
Fax +41 61 306 12 34E-Mail [email protected]
Cytogenet Genome Res 2012;137:218–232 DOI: 10.1159/000341379
Low, Complex and Probably Reticulated Chromosome Evolution of Sciuromorpha (Rodentia) and Lagomorpha
F. Richard a, b B. Dutrillaux a
a Muséum National d’Histoire Naturelle, Département de Systématique et Evolution, Origine Structure et Evolution de la Biodiversité, Paris , and b Université de Versailles Saint Quentin en Yvelines, UFR de Sciences, Département de Biologie, Versailles , France
for an extensive period. Finally, among Lagomorpha and Sci-uromorpha, we focused on Leporidae and Sciuridae chro-mosome evolution. In the various attempts to establish di-chotomic evolutionary schemes, it was necessary to admit that multiple homoplasies (convergent and reverse rear-rangements) occurred in Sciuridae and in a lesser degree, in Leporidae. In Leporidae, additional rearrangements were sufficient to propose a resolved phylogeny. However, a re-solved phylogeny was not possible for Sciuridae because most of the rearrangements occurred in terminal branches. We conclude that a reticulated evolution took place early during the evolution of both families and lasted longer in Sciuridae than in Leporidae. In Sciuridae, most chromosome rearrangements were pericentric inversions involving short fragments. Such rearrangements have only mild meiotic consequences, which may explain the long persistence of the heterozygous status characterizing reticulated evolu-tion. Copyright © 2012 S. Karger AG, Basel
The orders Rodentia and Lagomorpha are grouped in the same taxon, the Glires superorder. They constitute a clade supported by morphological [Luckette and Harten-berger, 1993], chromosomal [Beklemisheva et al., 2011]
Key Words
Ancestral karyotypes � Chromosomes � Glires � Lagomorpha � Leporid � Reticulated evolution � Sciurid � Sciuromorpha
Abstract
Lagomorpha (rabbits and pikas) and Sciuromorpha (squir-rels) are grouped in the Glires superorder. Their chromo-some diversification, since their separation from the euthe-rian mammalian common ancestor, was characterized by a low rate of chromosome rearrangements. Consequently, the structure of some chromosomes was either conserved or only slightly modified, making their comparison easy at the genus, family and even order level. Interspecific in situ hy-bridization (Zoo-FISH) largely corroborates classical cyto-genetic data but provides much more reliability in com-parisons, especially for distant species. We reconstructed common ancestral karyotypes for Glires, Lagomorpha, Sci-uromorpha, and Sciuridae species, and then, determined the chromosome changes separating these ancestors from their common eutherian ancestor. We propose that reticulated evolution occurred during the diversification of Glires, which implies that several pericentric inversions and Robertsonian translocations were conserved in the heterozygous status
Published online: July 26, 2012
Florence Richard Muséum National d’Histoire Naturelle, Département de Systématique et EvolutionUMR7205 CNRS , Case postale 39, 16 Rue Buffon FR–75005 Paris (France) Tel. +33 140 795 341, E-Mail frichard @ mnhn.fr
© 2012 S. Karger AG, Basel1424–8581/12/1374–0218$38.00/0
Accessible online at:www.karger.com/cgr

Chromosome Evolution in Sciuromorpha and Lagomorpha
Cytogenet Genome Res 2012;137:218–232 219
and molecular data [Murphy et al., 2001a, b; Huchon et al., 2002; Prasad et al., 2008]. They shared a common an-cestor with Primates, Dermoptera and Scandentia, which groups them in the Euarchotonglires clade, as demon-strated by molecular data [Murphy et al., 2001a, b; Huchon et al, 2002; Douzery and Huchon, 2004].
Lagomorpha constitutes a small order, with less than 100 living species, grouped into 2 families, the Ochotoni-dae (pikas, 30 living species) with a single genus divided in 3 subgenera and Leporidae (hares and rabbits, 91 living species) with 11 genera [Hoffmann and Smith, 2005; Yu et al., 2000]. In contrast, Rodentia constitute by far the largest order of mammals. They comprise about 42% of the mammalian living species, with more than 2,277 liv-ing species [Musser and Carleton, 2005]. Lagomorpha share many morphological characters, and constitute a fairly homogenous group, whereas morphotypes are more heterogeneous in rodents, which were classified into 5 suborders (Sciuromorpha, Castorimorpha, Myo-morpha, Anomaluromorpha, and Hystrichomorpha). This difference suggests that evolution worked at very different speeds, low in Lagomorpha and fast in Roden-tia. However, the diversity of Rodentia does not exclude the possibility that, among this fast-evolving order, some taxa evolved slowly and conserved ancestral characters. Sciuromorpha, divided in 3 families (Sciuridae, Aplodon-tidae and Gliridae) and composed of only 307 species [Musser and Carleton, 2005] is considered as either the first diverging branch of Rodentia [Huchon et al., 2002] or sharing a basal position with Hystrichomorpha [Mont-gelard et al., 2008]. Moreover, in the 1970s and early 1980s, chromosome banding comparison showed that the garden dormouse Eliomys quercinus (Gliridae) and, above all, several squirrel species (Sciuridae) shared many chromosomal homoeologies with man and other pri-mates, whereas the karyotype of other rodent species, in particular that of Muridae, appeared to be very distinct [Dutrillaux et al., 1979; Petit et al., 1984]. Thus, among Rodentia, Sciuromorpha have conserved fairly ancestral chromosomes that evolved slowly compared to other ro-dents. Similar studies were performed in Lagomorpha, which showed that Leporids karyotypes were easy to compare with each other [Stock, 1976; Robinson, 1980; Robinson et al., 1984] and that many chromosomes of Oryctolagus cuniculus could be easily compared with those of man and other primates [Dutrillaux et al., 1980]. Thus, they are fairly conserved. Hence, Lagomorpha and Sciuromorpha, which maintained fairly ancestral char-acters, might have remained cytogenetically close to some slowly evolving mammals belonging to other or-
ders, favoring comparisons between orders and the re-construction of ancestral karyotypes at supraorder level.
With the development of molecular technologies, chromosome comparisons, by interspecific chromosome painting (Zoo-FISH), became easier and more precise, and the evolution of selected DNA sequences could be investigated with a great efficiency. In interspecific com-parisons, homologies could be proposed for almost all chromosomal segments. The high degree of conservation of the rabbit and squirrel chromosomes was confirmed by chromosome painting [Korstanje et al., 1999; Richard et al., 2003a; Stanyon et al., 2003]. Zoo-FISH is an invalu-able tool to perform chromosome comparisons between distant taxa of placental mammals. It makes it possible to reconstruct the karyotype of the eutherian ancestor with reasonable accuracy [Richard et al., 2003b; Ferguson-Smith and Trifonov, 2007; Robinson and Ruiz-Herrera, 2008] and the subsequent chromosome evolution which has led to extant species. In many groups, however, mul-tiple intrachromosomal changes, not detected by chro-mosome painting, have occurred. A complementary analysis by high-resolution chromosome banding and/or Zoo-FISH with DNA probes (such as BACs) is necessary to provide information on intrachromosomal rearrange-ments. Currently, there are Zoo-FISH and banding com-parisons of a large number of species, and it is now pos-sible to compare representative species of most system-atic groups with a great refinement.
Taking advantage of these data, we focus on the mech-anisms of chromosome evolution in the Glires and the implication of the different kinds of rearrangements. The construction of presumed ancestral karyotypes, at vari-ous levels, supraorder (Glires), order (Lagomorpha), sub-order (Sciuromorpha) and families (Sciuridae, Gliridae, Leporidae, and Ochotonidae), allowed us to propose which chromosome changes took place between these different stages and their implications. Then, we propose a chromosomal history for Leporidae and Sciuridae which are compared to the phylogenies obtained with molecular data. Finally, we propose that a reticulated evo-lution is more likely to have occurred in the beginning of the diversification of leporids and sciurids.
