from whom the isolation
failed (population ‘KIVI negative’). The HAT focus of
origin and the ethnic group had a significant influence on a
positive in vitro isolation (P ¼ 0.0002 and P ¼ 0.0317,
respectively). More precisely, the isolation rate ranged
from 33.7% in the Sinfra focus to 85.2% in the Bonon one.
The isolation rate was particularly poor with the Baoule
group, natives of the area (7.7%) and was most elevated
for the Senoufo migrant group (66.7%).
Isozyme characterization
Of 64 stocks, 61 were zymodeme 3 (Z3, Truc et al. 1997a).
The three other stocks were zymodeme 38 (Z38, Truc et al.
1997a). The dendrogram (Figure 2) shows the relationships
between these two zymodemes, which differ only by the ME
locus: Z3 showed heterozygotes at this locus (three bands),
while Z38 was a homozygote (one band). These two
zymodemes are genetically closely related (d < 0.1). They
both belong to group 1 of T. b. gambiense (Gibson 1986).
Z27/TSW103
Z11/SINF1
Z40/Bub6
Z1/DAL072
Z12/SINF5
Z38/1972
Z2/TRAZIE
Z3/SIQUE
Z30/132
Z14/TH2
Z37/AB14
Z7/SH196
Z33/KK39
Z15/TSW53
Z6/SH017
Z10/SH276
T.b. bouafle
T.b. gam
biense group 1
0.1
T. congolense
Figure 2 UPGMA dendrogram based onthe matrix of Jacquard genetic distancescalculated on isoenzymatic results of 15known zymodemes of the species Trypa-nosoma brucei. T. congolense was chosenas the outgroup and is identified byzymodeme 27. Zymodemes 3 and 38underlined on the tree are the only zymo-demes that have been sampled in this study.
Tropical Medicine and International Health volume 7 no 7 pp 610–621 july 2002
V. Jamonneau et al. Human African trypanosomiasis in Cote d’Ivoire (Ivory Coast)
ª 2002 Blackwell Science Ltd 615

Thus, MLEE results showed very low genetic variability in
our sample.
RAPD characterization
Random amplified polymorphism DNA analysis allows
a very clear distinction between T. congolense and
T. brucei ssp. and a clear individualization of group 1
of T. b. gambiense within T. brucei (Waitumbi &
Murphy 1993). Owing to logistic constraints, only 50
stocks in our study were characterized. They all
belonged to T. b. gambiense group 1. These results
were concordant with those of MLEE, as a low genetic
polymorphism was observed. Of the seven primers
used, only one (A2) showed a microvariability with
three profiles differing between them by only one band
(data not shown). The dendrogram is presented in
Figure 3.
Characterization by PCR/microsatellite DNA
Because of logistic constraints, only 16 stocks were charac-
terized by PCR/microsatellite (Table 4). Using MT30-33
F/R, the 16 stocks showed two bands identical to Jua and
TRPZ105058Cl.A 3Eatro1125KP465TSW656346142582257425602508250724972498659PeyaJuaB120/9806/96706696686666646626616606576556546486386366226126106066002604260326022601260025982597259525882587258425702569256225612557254924992548
0.1
Figure 3 UPGMA dendrogram based onthe Jacquard genetic distances calculated onthe RAPD results of the 50 stocks of ourstudy. Trypanosoma congolense, repre-sented by stock TRPZ 105, was chosen asthe outgroup. Stocks 058CL.A3, Eatro1125, KP465, TSW65, Jua and Peya werechosen as reference stocks for T. brucei.
Tropical Medicine and International Health volume 7 no 7 pp 610–621 july 2002
V. Jamonneau et al. Human African trypanosomiasis in Cote d’Ivoire (Ivory Coast)
616 ª 2002 Blackwell Science Ltd

Peya bands (154 bp/162 bp). Using M6C8-CA F/R, 16
stocks showed two bands identical to those of Peya (85 bp/
89 bp). Two profiles were observed with TRBPA1/2
(149 bp/203 bp for nine stocks and 149 bp/185 bp for seven
stocks). Finally, when using TBDAC1/2, a higher micro-
variability was highlighted showing four different
patterns.