Diversity and Stability of Lagomorpha and
Sciuromorpha Karyotypes
The diploid numbers of these taxa are relatively stable ( table 1 ). Among Lagomorpha, it is more variable for Ochotonidae than Leporidae and among Sciuromorpha,
Richard /Dutrillaux
Cytogenet Genome Res 2012;137:218–232220
it is more stable for Sciuridae than Gliridae. The diploid number for Aplodontidae is known for only one species, Aplodontia rufa with 2n = 46 [Carrasco and Humphrey, 1968]. The diploid number of leporids varies from 42 to 52 chromosomes ( Pronolagus rupestris , Sylvilagus transi-
tionalis , respectively), while for Ochotonidae, the varia-tion is greater, from 40 to 68 ( Ochotona hyperborea, O. princeps , respectively) [Stock, 1976; Robinson, 1980; Rue-das et al., 1989; Romanenko et al., 2010]. There are differ-ent cytotypes for S. transitionalis with a diploid number
Table 1. List of Lagomorpha and Sciuromorpha species used for this analysis
Species Code 2n Number of auto-somal segments conserved
Karyotype references Zoo-FISH references
LagomorphaLeporidaeBrachylagus idahoensis BID 44 38 Robinson et al., 1984 Robinson et al., 2002 (OCU)Bunolagus monticularis BMO 44 38 Robinson and Skinner, 1983 Robinson et al., 2002 (OCU)Lepus europaeus LEU 48 38 Richard et al., 2003bLepus saxatilis LSA 48 38 Robinson, 1980 Robinson et al., 2002 (OCU)Oryctolagus cuniculus OCU 44 38 Committee for standardized karyotypes, 1981 Korstanje et al., 1999 (HSA); Chantry-
Darmon et al., 2005Pronolagus rupestris PRU 42 38 Robinson, 1980 Robinson et al., 2002 (OCU)Romerolagus diazi RDI 48 38 Robinson et al., 1981 Robinson et al., 2002 (OCU)Sylvilagus floridanus SFL 42 38 Robinson et al., 1983 Robinson et al., 2002 (OCU)OchotonidaeOchotona hyperborea OHY 40 39 Romanenko et al., 2010 Romanenko et al., 2010 (HSA)Ochotona forresti OFO 54 41 Ye et al., 2011 Ye et al., 2011 (HSA)
SciuromorphaGliridaeEliomys quercinus EQU 48–54 45 Dutrillaux et al., 1979Eliomys munbyanus EMU 46 45 Filippucci et al., 1988b Sannier et al., 2011 (HSA)Eliomys melanurus EME 46–48 45 Filippucci et al., 1988a; Sannier et al., 2011SciuridaeAtlantoxerus getulus AGE 38 35 Petit et al., 1984Xerus cf. erythropus XER 38 35 Li et al., 2006 Li et al., 2006 (SCA, HSA)Marmota baibacina MBA 38 36 Beklemisheva et al., 2011 Beklemisheva et al., 2011 (TSI)Marmota himalayana MHI 38 36 Li et al., 2006 Li et al., 2006 (SCA, HSA)Marmota katschenkoi MKA 36 36 Beklemisheva et al., 2011 Beklemisheva et al., 2011 (TSI)Marmota monax MMO 38 36 Petit et al., 1984Tamias sibiricus TSI 38 35 Petit et al., 1984 Li et al., 2004 (HSA, SCA)
Tamias striatus TST 38 35 Nadler and Block, 1962; this reviewSpermophilus erythrogenis
SER 36 35 Beklemisheva et al., 2011 Beklemisheva et al., 2011 (TSI)
Spermophilus major SMA 36 35 Beklemisheva et al., 2011 Beklemisheva et al., 2011 (TSI)Spermophilus undulatus SUN 32 35 Beklemisheva et al., 2011 Beklemisheva et al., 2011 (TSI)Spermophilus suslicus SSU 34 35 Beklemisheva et al., 2011 Beklemisheva et al., 2011 (TSI)Heliosciurus gambianus HGA 40 35 Petit et al., 1984Callosciurus erythraeus CER 40 35 Petit et al., 1984 Li et al., 2004 (SCA)Callosciurus finlaysonii CFI 40 35 Sannier et al., 2011Tamiops rodolphii TRO 38 35 Sannier et al., 2011Tamiops maritimus TMA 38 35 Oshida et al., 2002
Dremomys rufigensis DRU 38 35 Nadler and Hoffmann, 1970, this reviewMenetes berdomrei MBE 38 35 Sannier et al., 2011 Richard et al., 2003a (HSA)Sciurus vulgaris SVU 40 35 Petit et al., 1984 Beklemisheva et al., 2011 (TSI)Sciurus carolinensis SCA 40 38 Nadler and Sutton, 1967 Stanyon et al., 2003; Li et al., 2004 (HSA)
Hylopetes cf pharey HPH 38 35 this reviewPetaurista albiventer PAL 38 36 Li et al., 2004 Li et al., 2004 (HSA, SCA)
2 n = Diploid number; HSA = human; OCU = rabbit; TSI = chipmunk; SCA = grey squirrel.
The autosomal segments homologous to human chromosomes con-served in each species are directly obtained by Zoo-FISH data or deduced after chromosome banding comparisons with painted species. Hylopetes cf
pharey: the attribution of H. pharey is provisional because it may be con-fused with H. alboniger. In bold type: species whose karyotypes are shown in figure 1. For M. himalayana, the number of autosomal segments con-served is 36 instead of 35; see Li et al. [2006] and comments about HSA12 homologies in Beklemisheva et al. [2011].

Chromosome Evolution in Sciuromorpha and Lagomorpha
Cytogenet Genome Res 2012;137:218–232 221
of 46 or 52 [Robinson et al., 1983; Ruedas et al., 1989]. For Sciurids, the diploid number of 38 is very common ( fig. 1 ),except in some genera (Marmota, Spermophilus, Helio-sciurus, Callosciurus and Sciurus) in which small varia-tions exist ( table 1 ). For Spermophilus , the variation is from 32 (S. undulatus) to 42 (S. xanthroprymnus) , for Marmota, 36 to 42, and 40 for Sciurus, Callosciurus and Heliosciurus genera [Nadler and Hoffmann, 1970; Nadler et al., 1975; Petit et al., 1984; Li et al., 2004, 2006; Arslan, 2005; Beklemisheva et al., 2011]. In contrast, Gliridae have a diploid number varying from 46 (E. munbyanus) to 62 (Glis glis) . There are different cytotypes for some Eliomys species ( table 1 ) with a diploid number varying from 48 to 54 (E. quercinus) or 46–48 (E. melanurus) , fol-lowing Robertsonian fusion or fission events [Zima et al., 1995; Sannier et al., 2011].
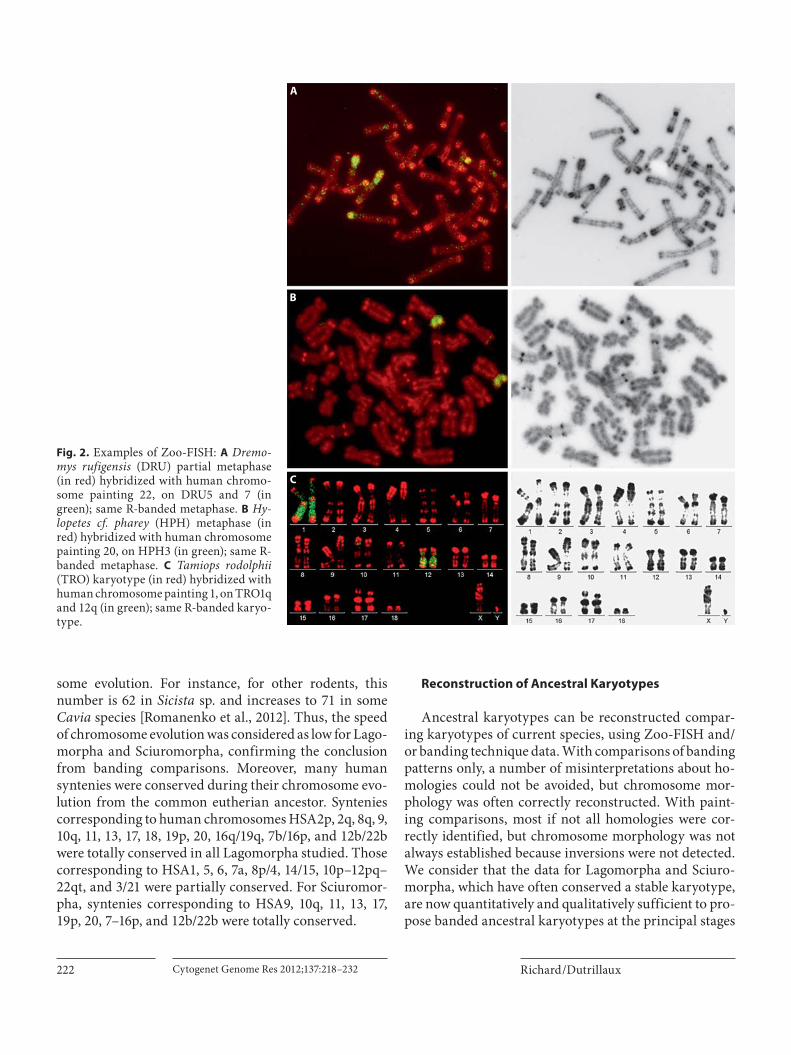
The successful comparison on the basis of chromo-some banding of some leporid karyotypes with each oth-er and with primates demonstrated that leporids have not experienced rapid chromosome evolution [Stock, 1976; Dutrillaux et al., 1980; Robinson, 1980; Robinson et al., 1984]. The same conclusion was made for Sciuridae and, to a lesser extent, for Gliridae [Dutrillaux et al., 1979;Petit et al., 1984; Petit and Dutrillaux, 1985]. Zoo-FISH using human chromosome probes performed in some Lagomorpha and Sciuromorpha species identified chro-mosomal segments conserved among species with un-known gene order. Thus, using the chromosomal homol-ogies deduced between human and each species, their chromosomes could be directly compared ( fig. 2 ). Zoo-FISH was also performed using flow-sorted rodent chro-mosomes from Sciurus carolinensis , Tamias sibiricus and Castor fiber , and O. cuniculus sorted chromosomes [Rob-inson et al., 2002; Stanyon et al., 2003; Li et al., 2004, 2006; Beklemisheva et al., 2011]. The number of autosomal seg-ments conserved between human and each species could be directly calculated or deduced from Zoo-FISH ( ta-ble 1 ). It varied from 35 to 45 human segments conserved. This number is informative about the speed of chromo-
A
B
C
Fig. 1. A RBG-banded karyotype of Hylopetes cf. pharey , with2n = 38,XX, showing 15 pairs of submetacentric and 3 pairs of metacentrics (HPH9, 14 and 15) autosomes. The p arm of chromo-somes 13, 17 and 18 is completely heterochromatic. B RBG-banded karyotype of Dremomys rufigensis , with 2n = 38,XY, showing 14 submetacentric, 3 metacentric (DRU9, 14, 15) and 1 acrocentric (DRU18) autosomes. Heterochromatin is observed at the end of the p-arm of chromosomes 13, 14, 16 and 17, and in an intercalary position in the p-arm of chromosome 8. C RBG-banded karyotype
of Tamias striatus , with 2n = 38,XX showing 13 submetacentric, 4 acrocentric (TST1, 11, 17 and 18) and 1 submetacentric (TST9). Chromosomes are classified according to Callosciurus sp . karyo-type in Sannier et al. [2011], and heterochromatic arms were not taken into consideration for chromosome orientation. h = Hetero-chromatin. Dots show the positions of the centromeres. The small differences in size or banding patterns observed sometimes be-tween the 2 homologs of a pair are owed to variation in the rate of BrdU incorporation during the S phase of the cells in culture.
Richard /Dutrillaux
Cytogenet Genome Res 2012;137:218–232222
some evolution. For instance, for other rodents, this number is 62 in Sicista sp . and increases to 71 in some Cavia species [Romanenko et al., 2012]. Thus, the speed of chromosome evolution was considered as low for Lago-morpha and Sciuromorpha, confirming the conclusion from banding comparisons. Moreover, many human syntenies were conserved during their chromosome evo-lution from the common eutherian ancestor. Syntenies corresponding to human chromosomes HSA2p, 2q, 8q, 9, 10q, 11, 13, 17, 18, 19p, 20, 16q/19q, 7b/16p, and 12b/22b were totally conserved in all Lagomorpha studied. Those corresponding to HSA1, 5, 6, 7a, 8p/4, 14/15, 10p–12pq–22qt, and 3/21 were partially conserved. For Sciuromor-pha, syntenies corresponding to HSA9, 10q, 11, 13, 17, 19p, 20, 7–16p, and 12b/22b were totally conserved.
Reconstruction of Ancestral Karyotypes
Ancestral karyotypes can be reconstructed compar-ing karyotypes of current species, using Zoo-FISH and/or banding technique data. With comparisons of banding patterns only, a number of misinterpretations about ho-mologies could not be avoided, but chromosome mor-phology was often correctly reconstructed. With paint-ing comparisons, most if not all homologies were cor-rectly identified, but chromosome morphology was not always established because inversions were not detected. We consider that the data for Lagomorpha and Sciuro-morpha, which have often conserved a stable karyotype, are now quantitatively and qualitatively sufficient to pro-pose banded ancestral karyotypes at the principal stages
A
B
C
Fig. 2. Examples of Zoo-FISH: A Dremo-mys rufigensis (DRU) partial metaphase (in red) hybridized with human chromo-some painting 22, on DRU5 and 7 (in green); same R-banded metaphase. B Hy-lopetes cf. pharey (HPH) metaphase (in red) hybridized with human chromosome painting 20, on HPH3 (in green); same R-banded metaphase. C Tamiops rodolphii (TRO) karyotype (in red) hybridized with human chromosome painting 1, on TRO1q and 12q (in green); same R-banded karyo-type.

Chromosome Evolution in Sciuromorpha and Lagomorpha
Cytogenet Genome Res 2012;137:218–232 223
of Glires evolution, starting from the ancestral eutherian karyotype.
For these reconstructions, human chromosome ho-mologies were obtained from painting data and deduced by banding comparisons for species not painted. We have compared the species studied with the eutherian ances-tral karyotype (EUT) [Dobigny et al., 2004] and consid-ered as ancestral (symplesiomorphic) the chromosomal segments with unknown gene order shared by all these species ( table 1 ). The EUT karyotype, with 2n = 46, was proposed by Sannier et al. [2011]. It is a consensual recon-struction with other authors [Murphy et al., 2001b; Rich-ard et al., 2003b; Ferguson-Smith and Trifonov, 2007;
Robinson and Ruiz-Herrera, 2008], except for the homol-ogous to HSA19p which remains an independent acro-centric chromosome as described in Ruiz-Herrera et al. [2012]. It is shown by R-banded chromosomes in figure 3 . Then, we have inferred the position of the centromere as submetacentric if this shape is shared by taxa not direct-ly related. The acrocentric shape is chosen if it is observed in taxa not directly related and/or if there was no com-mon submetacentric shape. With data from the literature and some new karyotype comparisons, we have recon-structed the R-banded karyotypes of the ancestor of Gli-res, Lagomorpha, Sciuromorpha and Sciuridae, using chromosomes of present species from these clades.