Clinical study
Forty-four patients were in the first stage of the disease (P1),
while 95 were in the second stage (P2). The degree of severity
for the five syndromes is given in Table 5. Syndromes were
not significantly linked to one another, and only the neuro-
psychiatric syndrome was significantly linked to stage of
disease (P < 0.0001), indicating a high diversity of clinical
symptoms among these patients before treatment.
The time of appearance of symptoms was significantly
longer (P ¼ 0.0002) for the patients in P2 than for the
patients in P1 (Table 6). Among the 62 patients feeling
unwell for more than 12 months, 50 (81%) were diag-
nosed in P2. However, of 30 patients whose symptoms
appeared within 6 months, 22 (73%) were already in P2.
Thus, though the appearance of symptoms for more than
12 months seemed to confirm a diagnosis of second stage,
a recent appearance of clinical signs was not necessarily a
criterion for a first-phase diagnosis. This result suggests a
diversity of clinical evolutions.
Correlation between clinical and genetic diversity
We compared the clinical patterns of the patients ‘Z3’
with those of the patients ‘Z38’. Whatever the clinical
variable, there was no significant difference between the
Table 4 Results of the PCR/microsatellitetechnique carried out using four primerpairs, on 16 stocks of our study
Stock TRBPA1/2 TBDAC1/2 M6C8-CAF/R MT30-33F/R
Jua* 149/203 152/160 83/87 154/162Peya* 149/149 154/164 85/89 154/1622499 149/203 154/162 85/89 154/1622508 149/185 150/156 85/89 154/1622562 149/203 154/162 85/89 154/162611 149/203 154/162 85/89 154/162614 149/203 154/162 85/89 154/162622 149/203 154/162 85/89 154/162634 149/203 154/162 85/89 154/162654 149/185 150/156 85/89 154/162659 149/203 152/160 85/89 154/162662 149/185 150/156 85/89 154/162664 149/203 156/164 85/89 154/162666 149/203 156/164 85/89 154/162668 149/185 150/156 85/89 154/162669 149/185 150/156 85/89 154/162384 149/185 152/156 85/89 154/162387 149/185 152/156 85/89 154/162
* Reference stocks (see Table 3).
Table 5 Occurrence of syndromes among139 patients Syndrome Absence Presence Heavy Very heavy Total
Infectious or inflammatory 4 62 73 0 139Cardiovascular 120 16 3 0 139Digestive 89 47 3 0 139Dermatological 50 53 36 0 139Neuro-psychiatric 32 66 29 12 139
The degree of importance of the different syndromes was determined as follows:Infectious or inflammatory, cardiovascular, digestive and dermatological syndrome:absence ¼ no clinical sign, presence ¼ 1–2 clinical signs, and heavy ¼ more than twoclinical signs.Neuro-psychiatric syndrome: absence ¼ no clinical sign, presence ¼ 1–2 clinical signs,heavy ¼ 3–8 clinical signs, and very heavy ¼ more than 8 clinical signs.
Tropical Medicine and International Health volume 7 no 7 pp 610–621 july 2002
V. Jamonneau et al. Human African trypanosomiasis in Cote d’Ivoire (Ivory Coast)
ª 2002 Blackwell Science Ltd 617

two populations by means of Fisher exact test. The low
genetic variability detected by MLEE did not seem to be
correlated to the clinical variability observed. The same
result and conclusion were obtained when comparing the
genetic groups identified by both RAPD and PCR/micro-
satellite analysis. Neither was there any significant differ-
ence between KIVI positive and negative populations in
terms of clinical variables. Thus, genetic variability did not
seem to be related to the observed clinical variability.