Fig. 3. Eutherian ancestral R-banded karyotype (EUT) reconstructed according to Richard et al. [2003b]with modifica-tions according to Ferguson-Smith and Trifonov [2007] and Sannier et al. [2011], with 2n = 46. Chromosomes belong to Carnivora (P. hermaphroditus) , Lagomor-pha (L. europaeus) , Primates (M. murinus and D. madagariensis) , and Xenarthra (T. chinensis) . The banding pattern of EUT1 remains uncertain. The centromeric posi-tion is not well defined for EUT4. The slight differences observed between the 2 homologs of EUT8 come from different amounts of pericentromeric heterochro-matin. Beside each pair, human homolo-gies are indicated. Dots show the positions of the centromeres.
Fig. 4. Glires ancestral R-banded karyo-type (GLI), with 2n = 46–50. Chromo-somes belong to Lagomorpha or Sciuridae karyotypes. Pair 4 was either homozy-gously acrocentric or submetacentric, or heterozygous (see text for details). GLI1 and 5 were homozygously fused or not, or heterozygous (see text for details). The slight differences observed between the 2 homologs of GLI8 come from different amounts of pericentromeric heterochro-matin. Beside each chromosome pair, hu-man homologies are indicated. Dots show the positions of the centromeres.

Richard /Dutrillaux
Cytogenet Genome Res 2012;137:218–232224
The Glires ancestor karyotype (GLI) is composed of 2n = 46–50 chromosomes ( fig. 4 ). Almost all the euthe-rian syntenies were conserved in this karyotype: the chromosomes homologous to human chromosomes HSA4–8p, 5, 6, 16q–19q, 7b–16p, 3–21, 14–15, 12qt–22q, 2qt, 8q, 7a, 10q, 13, 2pq, 17, 18, 20, 19p, and X. The syn-teny was disrupted for EUT1 (homologous to HSA1) and EUT5 (homologous to HSA10p–12pq–22qt), and new syntenies have occurred: HSA9–11 (GLI1) and HSA10p–1qt (GLI5). We could not decide on the ancestral shape for these 2 chromosomes. For HSA10p and 1qt, Sciuridae and Lagomorpha, but not Gliridae, displayed the fused form; for HSA9 and 11, Sciuridae, except the Sciurus ge-nus, rabbit and one pika (O. hyperborea) , displayed the fused form, while Gliridae and most of the Lagomorpha species have 2 acrocentrics. The centromere position was not well defined for the ancestral chromosome homolo-gous to HSA6: a submetacentric form was common to Sciuridae and Leporidae, but not to Gliridae, and one pika has an acrocentric form. A GLI composed of 2n = 50 was proposed by Beklemisheva et al. [2011]. They consid-ered 2 more disruptions, the 8p–4 into 4a and 4b–8p and the 3–21 into 3a–21 and 3b. However, the reassessment of the data of Beklemisheva et al. [2011], who used chip-munk (T. sibiricus) chromosome paintings on rabbit (O. cuniculus) metaphases, indicated that the breakpoints of the fissions differ in Lagomorpha and Sciuromorpha. Thus, they are not inherited from their common ances-tor.
The reconstruction of the Lagomorpha ancestor karyotype (LAG) was proposed by Beklemisheva et al. [2011] and Ye et al. [2011] with 2n = 56 or 60, respectively. We propose a LAG of 2n = 58–60 chromosomes ( fig. 5 ). The ancestral Glires syntenies are conserved, except for GLI2 (HSA4–8p), GLI3 (HSA5), GLI8 (HSA3–21), GLI9 (HSA14–15), GLI11 (HSA12pq–22qt), and GLI14 (HSA7a). Six fission events have occurred for these chromosomes and 1 inversion for the homologous to HSA14–15. The synteny 10p–1qt (GLI7) is fixed in this ancestral karyo-type. The centromeric position remains ambiguous for chromosome homologous to HSA1pq because submeta-centric and acrocentric forms are described for this chro-mosome in various taxa. The lower diploid number in Beklemisheva et al. [2011], compared to our reconstruct-ed Lagomorpha ancestral karyotype, lies in the presence of a small acrocentric chromosome homologous to HSA14b and observed in all the Lagomorpha karyotypes, not considered by these authors. For HSA9 and 11, Ye et al. [2011] proposed the acrocentric forms as ancestral whereas Beklemisheva et al. [2011] preferred the fusion into a single metacentric chromosome. For instance, the rabbit and one pika (O. hyperborea) display the fused form, and most of the Lagomorpha have the acrocentrics. Thus, we preferred to make no formal choice and con-served the 2 possibilities in our figure.
No reconstruction of the Sciuromorpha ancestral karyotype was proposed in the literature, but Beklemi-sheva et al. [2011] proposed that the Sciuridae ancestral
Fig. 5. Lagomorpha ancestral R-banded karyotype (LAG), with 2n = 58–60. Chro-mosomes belong to Lagomorpha (L. euro-paeus and O. cuniculus) karyotypes. Pairs 1, 3 and 4 were either homozygous for A or B, or heterozygous (see text for details). Be-side each chromosome pair, human ho-mologies are indicated. Dots show the po-sitions of the centromeres.

Chromosome Evolution in Sciuromorpha and Lagomorpha
Cytogenet Genome Res 2012;137:218–232 225
karyotype might be similar to that of the Sciuromorpha (SCI). However, they discarded the Gliridae family which is important for the reconstruction of the SCI karyotype [Sannier et al., 2011]. We propose that the SCI ancestor karyotype is composed of 2n = 48–52 chromosomes ( fig. 6 ). The ancestral Glires syntenies are conserved ex-cept for GLI8 (HSA3–21) and GLI2 (HSA4–8p). For 6 SCI chromosomes, some doubts remain (SCI1, 3, 5, 6, 7, and 8). One fission event, different from that in LAG [Bek-lemisheva et al., 2011], occurred for the homolog to GLI2 (HSA4–8p), followed by an inversion of the 4a–8p seg-ment and a fusion to the GLI11 (HSA12pq–22qt) leading to SCI2. Two fissions different from those of LAG [Bek-lemisheva et al., 2011] occurred for GLI8 (HSA3–21) leading to 3 acrocentrics or, alternately, to 2 acrocentrics (SCI22 and 24) and 1 submetacentric (SCI5). Uncertain-ty remains about the centromeric position of SCI5 be-cause SCI5 corresponds to an acrocentric form in the genera Marmota and Spermophilus and different sub-metacentric forms for the other Sciuridae and Gliridae. For SCI1, homologs to HSA9 and 11 are fused in a meta-centric chromosome in Sciuridae except Sciurus sp ., and not in Gliridae. For SCI6, homologs to HSA1q and 10p are fused in a submetacentric chromosome in Sciuridae, but not in Gliridae. Therefore, for SCI1 and 6, we cannot decide if the fused form is ancestral or not. For SCI3 (HSA5), submetacentric and acrocentric chromosomes are observed in Sciurids. SCI7, homolog to HSA2pq, has a submetacentric form similar in Gliridae and in some Sciuridae (A. getulus, Callosciurus sp . ) , an acrocentric form in other Sciuridae (Microphyla berdmorei, Tamias
sp., Hylopetes sp . ) and other submetacentric forms in the remaining Sciuridae. SCI8, homolog to HSA18, is sub-metacentric in Gliridae and a large part of the Sciuridae, and acrocentric in some other Sciuridae (M. berdmorei, Sciurus sp., Petaurista sp . ) . Hence, the centromeric posi-tion for SCI3, 7 and 8 chromosomes remains uncertain, and we maintain both possible forms, acrocentric or sub-metacentric.