Discussion
The isolation rates observed were particularly low com-
pared with previous works where success rates were up to
90% in Cote d’Ivoire and R.P. Congo (Aerts et al. 1992;
Truc et al. 1992). This may be because of the quality of
KIVI batches. To check this hypothesis, two different
batches were used during the whole study but the rates of
isolation remained low whatever the batch used. Circula-
tion of particular stocks in HAT foci of Cote d’Ivoire,
which could be difficult to isolate with KIVI, could also
explain the low rate of isolation. In fact, we observed that
the isolation rate varied significantly depending on the
focus of the origin of the patient. As the composition of the
human populations were similar between one focus and
another, a factor related to the parasite might have an
influence on the isolation success. Further investigations
are required in order to study a possible selective effect of
the use of the KIVI. We also observed that the isolation rate
varied significantly according to the ethnic group. Thus, a
factor related to the host might also have an influence on
the success of in vitro isolation of trypanosomes.
According to MacNamara et al. (1995), the failure or
the success of KIVI may depend first on the proportion of
short (stumpy) forms and long (slender) forms in the blood
of the patient at the moment of KIVI isolation. A high
number of stumpy forms in blood increases the success of
isolating trypanosomes using KIVI because, as within the
tsetse fly, only these stumpy forms are able to be
transformed into procyclic forms and multiply. Further
investigations are required to study the proportions of
stumpy and slender forms in patient’s blood when per-
forming KIVI isolation in various foci while taking into
account the ethnic origin of patient.
The genetic characterization by MLEE and RAPD
revealed a very low genetic polymorphism within the
stocks under study. Some genetic homogeneity among
T. b. gambiense stocks was already known (Gibson 1986;
Paindavoine et al. 1986; Godfrey et al. 1990; Hide et al.
1990) but a polymorphism as low as that described by
Jamonneau et al. (2000b) using 222 stocks isolated in Cote
d’Ivoire from 1992 to 1999 remains unusual. Moreover, a
recent study on the characterization of trypanosome stocks
from various geographical origins using PCR/microsatellite
also highlighted the absence of genetic variability among
nine stocks isolated in Cote d’Ivoire since 1993 belonging
to zymodeme Z3 (Biteau et al. 2000). In the present work,
stocks characterized by PCR/microsatellite analysis were
monomorphic for two primers, whereas a microvariability
was found when using the two other primers. We can
notice that the evaluation of the discriminatory potential of
the primers actually used for PCR/microsatellite is still in
progress. Some patterns obtained with these primers could
be specific to T. b. gambiense group 1 (Biteau et al. 2000;
Truc et al. in press).
Thus, whatever the technique used (MLEE, RAPD, PCR/
microsatellite), a low genetic polymorphism was revealed
within the stocks currently isolated in Cote d’Ivoire. Only
Z3 seems to be able to spread within the whole country,
confirming previous observations (Jamonneau et al.
2000b). However, according to the assumption of a
selection by KIVI, the isolated stocks might not be
representative of the natural populations (KIVI positive
stocks could be then genetically different from KIVI
negative stocks). This hypothesis confirms the need of
studying selectivity in isolation techniques. This could be
done by trying to directly identify trypanosomes within the
biological fluids of man (blood, CSF and lymph juice)
without isolation and culture, and using specific molecular
markers. The technique of PCR/microsatellite seems to be a
promising tool for this purpose, as two primers used in this
study highlighted a microvariability within stocks belong-
ing to the same zymodeme (Z3).
The results of the clinical study indicate a significant
diversity of clinical pictures. This is in accordance with a
previous longitudinal follow-up of patients refusing the
treatment in the Sinfra area between 1996 and 1999, which
revealed the existence of a diversity of clinical evolutions
Date of appearance of the first symptoms
Stage 0–6 months 6–12 months >12 months No response Total
1 8 20 12 4 442 22 15 50 8 95Total 30 35 62 12 139
Table 6 Stage (period) of the disease anddate of appearance of the first symptoms
Tropical Medicine and International Health volume 7 no 7 pp 610–621 july 2002
V. Jamonneau et al. Human African trypanosomiasis in Cote d’Ivoire (Ivory Coast)
618 ª 2002 Blackwell Science Ltd

(Jamonneau et al. 2000a). In this latter work various
clinical patterns were observed, from the chronic form to
an acute form, and self-cure cases were suspected. What-
ever the parasitic disease, diversity of clinical evolution can
be explained either by the virulence of the parasite or by
the host susceptibility to disease. In the particular case of
Chagas disease, Andrade et al. (1992) have suggested that
the various genotypes described within the species
Trypanosoma cruzi (Miles et al. 1981) could be partly
responsible for the diversity of clinical evolutions. Evidence
of a correlation between genotypes and clinical evolutions
was shown through experimental pathology (Laurent et al.
1997), as well as in humans (Montamat et al. 1996). For
HAT caused by T. b. rhodesiense in the Southeast of
Uganda, Smith and Bailey (1997) showed that stocks of the
Busoga group were more pathogenic than stocks of the
Zambezi group using isoenzymatic characterization. In our
study, no correlation between clinical diversity in human
and genetic diversity of parasites (observed or not) could be
shown.
The role of the host in clinical variability could be
considered. The existence of an individual susceptibility to
disease has been shown, for example, in malaria (Garcia
et al. 1998) and leishmaniasis (Mary et al. 1999). It has
been suspected for HAT (Ginoux & Frezil 1981), being
called trypanotolerance. This phenomenon has been des-
cribed for animal trypanosomiasis (Murray et al. 1990;
Authie 1994). There is no experimental data on individual
susceptibility to HAT and its genetic explanation, only
indirect evidence (Authie et al. 1991; Garcia et al. 2000;
Jamonneau et al. 2000a). In our study, the individual
susceptibility to disease seems to better explain the
observed data than the genotype of the infecting
T. b. gambiense strains. The hypothesis of an individual
susceptibility for HAT and its genetic mechanism must be
investigated. The fast evolution of part of the patients to
the second stage of the disease, together with the suspected
existence of trypanotolerant patients, who do not see a
physician and/or refuse treatment because they do not feel
sick, is a new useful plea again for early diagnosis and
treatment and for regular supervision of HAT foci by
National Control Programmes.
Acknowledgements
We thank the HAT team of the Institut Pierre Richet in
Bouake and the National Control Program for HAT of
Cote d’Ivoire. This work was supported by a grant from
the Fonds d’Aide a la Cooperation du Ministere des
Affaires Etrangeres, Direction du Developpement et de la
Cooperation and by the Agence Francaise de la Fran-
cophonie (AUF). We thank two anonymous referees for
their great help in the improvement of the manuscript, and
we thank G. Manners for editorial advice.
References
Aerts D, Truc P, Penchenier L, Claes Y & Le Ray D (1992) A kit
for in vitro isolation of trypanosomes in the field: first trial
with sleeping sickness patients in the Congo. Transactions of
the Royal Society of Tropical Medicine and Hygiene 86,
394–395.
Andrade SG, Rassi A, Magalhaes JB, Ferriolli Filho F & Luquetti
AO (1992) Specific chemotherapy of Chagas disease. A com-
parison between the response in patients and experimental
animals inoculated with the same strains. Transactions of the
Royal Society of Tropical Medicine and Hygiene 86, 624–626.
Authie E (1994) Bovine trypanosomosis and trypanotolerance: a
role for ‘congopain’? Parasitology Today 10, 360–364.
Authie E, Cuisance D, Force-Barge P et al. (1991) Some new
prospects in epidemiology and fight against human African
trypanosomiasis. Research Revue of Parasitology 51, 29–46.