The Sciuridae ancestral karyotype (SCIU) was de-scribed by Graphodatsky et al. [2008] and Beklemisheva et al. [2011] with a diploid number of 2n = 38. We pro-posed a SCIU with 2n = 38–40 ( fig. 7 ). In contrast to these authors, we concluded that SCI14 and 19 were fused in the ancestral form, but we think that data is in-sufficient to decide on the status of these chromosomes. The fused form does not exist in Callosciurus, Heliosci-urus, Marmota , and Spermophilus genera. So, we main-tain the 2 possibilities, fused and non-fused, in the Sciu-ridae ancestral karyotype. This reconstructed ancestral karyotype differs from that of Sciuromorpha by 6 fu-sions specific to the Sciuridae, SCI13+15, SCI14+19, SCI11+20, SCI16+21, SCI17+18, and SCI23+24. SCI6 was fixed in its fused form. The centromeric position for SCIU6, 10, 12, and 13 remain uncertain (see above for explanations).
The Gliridae ancestral karyotype was described in Sannier et al. [2011]. It differs from that of Sciuromorpha by 8 fissions, 10 fusions and 14 inversions. We propose a supplementary inversion of the segment of HSA3 fused with HSA9 according to the Sannier et al. [2011] nomen-clature.
Fig. 6. Sciuromorpha ancestral R-banded karyotype (SCI), with 2n = 50–54. Chro-mosomes belong to Sciuridae or Gliridae (M. berdmorei, T. rodolphii, M. monax and E. melanurus) karyotypes. For SCI3, 5, 7 and 8, two or three chromosomes could represent these ancestral forms, an acro-centric and 1 or 2 different submetacentric ones (see text for details). Pairs 1 and 6 were homozygously fused or not, or het-erozygous (see text for details). Beside each chromosome pair, human homologies are indicated. Dots show the positions of the centromeres.

Richard /Dutrillaux
Cytogenet Genome Res 2012;137:218–232226
The Leporidae and Ochotonidae ancestral karyotypes are very close to that of Lagomorpha. Two fusions sepa-rate the Leporidae from the Lagomorpha ancestral karyo-type, and 1 inversion and 2 fissions separate the Ochoto-nidae from the LAG ( fig. 8 ).
Chromosome Evolution since the Eutherian
Ancestor
The chromosome rearrangements that have occurred between the eutherian ancestor and the GLI, LAG, SCI, SCIU, Gliridae, Ochotonidae, and Leporidae ancestors are shown in figure 8 . The rate of rearrangements was very low during the first steps of Glires diversification and in the lineages leading to Ochotinidae, Leporidae and Sciuridae. For instance, about 0.3–0.5 rearrange-ment per million years would have occurred in the lin-eages leading to the ancestors of Lagomorpha and Sci-uromorpha. The rearrangement rate increased 10-fold in the Gliridae lineage (about 3 rearrangements per million years). This high rate is equivalent to that of the Muridae lineage [Stanyon et al., 1999]. Thus, the clock of chromo-some rearrangements varied in the different lineages. It was low in the lineage leading to Glires, Lagomorpha,Sciuromorpha, Ochotonidae, Leporidae, and Sciuridae ancestors, in contrast to the high rate observed in the lineage leading to the Gliridae ancestor. The types of re-arrangements that occurred in the branches leading to Lagomorpha and Sciuromorpha ancestors were differ-ent, with fissions in the LAG branch and a complex rear-
rangement in the SCI branch (involving 1 fission fol-lowed by 1 pericentric inversion and 1 Robertsonian translocation). Few rearrangements occurred during the diversification of Lagomorpha: 2 additional fissions oc-curred in the branch leading to the Ochotonidae ances-tor, whereas 2 Robertsonian translocations took place along the Leporidae branch. During the diversification of Sciuromorpha, the numbers and types of rearrange-ments were different in the Gliridae and Sciuridae branches. Robertsonian translocations accumulated in the Sciuridae branch, while many fissions, pericentric inversions and complex or Robertsonian translocations occurred in the Gliridae lineage [Sannier et al., 2011]. The time of occurrence for some rearrangements re-mains uncertain (indicated in white and grey in fig. 8 ). They might have been present in a homozygous state in the ancestor, but multiple reversions would later then be required in the chromosome evolution of these taxa (not shown in fig. 8 ). A second hypothesis is that they were heterozygous in ancestral populations. Then, the rear-rangements colored in grey would correspond to a pro-gression toward the homozygous state in various phylo-genetic lines. This is the interpretation chosen in figure 8 . The presence of heterozygosity for some inversions and Robertsonian translocations is proposed during the basal steps of the Glires diversification, which could re-sult from hybridization events between homozygous populations (introgression) or conservation of heterozy-gous individuals, leading to a reticulated evolution [Chang et al., 2011; Liu et al., 2011].
Fig. 7. Sciuridae ancestral R-banded karyotype (SCIU), with 2n = 38–40. Chro-mosomes belong to Sciuridae (M. berd-morei, T. rodolphii and M. monax) karyo-types. Pairs 6, 10, 12 and 13 were homozy-gously acrocentric or submetacentric, or heterozygous (see text for details). Chro-mosome 4 was homozygously fused or not, or heterozygous (see text for details). Be-side each chromosome pair, human ho-mologies are indicated. Dots show the po-sitions of the centromeres.

Chromosome Evolution in Sciuromorpha and Lagomorpha
Cytogenet Genome Res 2012;137:218–232 227
Chromosome Evolution in Ochotonidae, Leporidae
and Sciuridae
In the Ochotonidae family, 4 karyological groups have been proposed [Vorontsov and Ivanitskaya, 1973; Stock, 1976; Capanna et al., 1991]. However, only 2 species have been compared using Zoo-FISH [Romanenko et al., 2010; Ye et al., 2011] which makes it hazardous to reconstruct chromosome relationships between the species of the
family. Molecular phylogenies confirm the 2nd, 3rd and 4th karyological groups including O. macrotis , O. rutila , O. roylei (2n = 62) and O. rufescens (2n = 60) in the 2nd group, O. daurica in the 3rd, and O. hyperborea (2n = 40), O. alpina (2n = 42) and O. pallasi (2n = 38) in the 4th group. In contrast, the first karyological group including O. collaris , O. princeps and O. pusilla (2n = 68) is not sup-ported by molecular data [Capanna et al., 1991; Yu et al., 2000; Niu et al., 2004].
Fig. 8. Chromosome evolution of Glires since the eutherian (EUT) ancestor with the most parsimonious reconstruction of the rearrangements separating the differ-ent ancestral karyotypes. GLI = Glires, LAG = Lagomorpha, SCI = Sciuromorpha. Types of rearrangements: 1 = Arrow: fis-sion, 2 = white square: fusion, 3 = white circle: pericentric inversion, 4 = white and grey circle and 5 = white and grey square: pericentric inversion and fusion present in a heterozygous state, respectively (see text for details). Grey symbols: acquisition of the homozygousity from a previous het-erozygous state for pericentric inversions (6 = circle), and fusions (7 = square) (see text for details). On the left of the rear-rangements, EUT, GLI, SCI or LAG chro-mosomes involved and on the right, hu-man homologies. a, b, a’, b’ and c’: seg-ments of ancestral chromosomes which were fissioned during their evolution. Pre-sumed times of divergence are indicated in million years (My) [Huchon et al., 2002; Montgelard et al., 2002, 2008].