Biteau N, Bringaud F, Gibson WC, Truc P & Baltz T (2000)
Characterisation of Trypanozoon isolates using micro- and
minisatellite markers. Molecular and Biochemical Parasitology
105, 185–201.
Cattand P, Miezan TW & de Raadt P (1988) Human African
trypanosomiasis: use of double centrifugation of cerebrospinal
fluid to detect trypanosomes. Bulletin of the World Health
Organization 66, 83–86.
Cunningham I (1977) New culture medium for maintenance of
Tsetse tissues and growth of Trypanosomatids. Journal of
Protozoology 21, 325–329.
Dumas M & Bouteille B (1996) Human African trypanosomiasis.
Compte-Rendu de la Societe de Biologie 190, 395–408.
Garcia A, Cot M, Chippaux JP et al. (1998) Genetic control of
blood infection levels in human malaria: evidence for a complex
genetic model. American Journal of Tropical Medicine and
Hygiene 58, 480–488.
Garcia A, Jamonneau V, Magnus E et al. (2000) Longitudinal
survey of positive Card Agglutination Trypanosomiasis
Test (CATT) but apparently aparasitemic individuals in
Cote d’Ivoire: evidence for complex and heterogeneous popu-
lation. Tropical Medicine and International Health 5,
786–793.
Gashumba JK, Baker RD & Godfrey DG (1988) Trypanosoma
congolense: the distribution of enzymic variants in East and
West Africa. Parasitology 96, 475–486.
Gibson WC (1986) Will the real Trypanosoma brucei gambiense
please stand up. Parasitology Today 2, 255–257.
Gibson WC, Marshall TF & Godfrey DG (1980) Numerical
analysis of enzyme polymorphism: a new approach to the
epidemiology and taxonomy of trypanosomes of the subgenus
Trypanozoon. Advances in Parasitology 18, 175–246.
Ginoux PY & Frezil JL (1981) Recherches sur la latence clinique et
la trypanotolerance humaine dans le foyer du couloir du fleuve
Congo. Cahier ORSTOM, Serie Entomologie Medicale et
Parasitologie 1, 33–40.
Tropical Medicine and International Health volume 7 no 7 pp 610–621 july 2002
V. Jamonneau et al. Human African trypanosomiasis in Cote d’Ivoire (Ivory Coast)
ª 2002 Blackwell Science Ltd 619

Godfrey DG, Baker RD, Rickman LR & Mehlitz D (1990) The
distribution, relationships and identification of enzymatic var-
iants within the subgenus Trypanozoon. Advances in Parasit-
ology 29, 1–39.
Hide G, Cattand P, Le Ray D, Barry JD & Tait A (1990) The
identification of T. brucei subspecies using repetitive DNA
sequences. Molecular and Biochemical Parasitology 39, 213–226.
Jacquard A (1973) Distances genealogiques et distances genetiques.
Cahiers d’Anthropologie et d’Ecologie Humaine 1, 11–124.
Jamonneau V, Garcia A, N’Guessan P et al. (2000a) Clinical and
biological evolution of Human African Trypanosomiasis in Cote
d’Ivoire. Annals of Tropical Medicine and Parasitology 94, 831–
835.
Jamonneau V, N’Guessan P, N’Dri L, Simaro P & Truc P (2000b)
Exploration of the distribution of Trypanosoma brucei ssp. in
West Africa, by multilocus enzyme electrophoresis. Annals of
Tropical Medicine and Parasitology 94, 643–649.
Jannin J, Moulia-Pelat JP, Chanfreau B et al. (1993) Trypanos-
omiase Humaine Africaine: etude d’un score de presomption de
diagnostic au Congo. Bulletin of the World Health Organization
71, 215–222.
Lapeyssonnie L (1960) Deuxieme note concernant un cas exep-
tionnel de trypanosomiase, parasitemie observee pendant 21 ans
sans signes cliniques appreciables chez une malade traitee
inefficacement pendant les 10 premieres annees. Bulletin de la
Societe de Pathologie Exotique 57, 28–32.
Laurent JP, Barnabe C, Quesney V, Noel S & Tibayrenc M (1997)
Impact of clonal evolution on the biological diversity of
Trypanosoma cruzi. Parasitology 114, 213–218.
Lumsden WHR, Kimber CD, Evans DA & Doig SJ (1979)
Trypanosoma brucei: miniature anion-exchange centrifugation
technique for detection of low parasitemia; adapted for field use.
Transactions of the Royal Society of Tropical Medicine and
Hygiene 73, 312–317.
MacNamara JJ, Bailey J, Smith DH, Wakhooli S & Godfrey DG
(1995) Isolation of Trypanosoma brucei gambiense from Nor-
thern Uganda: evaluation of the Kit for in vitro isolation (KIVI)
in an epidemic focus. Transactions of the Royal Society of
Tropical Medicine and Hygiene 89, 388–389.
Magnus E, Vervoort T & Van Meirvenne N (1978) A card-
agglutination test with stained trypanosomes (CATT) for the
serological diagnosis of T. gambiense trypanosomiasis. Annales
de la Societe Belge de Medecine Tropicale 59, 169–176.
Mary C, Auriault V, Faugere B & Dessein AJ (1999) Control of
Leishmania infantum infection is associated with CD8(+) and
gamma interferon- and interleukin-5-producing CD4(+) antigen-
specific T cells. Infection and Immunity 67, 5559–5566.
Mehlitz D, Zillmann U, Scott CM & Godfrey DG (1982)
Epidemiological studies on the animal reservoir of gambiense
sleeping sickness. Part III. Characterisation of Trypanozoon
stocks by isoenzymes and sensitivity to human serum. Tropen-
medizin und Parasitologie 33, 113–118.
Miles MA, Cedillos RA, Povoa MM, Prata A, Sousa AA &
Macedo V (1981) Do radically dissimilar Trypanosoma cruzi
strains (zymodemes) cause Venezuelan and Brazilian forms of
Chagas’s disease? Lancet 20, 1338–1340.
Montamat EE, De Luca GM, Gallerano RH, Sosa R & Blanco A
(1996) Characterisation of Trypanosoma cruzi populations by
zymodemes: correlation with clinical pictures. American Journal
of Tropical Medicine and Hygiene 55, 625–628.
Murray M, Trail JCM & d’Ieteren GDM (1990) Trypanotolerance
in cattle and prospects for the control of trypanosomiasis by
selective breeding. Revue Scientifique et Technique de l’Office
Internationnal Des Epizooties 9, 369–386.
Oury B, Dutrait N, Bastrenta B & Tibayrenc M (1998) Trypan-
osoma cruzi: use of a synapomorphic RAPD fragment as a
species-specific DNA probe. Journal of Parasitology 83,
52–57.
Paindavoine P, Pays E, Laurent M et al. (1986) The use of DNA
hybridisation and taxonomy in determining the relationship
between Trypanosoma brucei stocks and subspecies. Parasitol-
ogy 92, 31–50.
Simo G, Njiokou F, Nkinin SW, Mgbedie M, Laveissiere C &
Herder S (2000) Etude de la prevalence des infections a
trypanosomes chez les animaux sauvages du foyer de la maladie
du sommeil de Bipindi, Cameroun. Bulletin de Liaison de
L’oceac 33, 8–15.
Smith DH & Bailey JW (1997) Human African trypanosomiasis in
South-eastern Uganda: clinical diversity and isoenzyme profiles.
Annals of Tropical Medicine and Parasitology 7, 851–856.
Sneath PHA & Sokal RR (1973) Numerical Taxonomy. The
Principles and Practice of Numerical Classification. W.H.
Freeman and Co, San Francisco, 573 pp.