Richard /Dutrillaux
Cytogenet Genome Res 2012;137:218–232228
During Leporidae diversification, Robertsonian fu-sions occurred frequently [Robinson, 1980; Robinson and Skinner, 1983; Robinson et al., 1983, 1984, 2002; Ruedas et al., 1989]. A resolved chromosomal phylogeny, recon-structed from species studied with Zoo-FISH [Korstanje et al., 1999; Robinson et al., 2002; Chantry-Darmon et al., 2005] is shown in figure 9 . The karyotype of the Lagomor-pha ancestor ( fig. 5 ) corresponds to the ancestral condi-tions. The grey symbols correspond to either homoplasies shared with Ochotonidae or acquisition of a homozygous status. Two branches diverge directly from the Leporidae ancestor: one leading to Bunolagus monticularis and the 2nd to the other leporid genera. B. monticularis has a very derived karyotype obtained by 6 rearrangements specific to this lineage and 3 acquisitions of a homozygous state or homoplastic rearrangements. A common trunk of 4 spe-cific rearrangements and 2 acquisitions of a homozygous state or homoplastic rearrangements leads to the other leporid genera. This situation differs from Robinson et al.
[2002] who considered the Lepus karyotype as the closest to the Leporidae ancestor and proposed that 4 fission events occurred in the Bunolagus lineage. After this com-mon trunk, few rearrangements occurred. The 1st diverg-ing branch leads to the rabbit and Brachylagus idahoensis , which share 1 Robertsonian fusion. The 2nd branch leads to Romerolagus , Lepus , Sylvilagus , and Pronolagus . L. sax-atilis and R. diazi do not acquire any other rearrangement and seem to share the same karyotype, according to Rob-inson et al. [2002]. One Robertsonian fusion is shared by Pronolagus rupestris and Sylvilagus floridanus . In the Syl-vilagus genus, the diploid number varies mainly following the occurrence of different Robertsonian translocations [data not shown, Ruedas et al., 1989]. Some Sylvilagus sp ., such as S. bachmani, which has a karyotype very similar to that of Lepus , did not share the Robertsonian fusionLAG1A+16 [Stock, 1976]. This suggests that either thisrearrangement is a convergent event or a population, het-erozygous for this rearrangement, has remained in this
Fig. 9. Chromosomal phylogeny of Leporidae. White circle = pericentric inversion; white square = Robertsonian translocation; black square = WART. Grey symbols represent the acquisition of the homozygousity from a pre-vious heterozygous state or homoplasies for pericentric inversions (grey circle) and Robertsonian translocations (grey square) (see text for details). On the left of the rearrangements, LAG chromosomes involved and on the right, human homologies.

Chromosome Evolution in Sciuromorpha and Lagomorpha
Cytogenet Genome Res 2012;137:218–232 229
lineage. Surprisingly, the position of Pronolagus , as sister group of Sylvilagus , is not supported by molecular data, which considered it as the most basal branch of Leporidae [Kriegs et al., 2010]. However, this interpretation is unac-ceptable because it would require us to ignore about 14 non-homoplastic translocations which separate Sylvila-gus - Pronolagus from Bunolagus karyotypes ( fig. 9 ). More-over, the intergeneric relationships deduced from chro-mosomal data are not supported by molecular data, but the molecular results remain controversial [Halanych and Robinson, 1997, 1999; Matthee et al., 2004]. If both chro-mosomal and molecular data were robust, these discrep-ancies would imply that a quite unusual number of homo-
plasies occurred either at the DNA or at the chromosome level. Finally, the absence of a correlation between the lep-orid geographical distribution over the world and their genetic (chromosomal and molecular) relationships sug-gests that either their origin is older than thought or that numerous dispersal events might have occurred during their diversification [Robinson et al., 2002; Matthee et al., 2004]. Whatever the case, their recent evolution has in-volved few chromosome rearrangements.
Sciuridae chromosome evolution is mainly composed of pericentric inversions and some Robertsonian fusions. It is also accompanied by the addition of constitutive het-erochromatin in some lineages (for instance: DRU, HPH,
Fig. 10. Positioning of the chromosome rearrangements having occurred in Sciuridae, in a published molecular tree [Mercer and Roth, 2003]. Species are named by a 3-letter code (cf. table 1). Circle = inversion; square = Robertsonian translocation; dark square = WART; dark rectangle = complex rearrangement; crossed rearrangement = reversion event. Grey symbols represent the acquisition of the homozygosity from a previous heterozygous
state for pericentric inversions (grey circle) and Robertsonian translocations (grey square) (see text for details). Numbers over the rearrangements: SCIU chromosomes involved; ac = acrocen-tric; sb = submetacentric; fi = fission; fu = fusion; a–d = different rearrangements involving the same chromosome; 17 = rearrange-ment of SCIU4, involving the homolog to HSA17.

Richard /Dutrillaux
Cytogenet Genome Res 2012;137:218–232230
SCA, Spermophilus lineages), not considered in figure 10 . Forty-five rearrangements are deduced in Sciuridae evo-lution since their common ancestor SCIU, including 20 autapomorphies (observed only in 1 species). The re-maining rearrangements are diversely shared and put to-gether genera which are not closely related according to the systematic classification or molecular phylogenies [Mercer and Roth, 2003; Herron et al., 2004; Steppan et al., 2004; Musser and Carleton, 2005]. As the status of SCIU4, 6, 10, 12, and 13 chromosomes is imprecise ( fig. 7 ), the 5 rearrangements involving these chromosomes (4 pericentric inversions and 1 Robertsonian fission or fu-sion) could not be polarized. But, whatever ancestral form chosen, multiple homoplasies would need to have occurred (not shown). An alternative hypothesis is that these chromosomes conserved a heterozygous status for many generations before becoming fixed in a homozy-gous state. This situation makes it impossible to construct a resolved phylogenetic tree based on chromosome rear-rangements, in accordance with the systematics: a fast radiation within Sciuridae family coupled to introgres-sion would have eliminated cytogenetic footprints. To il-lustrate this difficulty, we placed the chromosome rear-rangements on a molecular tree [Mercer and Roth, 2003]: obviously, they do not support the basal nodes ( fig. 10 ). The 5 rearrangements involving SCIU4, 6, 10, 12, and 13 chromosomes are homoplastic, whatever the SCIU forms chosen. Thus, the hypothesis of a long persistence of their heterozygous status until they became fixed in a homo-zygous state is preferred for these characters (grey rear-rangements in fig. 10 ). This conclusion means that sev-eral introgression events might have occurred leading to a reticulated evolution during the emergence of various genera [Nosil, 2008; Liu et al., 2011]. The long persistence of heterozygosity implies that these chromosome rear-rangements were probably permissive, i.e. without strong
effects on meiosis and reproduction. Interestingly, this is probably the case for the 4 inversions which involve only short pericentromeric segments [Dutrillaux et al., 1986; Morel et al., 2007]. For the remaining rearrangements,20 are autapomorphies and 20 are informative with 13 shared at the genus level (4 for the Sciurus , 6 for the Tamias , and 3 for Spermophilus , except SUN). Indeed, numerous rearrangements were accumulated at the edge of the branches, leading to genera or species. For instance, inversions occurred in the branch leading to Sciurus , Tamias or Hylopetes genus, Robertsonian translocations and inversions in the branch common to Marmota and Spermophilus genera, and WART in the Spermophilus lineage. Finally, the chromosome evolution remained very low for the genera Callosciurus, Heliosciurus, Dre-momys, Tamiops, Atlantoxerus , and Xerus .
In conclusion, Leporidae and Sciuridae chromosome evolution are both characterized by an early period of a reticulated evolution with conservation of heterozygosity for several chromosomes. This ‘hybrid’ period was cer-tainly longer for Sciuridae than Leporidae, and it was coupled with a fast radiation within Sciuridae family, eliminating cytogenetic footprints. Then, rearrange-ments more recently accumulated in some lineages ofSciuridae genera, while the karyotypes of Leporidae re-main fairly stable. Thus providing another example of marked difference in the speed of chromosomal change accumulation in 2 close groups, as in Papionini and Cer-copithecini among Cercopithecidae [Moulin et al., 2008].