Stevens JR, Nunes VLB, Lanham SM & Oshiro ET (1989)
Isoenzyme characterisation of Trypanosoma evansi isolated
from capybaras and dogs in Brazil. Acta Tropica 46, 213–222.
Stevens JR, Lanham SM, Allingham R & Gashumba JK (1992) A
simplified method for identifying subspecies and strain groups in
Trypanozoon by isoenzymes. Annals of Tropical Medicine and
Parasitology 86, 9–28.
Tibayrenc M, Neubauer K, Barnabe C, Guerrini F, Skarecky D &
Ayala FJ (1993) Genetic characterisation of six parasitic
protozoa: parity between random-primer DNA typing and
multilocus enzyme electrophoresis. Proceedings of the National
Academy of Sciences of the USA 90, 1335–1339.
Truc P, Mathieu Daude F & Tibayrenc M (1991) Multilocus
isoenzyme identification of Trypanosoma brucei stocks isolated
in Central Africa: evidence for an animal reservoir of sleeping
sickness in Congo. Acta Tropica 49, 127–135.
Truc P, Aerts D, McNamara JJ et al. (1992) The direct in vitro
isolation of Trypanosoma brucei from man and animals, and its
potential value for the diagnosis of Gambian trypanosomiasis.
Transactions of the Royal Society of Tropical Medicine and
Hygiene 86, 627–629.
Truc P, Formenty P, Diallo PB, Komoin-Oka C & Lauginie F
(1997a) Confirmation of two distincts classes of zymodemes of
Trypanosoma brucei infecting patients and wild mammals in
Cote d’Ivoire: suspected difference in pathogenicity. Annals of
Tropical Medicine and Parasitology 91, 951–956.
Truc P, Formenty P, Duvallet G, Komoin-Oka C, Diallo PB &
Lauginie F (1997b) Identification of trypanosomes isolated by
KIVI from wild mammals in Cote d’Ivoire: diagnostic, taxo-
Tropical Medicine and International Health volume 7 no 7 pp 610–621 july 2002
V. Jamonneau et al. Human African trypanosomiasis in Cote d’Ivoire (Ivory Coast)
620 ª 2002 Blackwell Science Ltd

nomic and epidemiological considerations. Acta Tropica 67,
187–196.
Truc P, Ravel S, Jamonneau V, N’Guessan P & Cuny G (2002)
Genetic variability within Trypanosoma brucei gambiense:
evidence of the circulation of different genotypes in Human
African Trypanosomiasis patients in Cote d’Ivoire. Transactions
of the Royal Society of Tropical Medicine and Hygiene 96,
1–4.
Truc P & Tibayrenc M (1993) Population genetics of Trypano-
soma brucei in Central Africa: taxonomic and epidemiological
significance. Parasitology 106, 137–149.
Waitumbi JN & Murphy NB (1993) Inter- and intra-species
differentiation of trypanosomes by genomic fingerprinting with
arbitrary primers. Molecular and Biochemical Parasitology 58,
181–186.
Welsh J & McClelland M (1990) Fingerprinting genomes using PCR
with arbitrary primers. Nucleic Acids Research 18, 7213–7218.
WHO (1998) Control and Surveillance of African Trypanosomi-
asis. WHO Technical Report Series 881, WHO, Geneva.
Williams JGK, Kubelik AR, Livak KJ, Rafalski JA & Tingley SV
(1990) DNA polymorphisms amplified by arbitrary primers are
useful as genetic markers. Nucleic Acids Research 18,
6531–6535.
Young CJ & Godfrey DG (1983) Enzyme polymorphism and the
distribution of Trypanosoma congolense isolates. Annals of
Tropical Medicine and Parasitology 77, 467–481.
Tropical Medicine and International Health volume 7 no 7 pp 610–621 july 2002
V. Jamonneau et al. Human African trypanosomiasis in Cote d’Ivoire (Ivory Coast)
ª 2002 Blackwell Science Ltd 621
Recommended