Acknowledgements
We thank Michèle Gerbault-Seureau for her excellent techni-cal assistance. This study was partly supported by the Muséum National d’Histoire Naturelle (ATM Biodiversité actuelle et fos-sile).
References
Arslan A: Cytogenetic studies on Spermophilus xanthoprymnus (Rodentia: Sciuridae) in central Anatolia. Folia Zool 54: 278–284 (2005).
Beklemisheva VR, Romanenko SA, Biltueva LS, Trifonov V, Vorobieva NV, et al: Reconstruc-tion of karyotype evolution in core Glires. I. The genome homology revealed by compara-tive chromosome painting. Chromosome Res 19: 549–565 (2011).
Capanna E, Bonomo M, Civitelli MV, Simonetta A: The chromosomes of Royle’s pika, Ochoto-na roylei (Mammalia, lagomorpha). Rend Fis Acc Lincei 2: 59–67 (1991).
Carrasco A, Humphrey D: Chromosomes of the mountain beaver Aplodontia rufa pacifica . Mamm Chr Newsletter 9: 238–239 (1968).
Chang SW, Oshida T, Endo H, Nguyen ST, Dang CN, et al: Ancient hybridization and under-estimated species diversity in Asian striped squirrels (genus Tamiops ): inference from paternal, maternal and biparental markers.J Zool 285: 128–138 (2011).
Chantry-Darmon C, Bertaud M, Urien C, Chadi-Taourit S, Perrocheau M, et al: Expended comparative mapping between man and rab-bit and detection of a new conserved segment between HSA22 and OCU4. Cytogenet Ge-nome Res 111: 134–139 (2005).
Committee for standardized karyotypes of Oryctolagus cuniculus : standard karyotype of the laboratory rabbit. Cytogenet Cell Gen-et 31: 240–248 (1981).
Dobigny G, Ducroz JF, Robinson TJ, Volobouev V: Cytogenetics and cladistics. Syst Biol 53: 470–484 (2004).

Chromosome Evolution in Sciuromorpha and Lagomorpha
Cytogenet Genome Res 2012;137:218–232 231
Douzery EJ, Huchon D: Rabbits, if anything, are likely glires. Mol Phylogenet Evol 33: 922–935 (2004).
Dutrillaux B, Aurias A, Lombard M: Presence of common chromosomes in a rodent ( Eliomys quercinus , garden dormouse) and in Pri-mates [in French]. Ann Génét 22: 21–24 (1979).
Dutrillaux B, Viegas-Pequignot E, Couturier J: Great homology of chromosome banding of the rabbit (Oryctolagus cuniculus) and pri-mates, including man (author’s transl) [in French]. Ann Génét 23: 22–25 (1980).
Dutrillaux B, Couturier J, Sabatier L, Muleris M, Prieur M: Inversions in evolution of man and closely related species. Ann Génét 29: 195–202 (1986).
Ferguson-Smith MA, Trifonov V: Mammalian karyotype evolution. Nature Rev Genet 8: 950–962 (2007).
Filippucci MG, Nevo SS, Capanna E: The chro-mosomes of the Israeli garden dormouse, Eliomys melanurus wagner , 1849 (Rodentia, Gliridae). Boll Zool 55: 31–33 (1988a).
Filippucci MG, Rodino E, Nevo E, Capanna E: Evolutionary genetics and systematics of the garden dormouse, Eliomys wagner 1840.1: al-lozyme diversity and differentiation of chro-mosomal races. Boll Zool 55: 47–54 (1988b).
Graphodatsky AS, Yang F, Dobigny G, Ro-manenko SA, Biltueva LS, et al: Tracking ge-nome organization in rodents by Zoo-FISH. Chromosome Res 16: 261–274 (2008).
Halanych KM, Robinson TJ: Phylogenetic rela-tionship of cottontails ( Sylvilagus , Lagomor-pha): congruence of 12s rDNA and cytoge-netic data. Mol Phylogenet Evol 3: 294–302 (1997).
Halanych KM, Robinson TJ: Multiple substitu-tions affect the phylogenetic utility of cyto-chrome b and 12s rDNA data: examining a rapid radiation in Leporid (Lagomorpha) evolution. J Mol Evol 48: 369–379 (1999).
Herron MD, Castoe TA, Parkinson CL: Sciurid phylogeny and the paraphyly of Holarctic ground squirrels (Spermophilus) . Mol Phylo-genet Evol 31: 1015–1030 (2004).
Hoffmann RS, Smith AT: Order Lagomorpha: Mammal Species of the World, pp 185–193 (The John Hopkins University Press, Balti-more 2005).
Huchon D, Madsen O, Sibbald MJ, Ament K, Stanhope MJ, et al: Rodent phylogeny and a timescale for the evolution of glires: evidence from an extensive taxon sampling using three nuclear genes. Mol Biol Evol 19: 1053–1065 (2002).
Korstanje R, O’Brien PC, Yang F, Rens W, Bosma AA, et al: Complete homology maps of the rabbit (Oryctolagus cuniculus) and human by reciprocal chromosome painting. Cytogenet Cell Genet 86: 317–322 (1999).
Kriegs JO, Zemann A, Churakov G, Matzke A, Ohme M, et al: Retroposon insertions pro-vide insights into deep lagomorph evolution. Mol Biol Evol 27: 2678–2681 (2010).
Li T, O’Brien PC, Biltueva L, Fu B, Wang J, et al: Evolution of genome organization of squir-rels (Sciuridae) revealed by cross-species chromosome painting. Chromosome Res 12: 317–335 (2004).
Li T, Wang J, Su W, Yang F: Karyotypic evolution of the family Sciuridae: inferences from the genome organizations of ground squirrels. Cytogenet Genome Res 112: 270–276 (2006).
Liu J, Li Y, Arnold ML, Wu CH, Wu SF, et al:Reticulate evolution: frequent introgressive hybridization among Chinese hares (genus Lepus ) revealed by analyses of multiple mito-chondrial and nuclear DNA loci. BMC Evol Biol 11: 223–236 (2011).
Luckette WP, Hartenberger JL: Monophyly or polyphyly of the order of Rodentia, possible conflict between morphological and molec-ular interpretations. J Mamm Evol 1: 124–147 (1993).
Matthee CA, van Vuuren BJ, Bell D, Robinson TJ: A molecular supermatrix of the rabbits and hares (Leporidae) allows for the identi-fication of five intercontinental exchanges during the Miocene. Syst Biol 53: 433–447 (2004).
Mercer JM, Roth VL: The effects of Cenozoic global change on squirrel phylogeny. Science 299: 1568–1572 (2003).
Montgelard C, Bentz S, Tirard C, Verneau O, Catzeflis FM: Molecular systematics of Sci-urognathi (Rodentia): the mitochondrial cy-tochrome b and 12S rRNA genes support the Anomaluroidea (Pedetidae and Anomaluri-dae). Mol Phylogenet Evol 22: 220–233 (2002).
Montgelard C, Forty E, Arnal V, Matthee CA: Suprafamilial relationships among Rodentia and the phylogenetic effect of removing fast-evolving nucleotides in mitochondrial, exon and intron fragments. BMC Evol Biol 8: 321 (2008).
Moulin S, Gerbault-Seureau M, Dutrillaux B, Richard FA: Phylogenomics of African gue-nons. Chromosome Res 16: 783–799 (2008).
Morel F, Laudier B, Guérif F, Couet ML, Royère D, et al: Meiotic segregation analysis in sper-matozoa of pericentric inversion carriers us-ing fluorescence in situ hybridization. Hum Reprod 22: 136–141 (2007).
Murphy WJ, Eizirik E, Johnson W, Zkang J, Ry-der O, O’Brien: Molecular phylogenics and the origins of placental mammals. Nature 409: 614–618 (2001a).
Murphy WJ, Stanyon R, O’Brien SJ: Evolution of mammalian genome organization inferred from comparative gene mapping. Genome Biol 2:reviews0005.1–0005.8 (2001b).
Musser GG, Carleton MD: Superfamily Muroi-dea: Mammal Species of the World, pp 894–1531 (The John Hopkins University Press, Baltimore 2005).
Nadler CF, Block MH: The chromosomes of some North American chipmunks (Sciuri-dae) belonging to the genera Tamias and Eu-tamias . Chromosoma 13: 1–15 (1962).
Nadler CF, Sutton DA: Chromosomes of some squirrels (Mammalia-Sciuridae) from the genera Sciurus and Glaucomys . Experientia 23: 249–251 (1967).
Nadler CF, Hoffmann RS: Chromosomes of some Asian and South American squirrels (Rodentia: Sciuridae). Experientia 26: 1383–1386 (1970).
Nadler CF, Hoffmann RS, Hight ME: Chromo-somes of three species of Asian tree squirrels, Callosciurus (Rodentia: Sciuridae). Experi-entia 31: 166–167 (1975).
Niu Y, Wei F, Li M, Liu X, Feng Z: Phylogenyof pikas (Lagomorpha, Ochotona ) inferred from mitochondrial cytochrome b sequenc-es. Folia Zool 53: 141–155 (2004).
Nosil P: Speciation with gene flow could be com-mon. Mol Ecol 17: 2103–2106 (2008).
Oshida T, Su JF, Lin LK: Chromosomal charac-terization of the Formosan striped squirrel Tamiops maritimus formosanus (Mammalia, Rodentia). Caryologia 55: 213–216 (2002).
Petit D, Dutrillaux B: Chromosomal phylogeny of the 7 species of Sciurinae [in French]. Ann Génét 28: 13–18 (1985).
Petit D, Couturier J, Viegas-Péquignot E, Lom-bard M, Dutrillaux B: Great degree of homol-ogy between the ancestral karyotype of squirrels (rodents) and that of primates and carnivores [in French]. Ann Génét 27: 201–212 (1984).
Prasad AB, Allard MW, NISC Comparative Se-quencing Program, Green ED: Confirming the phylogeny of mammals by use of large comparative sequence data sets. Mol Biol Evol 25: 1795–1808 (2008).
Richard F, Messaoudi C, Bonnet-Garnier A, Lombard M, Dutrillaux B: Highly conserved chromosomes in an Asian squirrel ( Menetes berdmorei , Rodentia: Sciuridae) as demon-strated by ZOO-FISH with human probes. Chromosome Res 11: 597–603 (2003a).
Richard F, Lombard M, Dutrillaux B: Recon-struction of the ancestral karyotype of eu-therian mammals. Chromosome Res 11: 605–618 (2003b).
Robinson TJ: Comparative chromosome studies in the family Leporidae (Lagomorpha, Mammalia). Cytogenet Cell Genet 28: 64–70 (1980).
Robinson TJ, Ruiz-Herrera A: Defining the eu-therian ancestral karyotype: a cladistic in-terpretation of chromosome painting and genome sequence assembly data. Chromo-some Res 16: 1133–1141 (2008).
Robinson TJ, Skinner JD: Karyology of the Riv-erin rabbit, Bunolagus monticularis , and its taxonomic implications. J Mamm 64: 678–681 (1983).
Robinson TJ, Elder FF, Lopez-Forment W: Band-ing studies in the Volcano rabbit, Romerola-gus diazi , and Crawshay’s hare, Lepus craw-shayi : evidence of the leporid ancestral karyotype. Can J Genet Cytol 23: 469–474 (1981).

Richard /Dutrillaux
Cytogenet Genome Res 2012;137:218–232232
Robinson TJ, Elder FF, Chapman JA: Evolution of chromosomal variation in cottontails, ge-nus Sylvilagus (Mammalia: Lagomorpha): S. aquaticus, S. f loridanus and S. transitionalis . Cytogenet Cell Genet 35: 216–222 (1983).
Robinson TJ, Elder FF, Chapman JA: Evolution of chromosomal variation in cottontails, ge-nus Sylvilagus (Mammalia: Lagomorpha). II Sylvilagus audubonii , S. idahoensis, S. nut-tallii and S. palustris . Cytogenet Cell Genet 38: 282–289 (1984).
Robinson TJ, Yang F, Harrison WR: Chromo-some painting refines the history of genome evolution in hares and rabbits (order Lago-morpha). Cytogenet Genome Res 96: 223–227 (2002).
Romanenko SA, Lemskaya NA, Beklemisheva VP, Perelman PL, Serdukova NA, Grapho-datsky AS: Comparative cytogenetics of ro-dents. Russ J Genet 46: 1138–1142 (2010).
Romanenko SA, Perelman PL, Trifonov VA, Graphodatsky AS: Chromosomal evolution in Rodentia. Heredity 108: 4–16 (2012).
Ruedas LA, Dowler RC, Aita E: Chromosomal variation in the New England cottontail, Syl-vilagus transitionalis . J Mamm 70: 860–864 (1989).
Ruiz-Herrera A, Farré M, Robinson TJ: Molecu-lar cytogenetic and genomic insights into chromosomal evolution. Heredity 108: 28–36 (2012).
Sannier J, Gerbault-Seureau M, Dutrillaux B, Richard FA: Conserved although very differ-ent karyotypes in Gliridae and Sciuridae and their contribution to chromosomal signa-tures in glires. Cytogenet Genome Res 134: 51–63 (2011).
Stanyon R, Yang F, Cavagna P, O’Brien PC, Bag-ga M, et al: Reciprocal chromosome painting shows that genomic rearrangement between rat and mouse proceeds ten times faster than between humans and cats. Cytogenet Ge-nome Res 84: 150–155 (1999).
Stanyon R, Stone G, Garcia M, Froenicke L: Re-ciprocal chromosome painting shows that squirrels, unlike murid rodents, have a high-ly conserved genome organization. Genom-ics 82: 245–249 (2003).
Steppan SJ, Storz BL, Hoffman RS: Nuclear DNA phylogeny of the squirrels (Mammalia: Ro-dentia) and the evolution of arboreality from c-myc and RAG1. Mol Phylogenet Evol 30: 703–719 (2004).
Stock AD: Chromosome banding pattern rela-tionships of hares, rabbits, and pikas (order Lagomorpha). A phyletic interpretation. Cy-togenet Cell Genet 17: 78–88 (1976).
Vorontsov NN, Ivanitskaya EY: Comparative karyology of north Palearctic pikas ( Ochoto-na , Ochotonidae, Lagomorpha). Caryologi-ca 26: 213–223 (1973).
Ye J, Nie W, Wang J, Su W, Jing M, et al: Genome-wide comparative chromosome map be-tween human and the Forrest’s pika (Ocho-tona forresti) established by cross-species chromosome painting: further support for the Glires hypothesis. Cytogenet Genome Res 132: 41–46 (2011).
Yu N, Zheng C, Zhang YP, Li WH: Molecular systematics of pikas (genus Ochotona ) in-ferred from mitochondrial DNA sequences. Mol Phylogenet Evol 16: 85–95 (2000).
Zima J, Macholan M, Filippucci MG: Chromo-somal variation and systematics of myoxids. Hystrix 6: 63–76 (1995).
Recommended

















